
Mechanosensitive Pannexin 1 Activity Is Modulated by Stomatin in Human Red Blood Cells

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Deformability Properties over an Osmotic Gradient Measured in RBCs from Controls and OHSt Patients
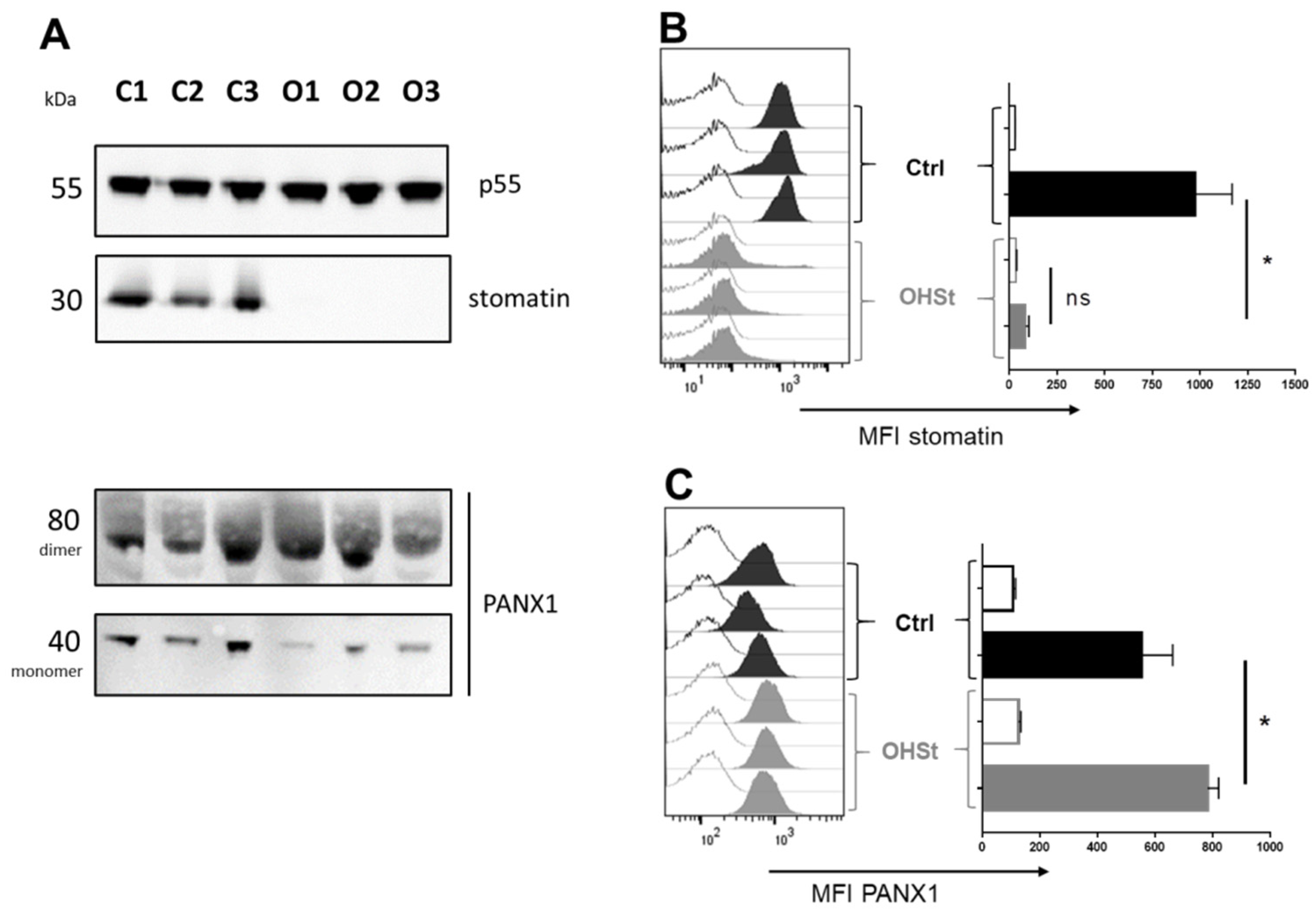
2.2. Stomatin and PANX1 Expression at the RBC Membrane of Controls and OHSt Patients
2.3. Proximity between Stomatin and PANX1 in Control RBCs
2.4. ATP Release from RBCs
2.5. 5(6)-Carboxyfluorescein (CF) Uptake after High K+ Stimulation of Control and OHSt RBC Membranes
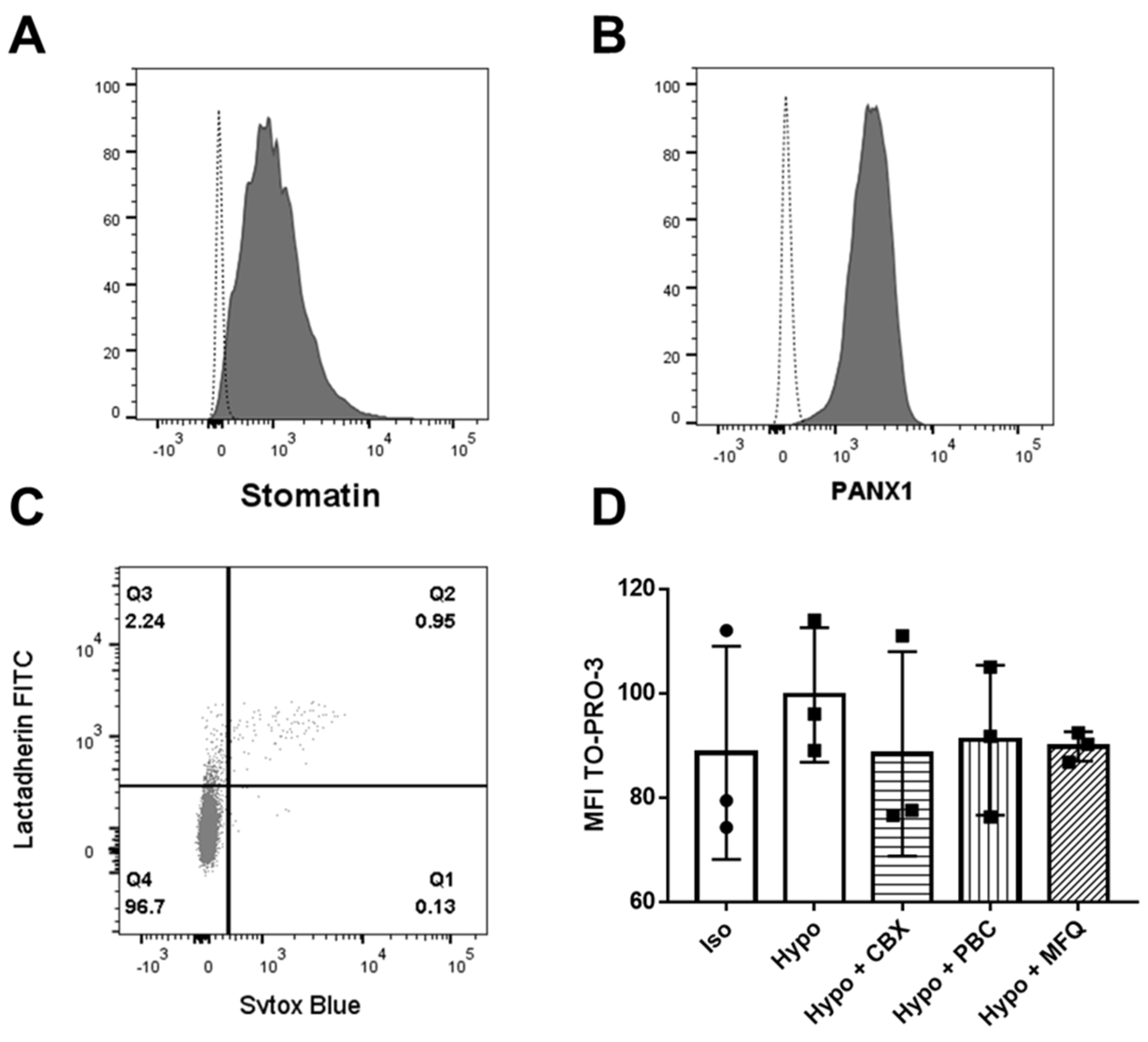
2.6. Endogenous Expression of Stomatin and PANX1 in K562 Cells and PANX1-Dependent TO-PRO-3 Uptake
2.7. Generation of Stomatin CRISPR/Cas9 Knockout K562 Cell Clones
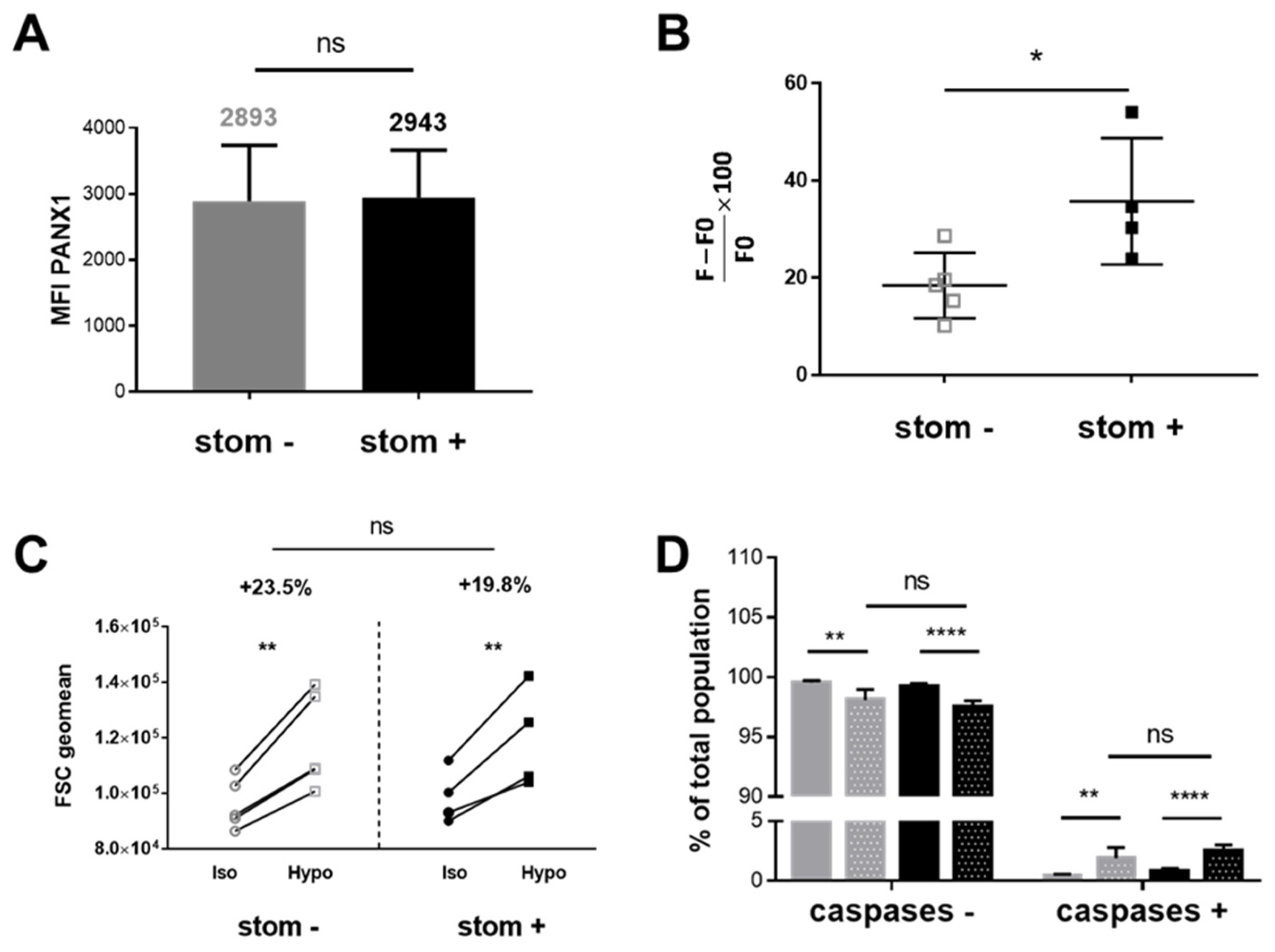
2.8. PANX1 Expression and Caspase-Independent TO-PRO-3 Uptake in stom+ and stom− K562 Cell Clones
3. Discussion
4. Methods and Materials
4.1. Control and Patient RBCs
4.2. RBC Deformability Index and Elongation Index Measurements by Ektacytometry
4.3. Cell Culture and Transfection
4.4. Stomatin and PANX1 Membrane Expression Analysis by Flow Cytometry
4.5. Stomatin and PANX1 Membrane Expression Analysis by Western Blot
4.6. Proximity Ligation Assay
4.7. Dye Uptakes
4.8. ATP Release and Hemolysis
4.9. PS Exposure and Caspase Activity on RBCs
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penuela, S.; Gehi, R.; Laird, D.W. The biochemistry and function of pannexin channels. Biochim. Biophys. Acta 2013, 1828, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Sosinsky, G.E.; Boassa, D.; Dermietzel, R.; Duffy, H.S.; Laird, D.W.; MacVicar, B.; Naus, C.C.; Penuela, S.; Scemes, E.; Spray, D.C.; et al. Pannexin channels are not gap junction hemichannels. Channels 2011, 5, 193–197. [Google Scholar] [CrossRef]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.H.; Jin, X.; Medina, C.B.; Leonhardt, S.A.; Kiessling, V.; Bennett, B.C.; Shu, S.; Tamm, L.K.; Yeager, M.; Ravichandran, K.S.; et al. A quantized mechanism for activation of pannexin channels. Nat. Commun. 2017, 8, 14324. [Google Scholar] [CrossRef]
