
Paradoxical Anxiety Level Reduction in Animal Chronic Stress: A Unique Role of Hippocampus Neurobiology

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. PS-Separated Sprague–Dawley Rats into Two Behavioral Phenotypes
2.2. Predator Scent Stress Reduced the Plasma Corticosterone Levels in Low-Anxiety Rats
2.3. Predator Scent Stress Increased the Blood Flow in Carotid Arteries in Low-Anxiety Rats
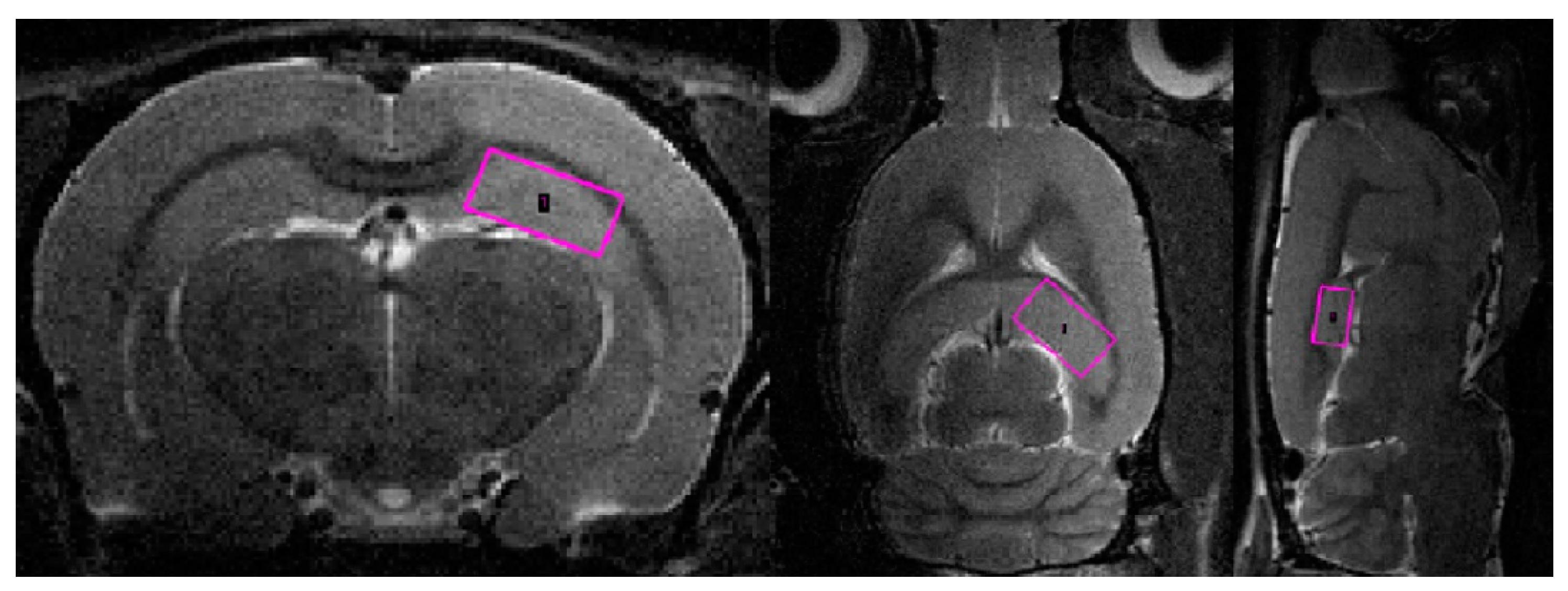
2.4. Predator Scent Stress Reduced Apparent Diffusion Coefficient Values in the Hippocampus of Low-Anxiety Rats
