
Age-Dependent Variations in Functional Quality and Proteomic Characteristics of Canine (Canis lupus familiaris) Epididymal Spermatozoa

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Epididymal Sperm Quality Assessment
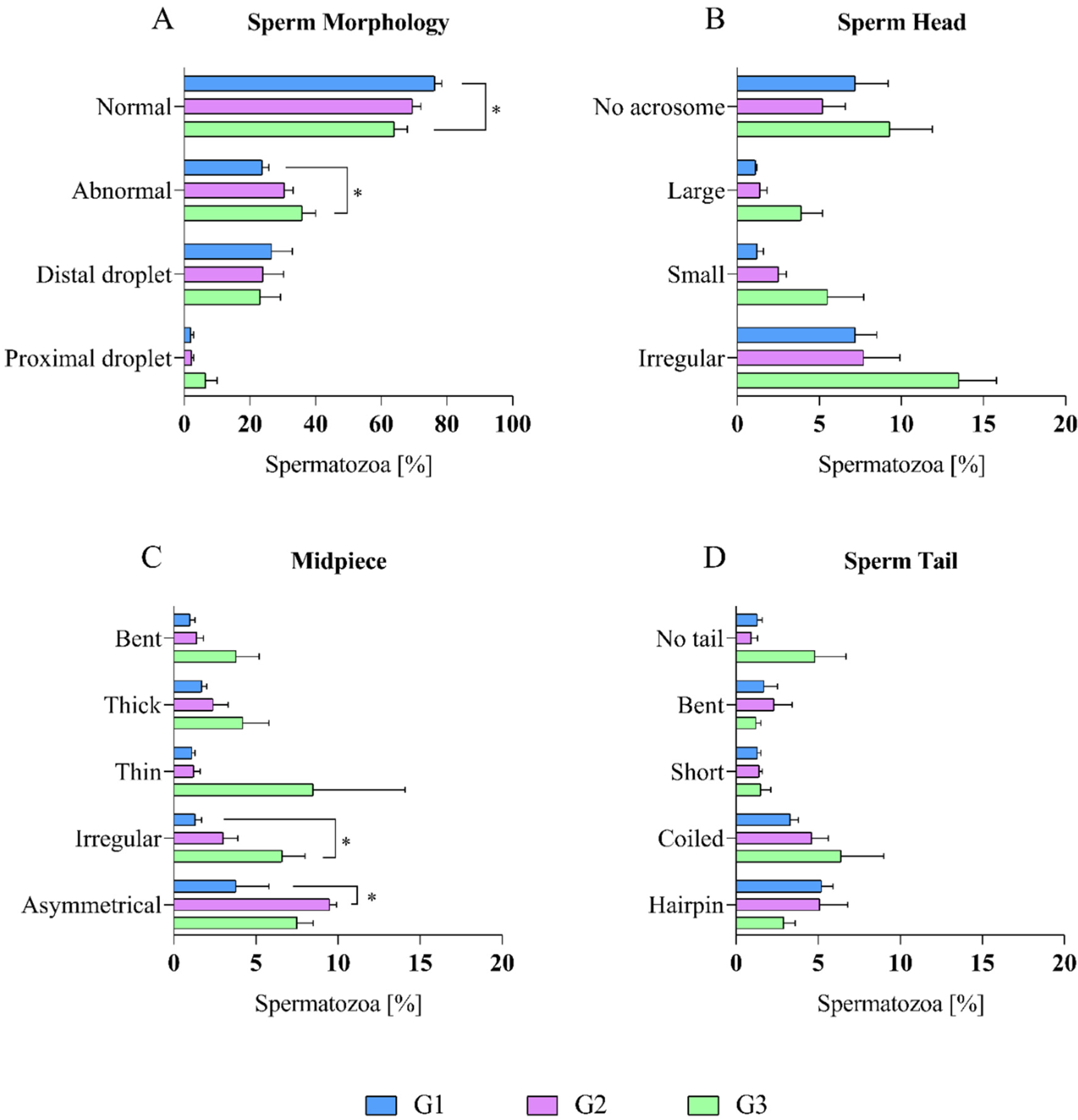
2.2. Epididymal Sperm Morphology Assessment
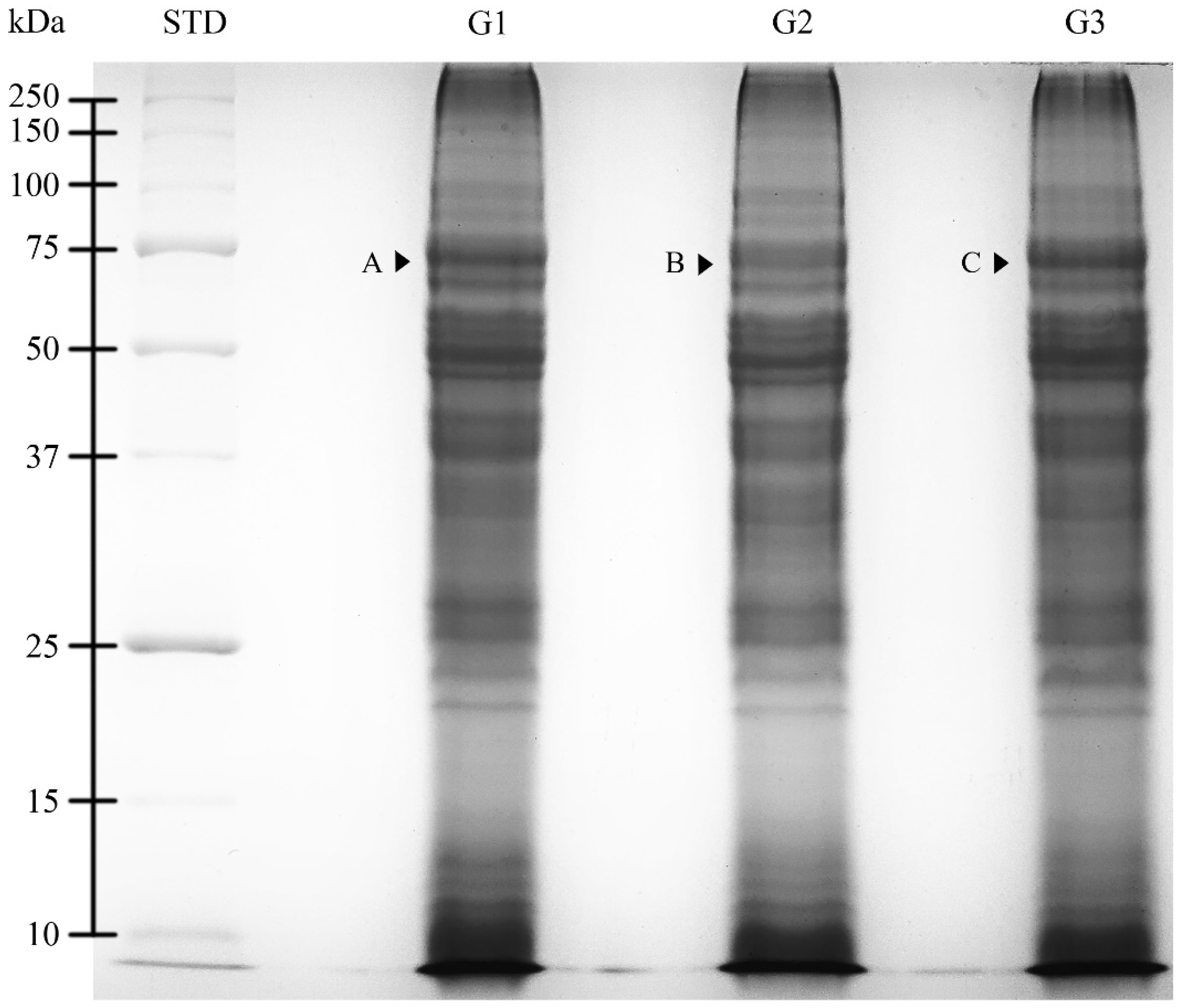
2.3. SDS-PAGE Analysis
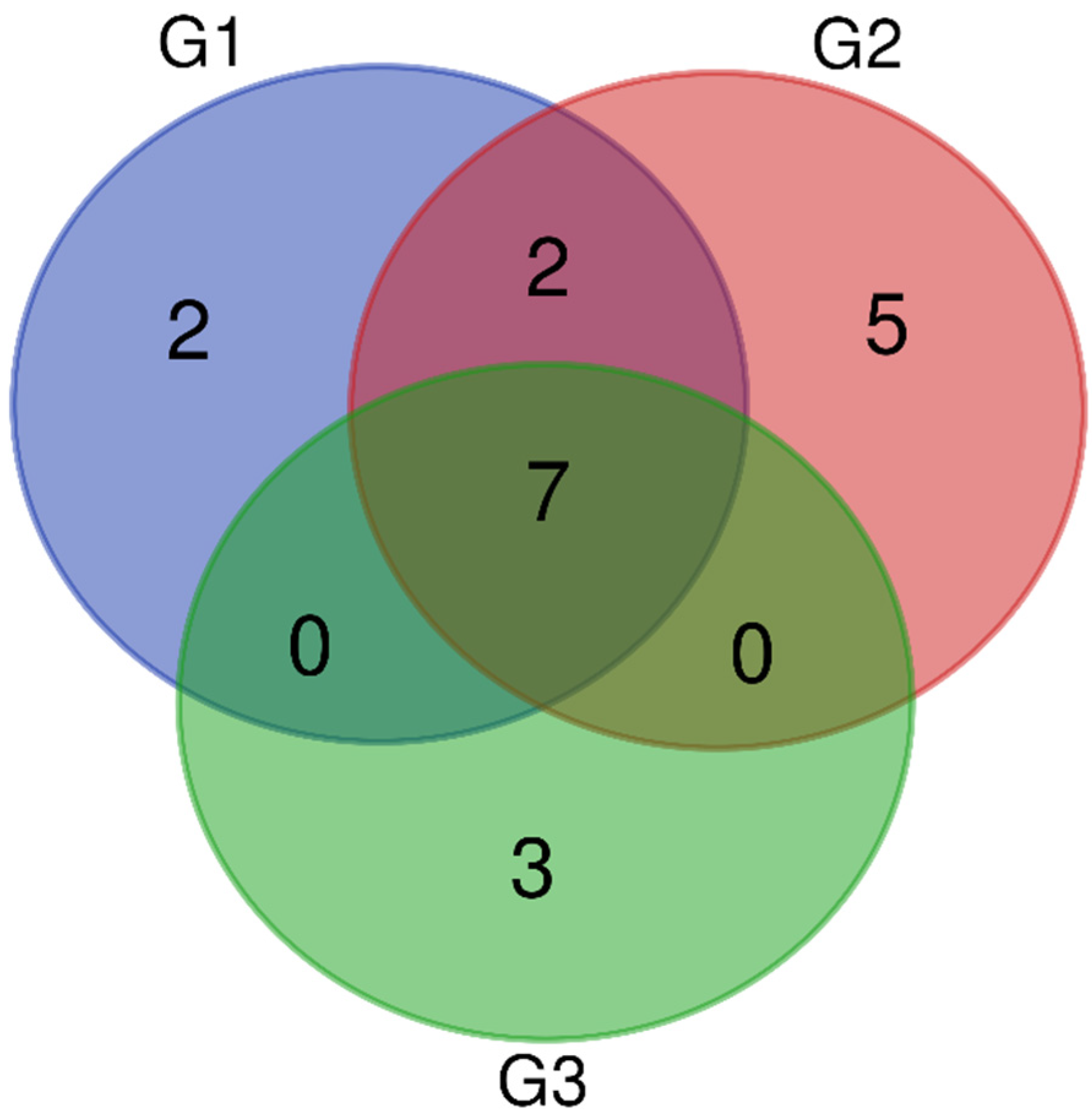
2.4. Qualitative and Quantitative Mass Spectrometry Analysis
2.4.1. Qualitative Analysis
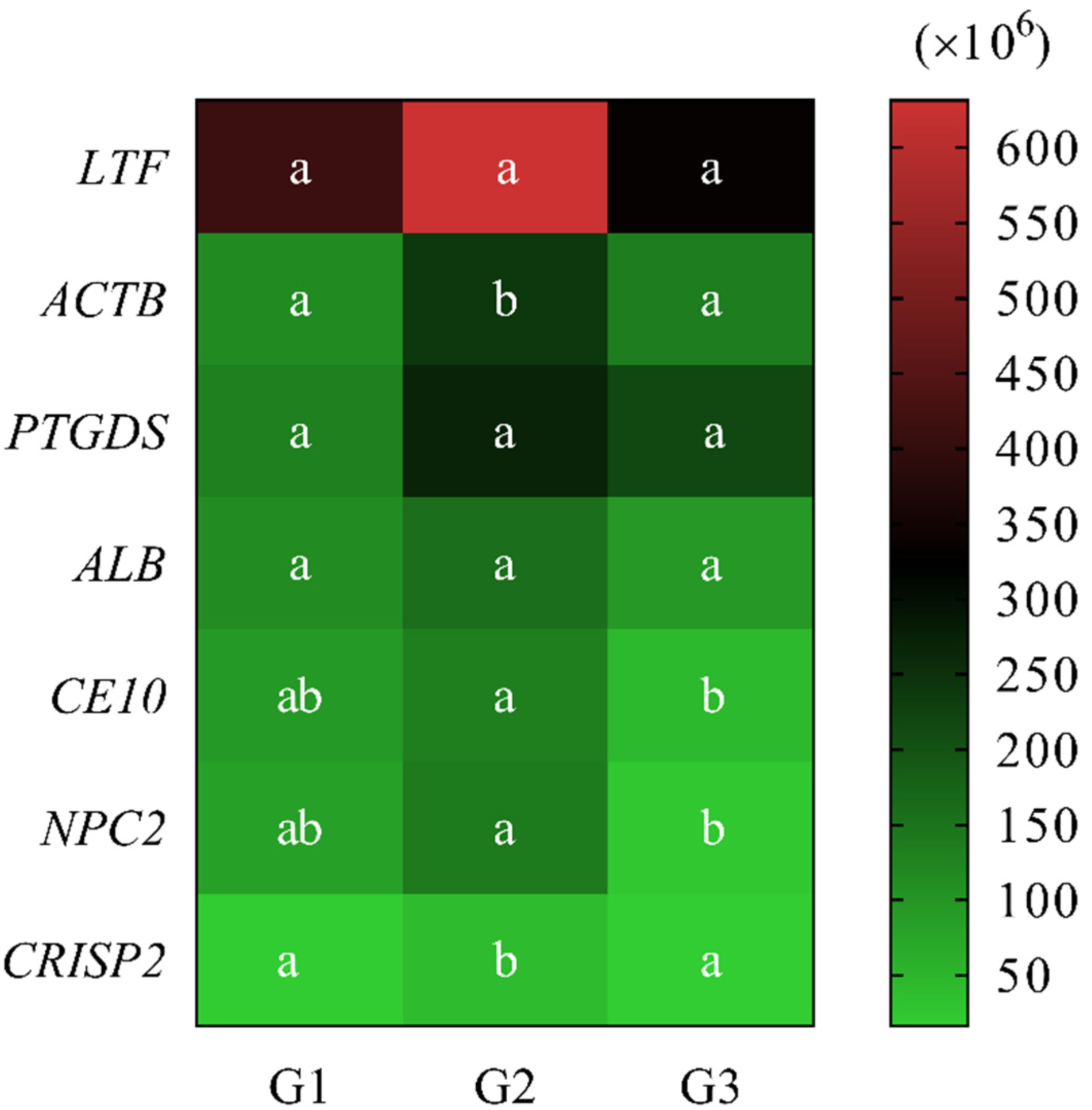
2.4.2. Quantitative Analysis
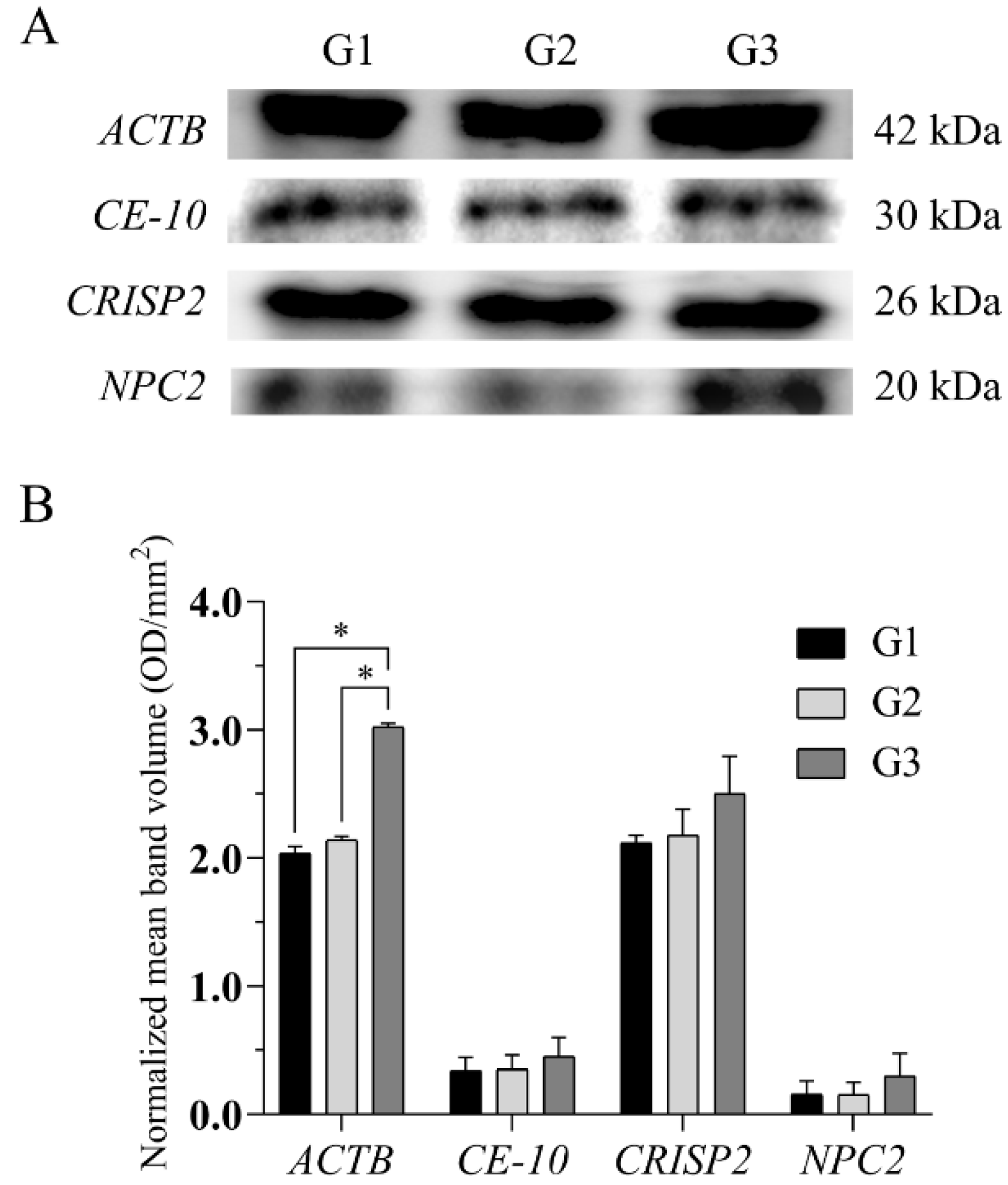
2.5. Western Blotting Analysis
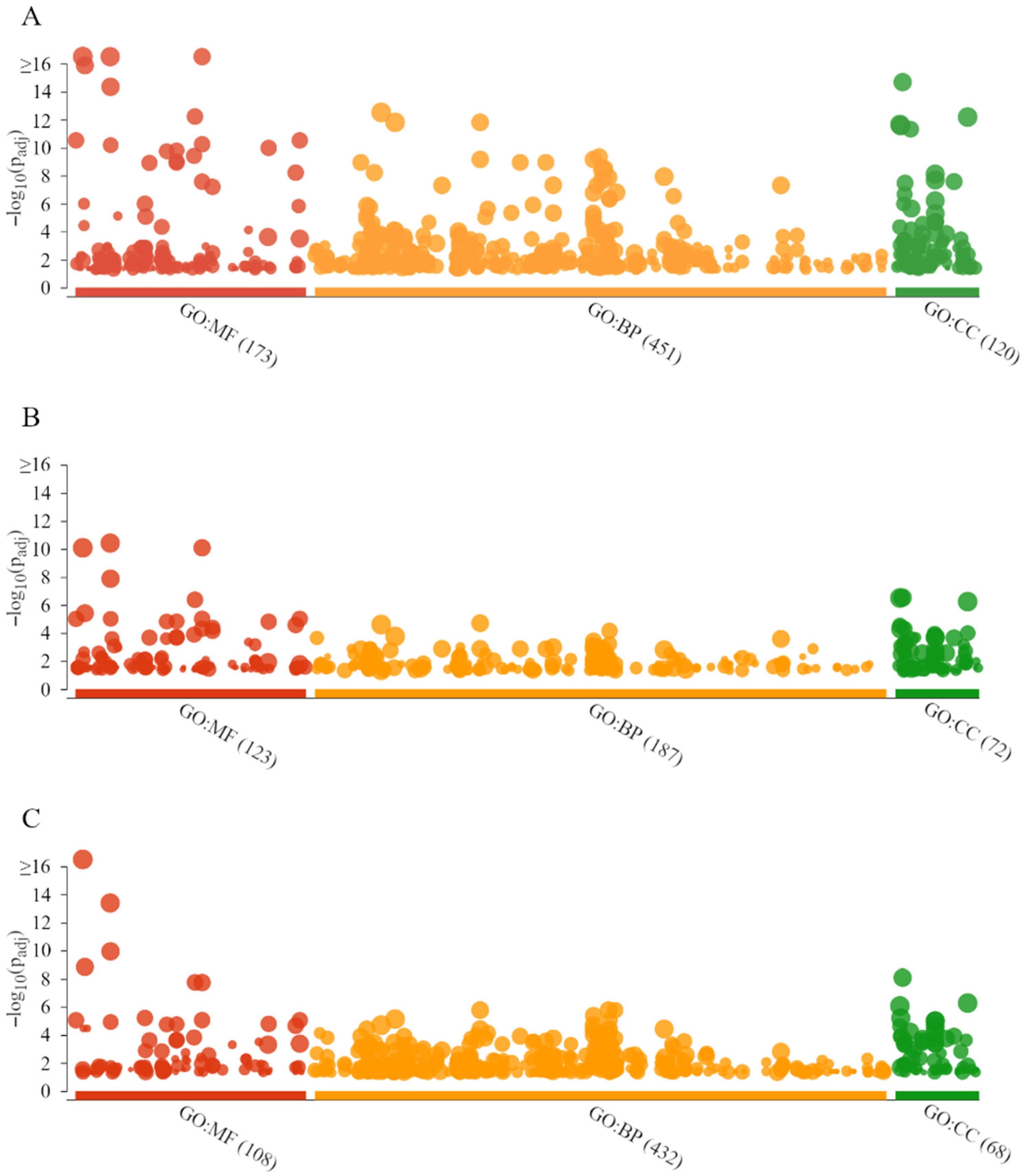
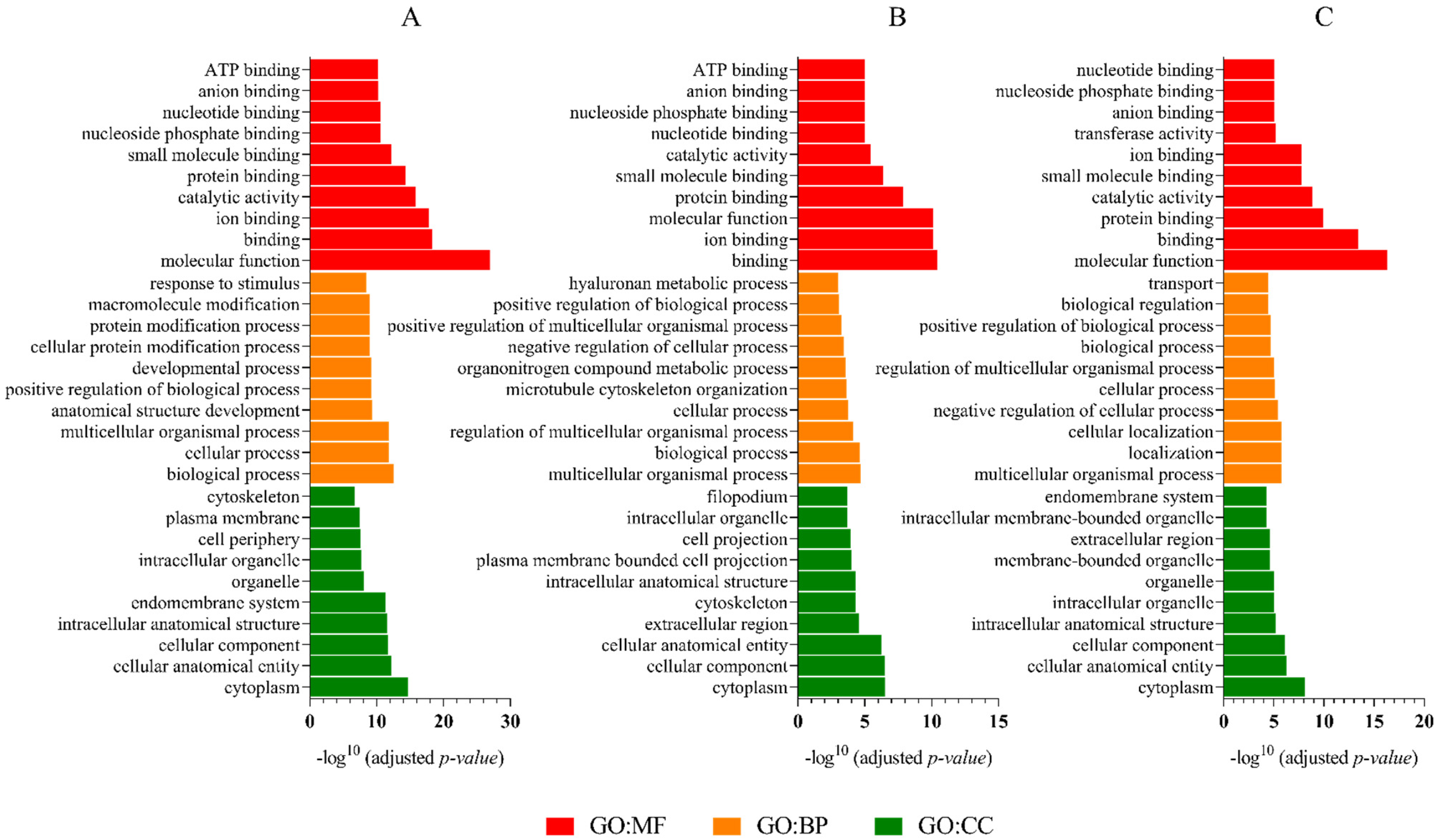
2.6. GO Analysis, KEGG Pathways, and Functional Annotation
3. Discussion
4. Materials and Methods
4.1. Chemicals and Media
4.2. Animals
4.3. Cauda Epididymal Semen Collection
4.4. Spermatozoa Quality Assessment
4.5. Morphology Assessment of Epididymal Spermatozoa
4.6. Preliminary Sample Preparation
4.6.1. Preliminary Sample Preparation for Protein Analysis
4.6.2. Total Protein Content Measurement
4.7. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.8. Western Blotting Analysis
4.9. Identification of Proteins in Liquid Samples by Mass Spectrometry
4.9.1. Sample Preparation for LC-MS Analysis
4.9.2. In Solution Digestion
4.9.3. NanoUPLC-Q-TOF/MS Analysis
4.10. Gene Ontology and Functional Annotation
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, H.A.; Domain, G.; Luvoni, G.C.; Chaaya, R.; Van Soom, A.; Wydooghe, E. Canine a/nd Feline Epididymal Semen—A Plentiful Source of Gametes. Animals 2021, 11, 2961. [Google Scholar] [CrossRef] [PubMed]
- Ellerbrock, K.; Pera, I.; Hartung, S.; Ivell, R. Gene expression in the dog epididymis: A model for human epididymal function. Int. J. Androl. 1994, 17, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, C. The dog as a model to study human epididymal function at a molecular level. Mol. Hum. Reprod. 2002, 8, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.V.; Cabral, M.; Correia, B.R.; Carvalho, P.; Sousa, M.; Oliveira, P.F.; Fardilha, M. mTOR Signaling Pathway Regulates Sperm Quality in Older Men. Cells 2019, 8, 629. [Google Scholar] [CrossRef]