- Sandilos, J.K.; Chiu, Y.H.; Chekeni, F.B.; Armstrong, A.J.; Walk, S.F.; Ravichandran, K.S.; Bayliss, D.A. Pannexin 1, an ATP release channel, is activated by caspase cleavage of its pore-associated C-terminal autoinhibitory region. J. Biol. Chem. 2012, 287, 11303–11311. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; He, Z.; Maksaev, G.; Bitter, R.M.; Rau, M.; Fitzpatrick, J.A.J.; Yuan, P. Cryo-EM structures of the ATP release channel pannexin 1. Nat. Struct. Mol. Biol. 2020, 27, 373–381. [Google Scholar] [CrossRef]
- Jin, Q.; Zhang, B.; Zheng, X.; Li, N.; Xu, L.; Xie, Y.; Song, F.; Bhat, E.A.; Chen, Y.; Gao, N.; et al. Cryo-EM structures of human pannexin 1 channel. Cell Res. 2020, 30, 449–451. [Google Scholar] [CrossRef]
- Michalski, K.; Syrjanen, J.L.; Henze, E.; Kumpf, J.; Furukawa, H.; Kawate, T. The Cryo-EM structure of pannexin 1 reveals unique motifs for ion selection and inhibition. eLife 2020, 9, e54670. [Google Scholar] [CrossRef] [Green Version]
- Mou, L.; Ke, M.; Song, M.; Shan, Y.; Xiao, Q.; Liu, Q.; Li, J.; Sun, K.; Pu, L.; Guo, L.; et al. Structural basis for gating mechanism of Pannexin 1 channel. Cell Res. 2020, 30, 452–454. [Google Scholar] [CrossRef] [Green Version]
- Qu, R.; Dong, L.; Zhang, J.; Yu, X.; Wang, L.; Zhu, S. Cryo-EM structure of human heptameric Pannexin 1 channel. Cell Res. 2020, 30, 446–448. [Google Scholar] [CrossRef]
- Ruan, Z.; Orozco, I.J.; Du, J.; Lu, W. Structures of human pannexin 1 reveal ion pathways and mechanism of gating. Nature 2020, 584, 646–651. [Google Scholar] [CrossRef]
- Mim, C.; Perkins, G.; Dahl, G. Structure versus function: Are new conformations of pannexin 1 yet to be resolved? J. Gen. Physiol. 2021, 153, e202012754. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Schappe, M.S.; Desai, B.N.; Bayliss, D.A. Revisiting multimodal activation and channel properties of Pannexin 1. J. Gen. Physiol. 2018, 150, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Kuzuya, M.; Hirano, H.; Hayashida, K.; Watanabe, M.; Kobayashi, K.; Terada, T.; Mahmood, M.I.; Tama, F.; Tani, K.; Fujiyoshi, Y.; et al. Structures of human pannexin-1 in nanodiscs reveal gating mediated by dynamic movement of the N terminus and phospholipids. Sci. Signal. 2022, 15, eabg6941. [Google Scholar] [CrossRef]