2.5. Predator Scent Stress Altered 1H MR Spectra in the Hippocampus in High- and Low-Anxiety Rats
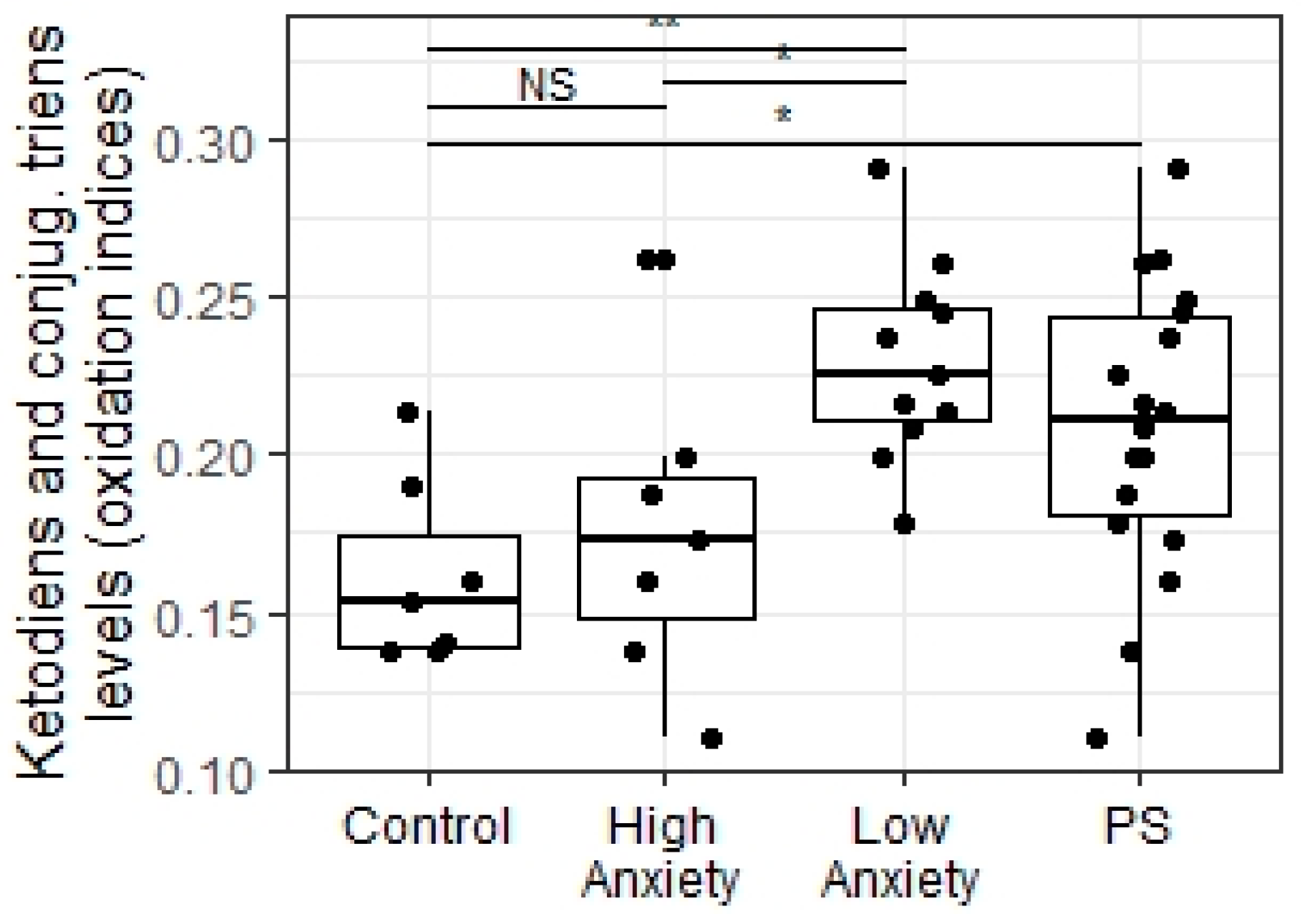
2.6. Predator Scent Stress Increased BDNF Gene Expression in Low-Anxiety Rats
3. Discussion
4. Methods
4.1. Experimental Procedure
- Days 1–10: PSS;
- Days 11–22: rest;
- Day 23: elevated plus maze test;
- Day 24 blood flow rate in carotid artery measurement by MRA;
- Day 25 hippocampal apparent diffusion coefficient measurement by MRI;
- Day 27: hippocampal metabolite measurement by MRS;
- Day 28: euthanasia, harvesting of blood and organs.