- Rijsselaere, T.; Maes, D.; Hoflack, G.; De Kruif, A.; Van Soom, A. Effect of Body Weight, Age and Breeding History on Canine Sperm Quality Parameters Measured by the Hamilton-Thorne Analyser. Reprod. Domest. Anim. 2007, 42, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Rota, A.; Tesi, M.; Di Petta, G.; Sabatini, C.; Vannozzi, I. A retrospective study on the relationships between semen quality, dogs’ ageing and fertility. In Proceedings of the 8th International Symposium on Canine and Feline Reproduction with EVSSAR IXX, Paris, France, 22–25 June 2016. [Google Scholar]
- Bhanmeechao, C.; Srisuwatanasagul, S.; Prapaiwan, N.; Ponglowhapan, S. Reproductive aging in male dogs: The epididymal sperm defects and expression of androgen receptor in reproductive tissues. Theriogenology 2018, 108, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Leung, A.; Sinha-Hikim, A.P. Reproductive aging in the male brown-Norway rat: A model for the human. Endocrinology 1993, 133, 2773–2781. [Google Scholar] [CrossRef]
- Wright, W.W.; Fiore, C.; Zirkin, B.R. The Effect of Aging on the Seminiferous Epithelium of the Brown Norway Rat. J. Androl. 1993, 14, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.; Pastor, L.M.; Martínez, E.; Vázquez, J.M.; Roca, J. Age-related changes in the hamster epididymis. Anat. Rec. 1999, 256, 335–346. [Google Scholar] [CrossRef]
- Wolf, K.N.; Wildt, D.E.; Vargas, A.; Marinari, P.E.; Kreeger, J.S.; Ottinger, M.A.; Howard, J.G. Age-Dependent Changes in Sperm Production, Semen Quality, and Testicular Volume in the Black-Footed Ferret (Mustela nigripes). Biol. Reprod. 2000, 63, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Elcock, L.H.; Schoning, P. Age-related changes in the cat testis and epididymis. Am. J. Vet. Res. 1984, 45, 2380–2384. [Google Scholar] [PubMed]