- Bao, L.; Locovei, S.; Dahl, G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Leal Denis, M.F.; Lefevre, S.D.; Alvarez, C.L.; Lauri, N.; Enrique, N.; Rinaldi, D.E.; Gonzalez-Lebrero, R.; Vecchio, L.E.; Espelt, M.V.; Stringa, P.; et al. Regulation of extracellular ATP of human erythrocytes treated with alpha-hemolysin. Effects of cell volume, morphology, rheology and hemolysis. Biochim. Biophys Acta Mol. Cell Res. 2019, 1866, 896–915. [Google Scholar] [CrossRef]
- Locovei, S.; Wang, J.; Dahl, G. Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett. 2006, 580, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, C.L.; Chene, A.; Semblat, J.P.; Gamain, B.; Lapoumeroulie, C.; Fader, C.M.; Hattab, C.; Sevigny, J.; Denis, M.F.L.; Lauri, N.; et al. Homeostasis of extracellular ATP in uninfected RBCs from a Plasmodium falciparum culture and derived microparticles. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183980. [Google Scholar] [CrossRef]
- Keller, A.S.; Diederich, L.; Panknin, C.; DeLalio, L.J.; Drake, J.C.; Sherman, R.; Jackson, E.K.; Yan, Z.; Kelm, M.; Cortese-Krott, M.M.; et al. Possible roles for ATP release from RBCs exclude the cAMP-mediated Panx1 pathway. Am. J. Physiol. Cell Physiol. 2017, 313, C593–C603. [Google Scholar] [CrossRef] [Green Version]
- Locovei, S.; Bao, L.; Dahl, G. Pannexin 1 in erythrocytes: Function without a gap. Proc. Natl. Acad. Sci. USA 2006, 103, 7655–7659. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Wang, J.; Spray, D.C.; Scemes, E.; Dahl, G. Two non-vesicular ATP release pathways in the mouse erythrocyte membrane. FEBS Lett. 2011, 585, 3430–3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, M.; Adderley, S.P.; Bowles, E.A.; Egan, T.M.; Stephenson, A.H.; Ellsworth, M.L.; Sprague, R.S. Pannexin 1 is the conduit for low oxygen tension-induced ATP release from human erythrocytes. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1146–H1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal Denis, M.F.; Incicco, J.J.; Espelt, M.V.; Verstraeten, S.V.; Pignataro, O.P.; Lazarowski, E.R.; Schwarzbaum, P.J. Kinetics of extracellular ATP in mastoparan 7-activated human erythrocytes. Biochim. Biophys. Acta 2013, 1830, 4692–4707. [Google Scholar] [CrossRef] [Green Version]