4.2. Behavioral Activity
4.3. Magnetic Resonance Imaging
4.4. Magnetic Resonance Angiography
4.5. Diffusion-Weighted Magnetic Resonance Imaging
4.6. Magnetic Resonance Spectroscopy (MRS)
4.7. Hormonal Measurements
4.8. Evaluation of Oxidative Stress
4.9. BDNF Gene Expression in the Hippocampus
4.10. Data Analyses
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saeedi, M.; Rashidy-Pour, A. Association between chronic stress and Alzheimer’s disease: Therapeutic effects of Saffron. Biomed. Pharmacother. 2020, 133, 110995. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; Juster, R.-P.; Raymond, C.; Marin, M.-F. The effects of chronic stress on the human brain: From neurotoxicity, to vulnerability, to opportunity. Front. Neuroendocrinol. 2018, 49, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.D.; Kogan, J.F.; Marrocco, J.; McEwen, B.S. Genomic and epigenomic mechanisms of glucocorticoids in the brain. Nat. Rev. Endocrinol. 2017, 13, 661–673. [Google Scholar] [CrossRef]
- Conrad, C.D.; Ortiz, J.B.; Judd, J.M. Chronic stress and hippocampal dendritic complexity: Methodological and functional considerations. Physiol. Behav. 2017, 178, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Oomen, C.A.; Naninck, E.F.; Fitzsimons, C.P.; Van Dam, A.-M.; Czeh, B.; Korosi, A. Regulation of Adult Neurogenesis and Plasticity by (Early) Stress, Glucocorticoids, and Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021303. [Google Scholar] [CrossRef] [PubMed]
- Laham, B.J.; Gould, E. How Stress Influences the Dynamic Plasticity of the Brain’s Extracellular Matrix. Front. Cell. Neurosci. 2022, 15, 814287. [Google Scholar] [CrossRef] [PubMed]
- Surget, A.; Belzung, C. Adult hippocampal neurogenesis shapes adaptation and improves stress response: A mechanistic and integrative perspective. Mol. Psychiatry 2021, 27, 403–421. [Google Scholar] [CrossRef]
- Vlachos, I.I.; Papageorgiou, C.; Margariti, M. Neurobiological Trajectories Involving Social Isolation in PTSD: A Systematic Review. Brain Sci. 2020, 10, 173. [Google Scholar] [CrossRef] [PubMed]
- Cabib, S.; Campus, P.; Conversi, D.; Orsini, C.; Puglisi-Allegra, S. Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress. Brain Sci. 2020, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Montes-Rodriguez, C.J.; Urteaga-Urias, E. Plasticidad sinaptica como sustrato de la resiliencia Synaptic plasticity as a substrate of resilience. Rev. Neurol. 2018, 67, 453–460. [Google Scholar]
- McEwen, B.S.; Eiland, L.; Hunter, R.G.; Miller, M.M. Stress and anxiety: Structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology 2012, 62, 3–12. [Google Scholar] [CrossRef]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, M.; Navidhamidi, M.; Rezaei, F.; Azizikia, A.; Mehranfard, N. Anxiety and hippocampal neuronal activity: Relationship and potential mechanisms. Cogn. Affect. Behav. Neurosci. 2021, 22, 431–449. [Google Scholar] [CrossRef]
- Tomar, A.; McHugh, T.J. The impact of stress on the hippocampal spatial code. Trends Neurosci. 2021, 45, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Shevelev, O.B.; Tseilikman, V.E.; Khotskin, N.V.; Khotskina, A.; Kontsevaya, G.V.; Lapshin, M.S.; Moshkin, M.P.; Komelkova, M.; Feklicheva, I.V.; Tseilikman, O.B.; et al. Erratum to: Anxiety and neurometabolite levels in the hippocampus and amygdala after prolonged exposure to predator-scent stress. Vavilov J. Genet. Breed. 2020, 24, 108. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, E.; Chrousos, G.; Perry, S.W.; Wong, M.-L.; Licinio, J.; Bornstein, S.R.; Tseilikman, O.; Komelkova, M.; Lapshin, M.S.; Vasilyeva, M.; et al. Offensive Behavior, Striatal Glutamate Metabolites, and Limbic-Hypothalamic-Pituitary-Adrenal Responses to Stress in Chronic Anxiety. Int. J. Mol. Sci. 2020, 21, 7440. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Stress-induced remodeling of hippocampal CA3 pyramidal neurons. Brain Res. 2016, 1645, 50–54. [Google Scholar] [CrossRef]