- Fraser, L.; Brym, P.; Pareek, C.S.; Mogielnicka-Brzozowska, M.; Paukszto, Ł.; Jastrzębski, J.P.; Wasilewska-Sakowska, K.; Mańkowska, A.; Sobiech, P.; Żukowski, K. Transcriptome analysis of boar spermatozoa with different freezability using RNA-Seq. Theriogenology 2020, 142, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Ferretti, F.; Masini, M.; Gualtieri, G.; Moretti, E. Influence of age on sperm characteristics evaluated by light and electron microscopies. Sci. Rep. 2021, 11, 4989. [Google Scholar] [CrossRef] [PubMed]
- Elzanaty, S.; Malm, J. Effects of Ejaculation-to-Analysis Delay on Levels of Markers of Epididymal and Accessory Sex Gland Functions and Sperm Motility. J. Androl. 2007, 28, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, E.T.; Lewis, S.E.M.; McNally, J.A.; Thompson, W. In vitro fertilization and pregnancy rates: The influence of sperm motility and morphology on IVF outcome. Fertil. Steril. 1998, 70, 305–314. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Gatti, J.L.; Dacheux, F. Contribution of epididymal secretory proteins for spermatozoa maturation. Microsc. Res. Tech. 2003, 61, 7–17. [Google Scholar] [CrossRef]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef]
- Belloc, S.; Hazout, A.; Zini, A.; Merviel, P.; Cabry, R.; Chahine, H.; Copin, H.; Benkhalifa, M. How to overcome male infertility after 40: Influence of paternal age on fertility. Maturitas 2014, 78, 22–29. [Google Scholar] [CrossRef]
- Rosiak-Gill, A.; Gill, K.; Jakubik, J.; Fraczek, M.; Patorski, L.; Gaczarzewicz, D.; Kurzawa, R.; Kurpisz, M.; Piasecka, M. Age-related changes in human sperm DNA integrity. Aging 2019, 11, 5399–5411. [Google Scholar] [CrossRef]
- Eskenazi, B.; Wyrobek, A.J.; Sloter, E.; Kidd, S.A.; Moore, L.; Young, S.; Moore, D. The association of age and semen quality in healthy men. Hum. Reprod. 