- Montalbetti, N.; Leal Denis, M.F.; Pignataro, O.P.; Kobatake, E.; Lazarowski, E.R.; Schwarzbaum, P.J. Homeostasis of extracellular ATP in human erythrocytes. J. Biol. Chem. 2011, 286, 38397–38407. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Dahl, G. A permeant regulating its permeation pore: Inhibition of pannexin 1 channels by ATP. Am. J. Physiol. Cell Physiol. 2009, 296, C250–C255. [Google Scholar] [CrossRef]
- Kirby, B.S.; Sparks, M.A.; Lazarowski, E.R.; Lopez Domowicz, D.A.; Zhu, H.; McMahon, T.J. Pannexin 1 channels control the hemodynamic response to hypoxia by regulating O2-sensitive extracellular ATP in blood. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1055–H1065. [Google Scholar] [CrossRef]
- Zhan, H.; Moore, C.S.; Chen, B.; Zhou, X.; Ma, X.M.; Ijichi, K.; Bennett, M.V.; Li, X.J.; Crocker, S.J.; Wang, Z.W. Stomatin inhibits pannexin-1-mediated whole-cell currents by interacting with its carboxyl terminal. PLoS ONE 2012, 7, e39489. [Google Scholar] [CrossRef] [Green Version]
- Brand, J.; Smith, E.S.; Schwefel, D.; Lapatsina, L.; Poole, K.; Omerbasic, D.; Kozlenkov, A.; Behlke, J.; Lewin, G.R.; Daumke, O. A stomatin dimer modulates the activity of acid-sensing ion channels. EMBO J. 2012, 31, 3635–3646. [Google Scholar] [CrossRef] [Green Version]
- Langhorst, M.F.; Reuter, A.; Stuermer, C.A. Scaffolding microdomains and beyond: The function of reggie/flotillin proteins. Cell. Mol. Life Sci.CMLS 2005, 62, 2228–2240. [Google Scholar] [CrossRef] [Green Version]
- Tavernarakis, N.; Driscoll, M.; Kyrpides, N.C. The SPFH domain: Implicated in regulating targeted protein turnover in stomatins and other membrane-associated proteins. Trends Biochem. Sci. 1999, 24, 425–427. [Google Scholar] [CrossRef]