- Nasca, C.; Zelli, D.; Bigio, B.; Piccinin, S.; Scaccianoce, S.; Nisticò, R.; McEwen, B.S. Stress dynamically regulates behavior and glutamatergic gene expression in hippocampus by opening a window of epigenetic plasticity. Proc. Natl. Acad. Sci. USA 2015, 112, 14960–14965. [Google Scholar] [CrossRef]
- Podgorny, O.V.; Gulyaeva, N.V. Glucocorticoid-mediated mechanisms of hippocampal damage: Contribution of subgranular neurogenesis. J. Neurochem. 2020, 157, 370–392. [Google Scholar] [CrossRef]
- Brocca, M.E.; Pietranera, L.; de Kloet, E.R.; De Nicola, A.F. Mineralocorticoid Receptors, Neuroinflammation and Hypertensive Encephalopathy. Cell. Mol. Neurobiol. 2018, 39, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Uchoa, E.T.; Aguilera, G.; Herman, J.P.; Fiedler, J.L.; Deak, T.; de Sousa, M.B.C. Novel aspects of glucocorticoid actions. J. Neuroendocr. 2014, 26, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Chaouloff, F.; Groc, L. Temporal modulation of hippocampal excitatory transmission by corticosteroids and stress. Front. Neuroendocr. 2011, 32, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Rogalska, J. Mineralocorticoid and glucocorticoid receptors in hippocampus: Their impact on neurons survival and behavioral impairment after neonatal brain injury. Vitam. Horm. 2010, 82, 391–419. [Google Scholar] [CrossRef]
- Joëls, M. Functional actions of corticosteroids in the hippocampus. Eur. J. Pharmacol. 2008, 583, 312–321. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Karst, H.; Joëls, M. Corticosteroid hormones in the central stress response: Quick-and-slow. Front. Neuroendocr. 2008, 29, 268–272. [Google Scholar] [CrossRef]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid Generates ROS to Induce Oxidative Injury in the Hippocampus, Leading to Impairment of Cognitive Function of Rats. J. Clin. Biochem. Nutr. 2010, 47, 224–232. [Google Scholar] [CrossRef]
- You, J.-M.; Yun, S.-J.; Nam, K.N.; Kang, C.; Won, R.; Lee, E.H. Mechanism of glucocorticoid-induced oxidative stress in rat hippocampal slice cultures. Can. J. Physiol. Pharmacol. 2009, 87, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Lope-Piedrafita, S. Diffusion Tensor Imaging (DTI). Methods Mol. Biol. 2018, 1718, 103–116. [Google Scholar] [CrossRef]
- Scotton, E.; Colombo, R.; Reis, J.C.; Possebon, G.M.; Hizo, G.H.; Valiati, F.E.; Géa, L.P.; Bristot, G.; Salvador, M.; Silva, T.M.; et al. BDNF prevents central oxidative damage in a chronic unpredictable mild stress model: The possible role of PRDX-1 in anhedonic behavior. Behav. Brain Res. 2019, 378, 112245. [Google Scholar] [CrossRef] [PubMed]
- Merz, E.C.; Monk, C.; Bansal, R.; Sawardekar, S.; Lee, S.; Feng, T.; Spann, M.; Foss, S.; McDonough, L.; Werner, E.; et al. Neonatal brain metabolite concentrations: Associations with age, sex, and developmental outcomes. PLoS ONE 2020, 15, e0243255. [Google Scholar] [CrossRef]
- Yamazaki, K.; Yoshimura, A.; Miyahara, S.; Sugi, S.; Itono, M.; Kondo, M.; Tsuji, N.; Shimizu, M.; Fukushima, R.; Kishimoto, M. Evaluation of cerebral blood flow in the hippocampus, thalamus, and basal ganglia and the volume of the hippocampus in dogs before and during treatment with prednisolone. Am. J. Veter. Res. 2021, 82, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.A.; Cada, A.M. Spatial learning/memory and social and nonsocial behaviors in the spontaneously hypertensive, Wistar-Kyoto and Sprague-Dawley rat strains. Pharmacol. Biochem. Behav. 2004, 77, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Tseilikman, V.; Komelkova, M.; Lapshin, M.; Alliluev, A.; Tseilikman, O.; Karpenko, M.; Pestereva, N.; Manukhina, E.; Downey, H.F.; Kondashevskaya, M.; et al. High and low anxiety phenotypes in a rat model of complex post-traumatic stress disorder are associated with different alterations in regional brain monoamine neurotransmission. Psychoneuroendocrinology 2020, 117, 104691. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, E.; Perry, S.W.; Licinio, J.; Wong, M.-L.; Dremencov, E.; Zavjalov, E.L.; Shevelev, O.B.; Khotskin, N.V.; Koncevaya, G.V.; Khotshkina, A.S.; et al. From Allostatic Load to Allostatic State-An Endogenous Sympathetic Strategy to Deal with Chronic Anxiety and Stress? Front. Behav. Neurosci. 2019, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Komelkova, M.; Manukhina, E.; Downey, H.F.; Sarapultsev, A.; Cherkasova, O.; Kotomtsev, V.; Platkovskiy, P.; Fedorov, S.; Sarapultsev, P.; Tseilikman, O.; et al. Hexobarbital Sleep Test for Predicting the Susceptibility or Resistance to Experimental Posttraumatic Stress Disorder. Int. J. Mol. Sci. 2020, 21, 5900. [Google Scholar] [CrossRef] [PubMed]
- Del Moro, L.; Rota, E.; Pirovano, E.; Rainero, I. Migraine, Brain Glucose Metabolism and the “Neuroenergetic” Hypothesis: A Scoping Review. J. Pain 2022, 23, 1294–1317. [Google Scholar] [CrossRef]
- Nofal, A.M.; Sayyed, T.M.; Mahmoud, H.S.; Atia, T.Y. Effects of dexamethasone on foetal Doppler flow velocimetry. J. Obstet. Gynaecol. 2020, 41, 390–394. [Google Scholar] [CrossRef]
- Ben Alaya, I.; Limam, H.; Kraiem, T. Applications of artificial intelligence for DWI and PWI data processing in acute ischemic stroke: Current practices and future directions. Clin. Imaging 2021, 81, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, N. Diffusion weighted imaging in acute ischemic stroke: A review of its interpretation pitfalls and advanced diffusion imaging application. J. Neurol. Sci. 2021, 425, 117435. [Google Scholar] [CrossRef]
- Birken, D.L.; Oldendorf, W.H. N-acetyl-L-aspartic acid: A literature review of a compound prominent in 1H-NMR spectroscopic studies of brain. Neurosci. Biobehav. Rev. 1989, 13, 23–31. [Google Scholar] [CrossRef]
- Neylan, T.C.; Schuff, N.; Lenoci, M.; Yehuda, R.; Weiner, M.W.; Marmar, C.R. Cortisol levels are positively correlated with hippocampal N-acetylaspartate. Biol. Psychiatry 2003, 54, 1118–1121. [Google Scholar] [CrossRef]
- Giacobbo, B.L.; Doorduin, J.; Klein, H.C.; Dierckx, R.A.J.O.; Bromberg, E.; de Vries, E.F.J. Brain-derived neurotrophic factor in brain disorders: Focus on neuroinflammation. Mol. Neurobiol. 2019, 56, 3295–3312. [Google Scholar] [CrossRef] [PubMed]
- Camuso, S.; La Rosa, P.; Fiorenza, M.T.; Canterini, S. Pleiotropic effects of BDNF on the cerebellum and hippocampus: Implications for neurodevelopmental disorders. Neurobiol. Dis. 2021, 163, 105606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lucassen, P.J.; Salta, E.; Verhaert, P.D.; Swaab, D.F. Hippocampal neuropathology in suicide: Gaps in our knowledge and opportunities for a breakthrough. Neurosci. Biobehav. Rev. 2021, 132, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Łoś, K.; Waszkiewicz, N. Biological Markers in Anxiety Disorders. J. Clin. Med. 2021, 10, 1744. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int. J. Mol. Sci. 2020, 21, 7777. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- De Vincenti, A.P.; Ríos, A.S.; Paratcha, G.; Ledda, F. Mechanisms That Modulate and Diversify BDNF Functions: Implications for Hippocampal Synaptic Plasticity. Front. Cell. Neurosci. 2019, 13, 135. [Google Scholar] [CrossRef]
- Marjańska, M.; McCarten, J.R.; Hodges, J.; Hemmy, L.S.; Grant, A.; Deelchand, D.K.; Terpstra, M. Region-specific aging of the human brain as evidenced by neurochemical profiles measured noninvasively in the posterior cingulate cortex and the occipital lobe using 1H magnetic resonance spectroscopy at 7 T. Neuroscience 2017, 354, 168–177. [Google Scholar] [CrossRef]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef]