2003, 18, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Levitas, E.; Lunenfeld, E.; Weisz, N.; Friger, M.; Potashnik, G. Relationship between age and semen parameters in men with normal sperm concentration: Analysis of 6022 semen samples. Andrologia 2007, 39, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Verón, G.L.; Tissera, A.D.; Bello, R.; Beltramone, F.; Estofan, G.; Molina, R.I.; Vazquez-Levin, M.H. Impact of age, clinical conditions, and lifestyle on routine semen parameters and sperm kinematics. Fertil. Steril. 2018, 110, 68–75. [Google Scholar] [CrossRef]
- Lechner, D.; Aurich, J.; Schäfer-Somi, S.; Aurich, C. Effects of age, size and season on cryotolerance of dog semen—A retrospective analysis. Anim. Reprod. Sci. 2022, 236, 106912. [Google Scholar] [CrossRef] [PubMed]
- du Plessis, S.S.; Kashou, A.H.; Benjamin, D.J.; Yadav, S.P.; Agarwal, A. Proteomics: A subcellular look at spermatozoa. Reprod. Biol. Endocrinol. 2011, 9, 36. [Google Scholar] [CrossRef]
- Zmudzinska, A.; Bromke, M.A.; Strzezek, R.; Zielinska, M.; Olejnik, B.; Mogielnicka-Brzozowska, M. Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation. Animals 2022, 12, 772. [Google Scholar] [CrossRef]
- Baker, M.A.; Witherdin, R.; Hetherington, L.; Cunningham-Smith, K.; Aitken, R.J. Identification of post-translational modifications that occur during sperm maturation using differences in two-dimensional gel electrophoresis. Proteomics 2005, 5, 1003–1012. [Google Scholar] [CrossRef]
- Asano, A.; Selvaraj, V.; Buttke, D.E.; Nelson, J.L.; Green, K.M.; Evans, J.E.; Travis, A.J. Biochemical characterization of membrane fractions in murine sperm: Identification of three distinct sub-types of membrane rafts. J. Cell. Physiol. 2009, 218, 537–548. [Google Scholar] [CrossRef]
- Baker, M.A.; Hetherington, L.; Reeves, G.; Müller, J.; Aitken, R.J. The rat sperm proteome characterized via IPG strip prefractionation and LC-MS/MS identification. Proteomics 2008, 8, 2312–2321. [Google Scholar] [CrossRef]
- Baker, M.A.; Hetherington, L.; Reeves, G.M.; Aitken, R.J. The mouse sperm proteome characterized via IPG strip prefractionation and LC-MS/MS identification. Proteomics 2008, 8, 1720–1730. [Google Scholar] [CrossRef]