- Lapatsina, L.; Brand, J.; Poole, K.; Daumke, O.; Lewin, G.R. Stomatin-domain proteins. Eur. J. Cell Biol. 2012, 91, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Klipp, R.C.; Cullinan, M.M.; Bankston, J.R. Insights into the molecular mechanisms underlying the inhibition of acid-sensing ion channel 3 gating by stomatin. J. Gen. Physiol. 2020, 152, e201912471. [Google Scholar] [CrossRef] [PubMed]
- Appelman, M.D.; Robin, M.J.D.; Vogels, E.W.M.; Wolzak, C.; Vos, W.G.; Vos, H.R.; Van Es, R.M.; Burgering, B.M.T.; Van de Graaf, S.F.J. The Lipid Raft Component Stomatin Interacts with the Na+ Taurocholate Cotransporting Polypeptide (NTCP) and Modulates Bile Salt Uptake. Cells 2020, 9, 986. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.K.; Turner, E.J.; Parkin, E.T.; Garner, A.E.; Harrison, P.J.; Crawford, M.; Stewart, G.W.; Hooper, N.M. Membrane raft actin deficiency and altered Ca2+-induced vesiculation in stomatin-deficient overhydrated hereditary stomatocytosis. Biochim. Biophys. Acta 2008, 1778, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Hiebl-Dirschmied, C.M.; Adolf, G.R.; Prohaska, R. Isolation and partial characterization of the human erythrocyte band 7 integral membrane protein. Biochim. Biophys. Acta 1991, 1065, 195–202. [Google Scholar] [CrossRef]
- Hiebl-Dirschmied, C.M.; Entler, B.; Glotzmann, C.; Maurer-Fogy, I.; Stratowa, C.; Prohaska, R. Cloning and nucleotide sequence of cDNA encoding human erythrocyte band 7 integral membrane protein. Biochim. Biophys. Acta 1991, 1090, 123–124. [Google Scholar] [CrossRef]
- Stewart, G.W.; Hepworth-Jones, B.E.; Keen, J.N.; Dash, B.C.; Argent, A.C.; Casimir, C.M. Isolation of cDNA coding for an ubiquitous membrane protein deficient in high Na+, low K+ stomatocytic erythrocytes. Blood 1992, 79, 1593–1601. [Google Scholar] [CrossRef] [Green Version]
- Rungaldier, S.; Oberwagner, W.; Salzer, U.; Csaszar, E.; Prohaska, R. Stomatin interacts with GLUT1/SLC2A1, band 3/SLC4A1, and aquaporin-1 in human erythrocyte membrane domains. Biochim. Biophys. Acta 2013, 1828, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Genetet, S.; Desrames, A.; Chouali, Y.; Ripoche, P.; Lopez, C.; Mouro-Chanteloup, I. Stomatin modulates the activity of the Anion Exchanger 1 (AE1, SLC4A1). Sci. Rep. 2017, 7, 46170. [Google Scholar] [CrossRef] [Green Version]