- Choi, J.; Kwon, H.-J.; Lee, J.-E.; Lee, Y.; Seoh, J.-Y.; Han, P.-L. Hyperoxygenation revitalizes Alzheimer’s disease pathology through the upregulation of neurotrophic factors. Aging Cell 2019, 18, e12888. [Google Scholar] [CrossRef] [PubMed]
- Manukhina, E.B.; Tseilikman, V.E.; Karpenko, M.N.; Pestereva, N.S.; Tseilikman, O.B.; Komelkova, M.V.; Kondashevskaya, M.V.; Goryacheva, A.V.; Lapshin, M.S.; Platkovskii, P.O.; et al. Intermittent Hypoxic Conditioning Alleviates Post-Traumatic Stress Disorder-Induced Damage and Dysfunction of Rat Visceral Organs and Brain. Int. J. Mol. Sci. 2020, 21, 345. [Google Scholar] [CrossRef]
- Manukhina, E.B.; Tseilikman, V.E.; Komelkova, M.V.; Lapshin, M.S.; Goryacheva, A.V.; Kondashevskaya, M.V.; Mkhitarov, V.A.; Lazuko, S.S.; Tseilikman, O.B.; Sarapultsev, A.P.; et al. Cardiac injury in rats with experimental posttraumatic stress disorder and mechanisms of its limitation in experimental posttraumatic stress disorder-resistant rats. J. Appl. Physiol. 2021, 130, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.J.; Drayer, B.P.; Fram, E.K.; Williams, K.D.; Dumoulin, C.L.; Souza, S.P. MR angiography with two-dimensional acquisition and three-dimensional display. Work in progress. Radiology 1989, 173, 527–532. [Google Scholar] [CrossRef]
- Seryapina, A.A.; Shevelev, O.B.; Moshkin, M.P.; Markel, A.L.; Akulov, A.E. Stress-sensitive arterial hypertension, haemodynamic changes and brain metabolites in hypertensive ISIAH rats: MRI investigation. Exp. Physiol. 2017, 102, 523–532. [Google Scholar] [CrossRef]
- Naumova, A.V.; Akulov, A.E.; Khodanovich, M.Y.; Yarnykh, V.L. High-resolution three-dimensional quantitative map of the macromolecular proton fraction distribution in the normal rat brain. Data Brief 2016, 10, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef]
- Menshanov, P.N.; Akulov, A.E. The neurochemical profile of the hippocampus in isoflurane-treated and unanesthetized rat pups. Interdiscip. Toxicol. 2015, 8, 113–117. [Google Scholar] [CrossRef]
- Volchegorskii, I.A.; Rassokhina, L.M.; Miroshnichenko, I.Y. Dynamics of lipid peroxidation-antioxidant defense system during alloxan diabetes in rats. Bull. Exp. Biol. Med. 2013, 155, 26–29. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 7) | PS (n = 18) | High Anxiety (n = 11) | Low Anxiety (n = 7) | |
|---|---|---|---|---|
| Time spent in open arms (s) | 45 ± 14 | 82 ± 9 | 60 ± 12 | 103 ± 7 * # |
| Time spent in closed arms (s) | 565 ± 14 | 518 ± 15 | 450 ± 14 | 497 ± 7 * # |
| Entries to open arms | 2.1 ± 0.8 | 2.5 ± 0.6 | 1.7 ± 0.2 | 4.4 ± 0.4 |
| Entries to closed arms | 12.9 ± 1.7 | 9.5 ± 2.01 | 7.6 ± 0.6 | 9.0 ± 0.7 |
| AI (anxiety index) | 0.89 ± 0.02 | 0.82 ± 0.27 | 0.86 ± 0.01 | 0.75 ± 0.03 ** # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseilikman, V.; Akulov, A.; Shevelev, O.; Khotskina, A.; Kontsevaya, G.; Moshkin, M.; Fedotova, J.; Pashkov, A.; Tseilikman, O.; Agletdinov, E.; et al. Paradoxical Anxiety Level Reduction in Animal Chronic Stress: A Unique Role of Hippocampus Neurobiology. Int. J. Mol. Sci. 2022, 23, 9151. https://doi.org/10.3390/ijms23169151
Tseilikman V, Akulov A, Shevelev O, Khotskina A, Kontsevaya G, Moshkin M, Fedotova J, Pashkov A, Tseilikman O, Agletdinov E, et al. Paradoxical Anxiety Level Reduction in Animal Chronic Stress: A Unique Role of Hippocampus Neurobiology. International Journal of Molecular Sciences. 2022; 23(16):9151. https://doi.org/10.3390/ijms23169151
Chicago/Turabian StyleTseilikman, Vadim, Andrey Akulov, Oleg Shevelev, Anna Khotskina, Galina Kontsevaya, Mikhail Moshkin, Julia Fedotova, Anton Pashkov, Olga Tseilikman, Eduard Agletdinov, and et al. 2022. "Paradoxical Anxiety Level Reduction in Animal Chronic Stress: A Unique Role of Hippocampus Neurobiology" International Journal of Molecular Sciences 23, no. 16: 9151. https://doi.org/10.3390/ijms23169151