- Araujo, M.S.; Oliveira Henriques Paulo, O.L.D.; Paranzini, C.S.; Scott, C.; Codognoto, V.M.; de Paula Freitas Dell’Aqua, C.; Papa, F.O.; de Souza, F.F. Proteomic data of seminal plasma and spermatozoa of four purebred dogs. Data Brief 2020, 30, 105498. [Google Scholar] [CrossRef]
- Araujo, M.S.; Oliveira Henriques Paulo, O.L.D.; Scott, C.; Paranzini, C.S.; Codognoto, V.M.; de Paula Freitas Dell’Aqua, C.; Papa, F.O.; de Souza, F.F. Insights into the influence of canine breed on proteomics of the spermatozoa and seminal plasma. J. Proteom. 2022, 257, 104508. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.M.; López, M.L.; Ferreira, A.T.S.; Chapeaurouge, D.A.; Rodrigues, A.; Perales, J.; Retamal, C.A. Thiol-disulfide proteins of stallion epididymal spermatozoa. Anim. Reprod. Sci. 2014, 145, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Proteomic identification of cryostress in epididymal spermatozoa. J. Anim. Sci. Biotechnol. 2016, 7, 67. [Google Scholar] [CrossRef]
- Labas, V.; Spina, L.; Belleannee, C.; Teixeira-Gomes, A.P.; Gargaros, A.; Dacheux, F.; Dacheux, J.L. Analysis of epididymal sperm maturation by MALDI profiling and top-down mass spectrometry. J. Proteom. 2015, 15, 226–243. [Google Scholar] [CrossRef]
- Chauvin, T.; Xie, F.; Liu, T.; Nicora, C.D.; Yang, F.; Camp, D.G.; Smith, R.D.; Roberts, K.P. A systematic analysis of a deep mouse epididymal sperm proteome. Biol. Reprod. 2012, 87, 141. [Google Scholar] [CrossRef]
- Brito, M.M.; Angrimani, D.S.R.; Lucio, C.F.; Vannucchi, C.I. A case trial study of the effect of ageing on fresh and post-thaw sperm in dogs. Andrologia 2018, 50, e13123. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Hoffmann, E.K.; Mills, J.W. The cytoskeleton and cell volume regulation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 385–399. [Google Scholar] [CrossRef]
- Cabello-Agüeros, J.F.; Hernández-González, E.O.; Mújica, A. The role of F-actin cytoskeleton-associated gelsolin in the guinea pig capacitation and acrosome reaction. Cell Motil. Cytoskelet. 2003, 56, 94–108. [Google Scholar] [CrossRef]