- Montel-Hagen, A.; Kinet, S.; Manel, N.; Mongellaz, C.; Prohaska, R.; Battini, J.L.; Delaunay, J.; Sitbon, M.; Taylor, N. Erythrocyte Glut1 triggers dehydroascorbic acid uptake in mammals unable to synthesize vitamin C. Cell 2008, 132, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Bruce, L.J. Hereditary stomatocytosis and cation-leaky red cells—Recent developments. Blood Cells Mol. Dis. 2009, 42, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Caulier, A.; Rapetti-Mauss, R.; Guizouarn, H.; Picard, V.; Garcon, L.; Badens, C. Primary red cell hydration disorders: Pathogenesis and diagnosis. Int. J. Lab. Hematol. 2018, 40 (Suppl. S1), 68–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, P.G. Disorders of erythrocyte hydration. Blood 2017, 130, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Roussel, C.; Dussiot, M.; Ndour, P.A.; Hermine, O.; Colin, Y.; Gray, A.; Landrigan, M.; Le Van Kim, C.; Buffet, P.A.; et al. Metabolic rejuvenation upgrades circulatory functions of red blood cells stored under blood bank conditions. Transfusion 2021, 61, 903–918. [Google Scholar] [CrossRef]
- Gautier, E.F.; Ducamp, S.; Leduc, M.; Salnot, V.; Guillonneau, F.; Dussiot, M.; Hale, J.; Giarratana, M.C.; Raimbault, A.; Douay, L.; et al. Comprehensive Proteomic Analysis of Human Erythropoiesis. Cell Rep. 2016, 16, 1470–1484. [Google Scholar] [CrossRef] [Green Version]
- Boassa, D.; Ambrosi, C.; Qiu, F.; Dahl, G.; Gaietta, G.; Sosinsky, G. Pannexin1 channels contain a glycosylation site that targets the hexamer to the plasma membrane. J. Biol. Chem. 2007, 282, 31733–31743. [Google Scholar] [CrossRef] [Green Version]
- Salzer, U.; Prohaska, R. Stomatin, flotillin-1, and flotillin-2 are major integral proteins of erythrocyte lipid rafts. Blood 2001, 97, 1141–1143. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G. The Pannexin1 membrane channel: Distinct conformations and functions. FEBS Lett. 2018, 592, 3201–3209. [Google Scholar] [CrossRef] [Green Version]