- Breitbart, H.; Cohen, G.; Rubinstein, S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction 2005, 129, 263–268. [Google Scholar] [CrossRef]
- Naresh, S. Effect of cooling (4 °C) and cryopreservation on cytoskeleton actin and protein tyrosine phosphorylation in buffalo spermatozoa. Cryobiology 2016, 72, 7–13. [Google Scholar] [CrossRef]
- Pera, I.; Ivell, R.; Kirchhoff, C. Regional variation of gene expression in the dog epididymis as revealed by in-situ transcript hybridization. Int. J. Androl. 1994, 17, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Osterhoff, C.; Kirchhoff, C.; Krull, N.; Ivell, R. Molecular cloning and characterization of a novel human sperm antigen (HE2) specifically expressed in the proximal epididymis. Biol. Reprod. 1994, 50, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Naureckiene, S.; Sleat, D.E.; Lackland, H.; Fensom, A.; Vanier, M.T.; Wattiaux, R.; Jadot, M.; Lobel, P. Identification of HE1 as the second gene of Niemann-Pick C disease. Science 2000, 290, 2298–2301. [Google Scholar] [CrossRef] [PubMed]
- Jamsai, D.; Reilly, A.; Smith, S.J.; Gibbs, G.M.; Baker, H.W.; McLachlan, R.I.; de Kretser, D.M.; O’Bryan, M.K. Polymorphisms in the human cysteine-rich secretory protein 2 (CRISP2) gene in Australian men. Hum. Reprod. 2008, 23, 2151–2159. [Google Scholar] [CrossRef]
- Zhou, J.H.; Zhou, Q.Z.; Lyu, X.M.; Zhu, T.; Chen, Z.J.; Chen, M.K.; Xia, H.; Wang, C.Y.; Qi, T.; Li, X.; et al. The expression of cysteine-rich secretory protein 2 (CRISP2) and its specific regulator miR-27b in the spermatozoa of patients with asthenozoospermia. Biol. Reprod. 2015, 92, 28. [Google Scholar] [CrossRef]
- Gottschalk, M.; Metzger, J.; Martinsson, G.; Sieme, H.; Distl, O. Genome-wide association study for semen quality traits in GermanWarmblood stallions. Anim. Reprod. Sci. 2016, 171, 81–86. [Google Scholar] [CrossRef]
- Foster, J.A.; Gerton, G.L. Autoantigen 1 of the guinea pig sperm acrosome is the homologue of mouse Tpx-1 and human TPX1 and is a member of the cysteine-rich secretory protein (CRISP) family. Mol. Reprod. Dev. 1996, 44, 221–229. [Google Scholar] [CrossRef]