- Marginedas-Freixa, I.; Alvarez, C.L.; Moras, M.; Leal Denis, M.F.; Hattab, C.; Halle, F.; Bihel, F.; Mouro-Chanteloup, I.; Lefevre, S.D.; Le Van Kim, C.; et al. Human erythrocytes release ATP by a novel pathway involving VDAC oligomerization independent of pannexin-1. Sci. Rep. 2018, 8, 11384. [Google Scholar] [CrossRef] [Green Version]
- Eber, S.W.; Lande, W.M.; Iarocci, T.A.; Mentzer, W.C.; Hohn, P.; Wiley, J.S.; Schroter, W. Hereditary stomatocytosis: Consistent association with an integral membrane protein deficiency. Br. J. Haematol. 1989, 72, 452–455. [Google Scholar] [CrossRef]
- Lande, W.M.; Thiemann, P.V.; Mentzer, W.C., Jr. Missing band 7 membrane protein in two patients with high Na, low K erythrocytes. J. Clin. Investig. 1982, 70, 1273–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarkowsky, H.S.; Oski, F.A.; Sha’afi, R.; Shohet, S.B.; Nathan, D.G. Congenital hemolytic anemia with high sodium, low potassium red cells. I. Studies of membrane permeability. N. Engl. J. Med. 1968, 278, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Fricke, B.; Argent, A.C.; Chetty, M.C.; Pizzey, A.R.; Turner, E.J.; Ho, M.M.; Iolascon, A.; von During, M.; Stewart, G.W. The “stomatin” gene and protein in overhydrated hereditary stomatocytosis. Blood 2003, 102, 2268–2277. [Google Scholar] [CrossRef] [PubMed]
- Llaudet-Planas, E.; Vives-Corrons, J.L.; Rizzuto, V.; Gomez-Ramirez, P.; Sevilla Navarro, J.; Coll Sibina, M.T.; Garcia-Bernal, M.; Ruiz Llobet, A.; Badell, I.; Velasco-Puyo, P.; et al. Osmotic gradient ektacytometry: A valuable screening test for hereditary spherocytosis and other red blood cell membrane disorders. Int. J. Lab. Hematol. 2018, 40, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.L.; Thompson, R.J. Intrapore lipids hydrophobically gate pannexin-1 channels. Sci. Signal. 2022, 15, eabn2081. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, R.; Spray, D.C.; Scemes, E. Mefloquine blockade of Pannexin1 currents: Resolution of a conflict. Cell Commun. Adhes. 2009, 16, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navis, K.E.; Fan, C.Y.; Trang, T.; Thompson, R.J.; Derksen, D.J. Pannexin 1 Channels as a Therapeutic Target: Structure, Inhibition, and Outlook. ACS Chem. Neurosci. 2020, 11, 2163–2172. [Google Scholar] [CrossRef]