- O’Bryan, M.K.; Loveland, K.L.; Herszfeld, D.; McFarlane, J.R.; Hearn, M.T.; de Kretser, D.M. Identification of a rat testis-specific gene encoding a potential rat outer dense fibre protein. Mol. Reprod. Dev. 1998, 50, 313–322. [Google Scholar] [CrossRef]
- Reddy, T.; Gibbs, G.M.; Merriner, D.J.; Kerr, J.B.; O’Bryan, M.K. Cysteine-rich secretory proteins are not exclusively expressed in the male reproductive tract. Dev. Dyn. 2008, 237, 3313–3323. [Google Scholar] [CrossRef]
- Lim, S.; Kierzek, M.; O’Connor, A.E.; Brenker, C.; Merriner, D.J.; Okuda, H.; Volpert, M.; Gaikwad, A.; Bianco, D.; Potter, D.; et al. CRISP2 Is a Regulator of Multiple Aspects of Sperm Function and Male Fertility. Endocrinology 2019, 160, 915–924. [Google Scholar] [CrossRef]
- Gebhardt, K.; Ellerbrock, K.; Pera, I.; Ivell, R.; Kirchhoff, C. Differential expression of novel abundant and highly regionalized mRNAs of the canine epididymis. Reproduction 1999, 116, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, C.; Habben, I.; Ivell, R.; Krull, N. A Major Human Epididymis-Specific cDNA Encodes a Protein with Sequence Homology to Extracellular Proteinase Inhibitors. Biol. Reprod. 1991, 45, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Angrimani, D.S.R.; Losano, J.D.A.; Lucio, C.F.; Veiga, G.A.L.; Pereda, M.C.; Nichi, M.; Vannucchi, C.I. Role of residual cytoplasm on oxidative status during sperm maturation in dogs. Anim. Reprod. Sci. 2014, 151, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Michishita, E.; Garcés, G.; Barrett, J.C.; Horikawa, I. Upregulation of the KIAA1199 gene is associated with cellular mortality. Cancer Lett. 2006, 239, 71–77. [Google Scholar] [CrossRef]
- Chen, L.; Shi, K.; Andersen, T.L.; Qiu, W.; Kassem, M. KIAA1199 is a secreted molecule that enhances osteoblastic stem cell migration and recruitment. Cell Death Dis. 2019, 10, 126. [Google Scholar] [CrossRef]