- Jiang, L.; Tixeira, R.; Caruso, S.; Atkin-Smith, G.K.; Baxter, A.A.; Paone, S.; Hulett, M.D.; Poon, I.K. Monitoring the progression of cell death and the disassembly of dying cells by flow cytometry. Nat. Protoc. 2016, 11, 655–663. [Google Scholar] [CrossRef]
- Desrames, A.; Genetet, S.; Delcourt, M.P.; Goossens, D.; Mouro-Chanteloup, I. Detergent-free isolation of native red blood cell membrane complexes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183126. [Google Scholar] [CrossRef]
- Clark, M.R.; Mohandas, N.; Shohet, S.B. Osmotic gradient ektacytometry: Comprehensive characterization of red cell volume and surface maintenance. Blood 1983, 61, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Mohandas, N.; Clark, M.R.; Health, B.P.; Rossi, M.; Wolfe, L.C.; Lux, S.E.; Shohet, S.B. A technique to detect reduced mechanical stability of red cell membranes: Relevance to elliptocytic disorders. Blood 1982, 59, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, A.; Tudela, J.; Varon, R.; Garcia-Canovas, F. Determination of hemoglobin through its peroxidase activity on chlorpromazine. J. Biochem. Biophys. Methods 1991, 23, 45–52. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rougé, S.; Genetet, S.; Leal Denis, M.F.; Dussiot, M.; Schwarzbaum, P.J.; Ostuni, M.A.; Mouro-Chanteloup, I. Mechanosensitive Pannexin 1 Activity Is Modulated by Stomatin in Human Red Blood Cells. Int. J. Mol. Sci. 2022, 23, 9401. https://doi.org/10.3390/ijms23169401
Rougé S, Genetet S, Leal Denis MF, Dussiot M, Schwarzbaum PJ, Ostuni MA, Mouro-Chanteloup I. Mechanosensitive Pannexin 1 Activity Is Modulated by Stomatin in Human Red Blood Cells. International Journal of Molecular Sciences. 2022; 23(16):9401. https://doi.org/10.3390/ijms23169401
Chicago/Turabian StyleRougé, Sarah, Sandrine Genetet, Maria Florencia Leal Denis, Michael Dussiot, Pablo Julio Schwarzbaum, Mariano Anibal Ostuni, and Isabelle Mouro-Chanteloup. 2022. "Mechanosensitive Pannexin 1 Activity Is Modulated by Stomatin in Human Red Blood Cells" International Journal of Molecular Sciences 23, no. 16: 9401. https://doi.org/10.3390/ijms23169401