- Suzuki, T.; Yano, K.; Sugimoto, S.; Kitajima, K.; Lennarz, W.J.; Inoue, S.; Inoue, Y.; Emori, Y. Endo-β-N-acetylglucosaminidase, an enzyme involved in the processing of free oligosaccharides in the cytosol. Proc. Natl. Acad. Sci. USA 2002, 99, 9691–9696. [Google Scholar] [CrossRef]
- Huang, C.; Harada, Y.; Hosomi, A.; Masahara-Negishi, Y.; Seino, J.; Fujihira, H.; Funakoshi, Y.; Suzuki, T.; Dohmae, N.; Suzuki, T. Endo-β-N-acetylglucosaminidase forms N-GlcNAc protein aggregates during ER-associated degradation in Ngly1-defective cells. Proc. Natl. Acad. Sci. USA 2015, 112, 1398–1403. [Google Scholar] [CrossRef]
- Saalmann, A.; Münz, S.; Ellerbrock, K.; Ivell, R.; Kirchhoff, C. Novel sperm-binding proteins of epididymal origin contain four fibronectin type II-modules. Mol. Reprod. Dev. 2001, 58, 88–100. [Google Scholar] [CrossRef]
- Ekhlasi-Hundrieser, M.; Schäfer, B.; Kirchhoff, C.; Hess, O.; Bellair, S.; Müller, P.; Töpfer-Petersen, E. Structural and molecular characterization of equine sperm-binding fibronectin-II module proteins. Mol. Reprod. Dev. 2005, 70, 45–57. [Google Scholar] [CrossRef]
- Schäfer, B.; von Horsten, H.; Dacheux, J.; Holtz, W.; Kirchhoff, C. Cloning and Characterization of Boar Epididymal Secretory Proteins by Homology to the Human. Reprod. Domest. Anim. 2003, 38, 111–118. [Google Scholar] [CrossRef]
- Ekhlasi-Hundrieser, M.; Schäfer, B.; Philipp, U.; Kuiper, H.; Leeb, T.; Mehta, M.; Kirchhoff, C.; Töpfer-Petersen, E. Sperm-binding fibronectin type II-module proteins are genetically linked and functionally related. Gene 2007, 392, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Petrunkina, A.M.; Ekhlasi-Hundrieser, M.; Hettel, C.; Waberski, D.; Harrison, R.A.P.; Töpfer-Petersen, E. Fibronectin type II-module proteins in the bovine genital tract and their putative role in cell volume control during sperm maturation. Reprod. Fertil. Dev. 2009, 21, 479–488. [Google Scholar] [CrossRef] [PubMed]
- D’Amours, O.; Frenette, G.; Fortier, M.; Leclerc, P.; Sullivan, R. Proteomic comparison of detergent-extracted sperm proteins from bulls with different fertility indexes. Reproduction 2010, 139, 545–556. [Google Scholar] [CrossRef] [PubMed]
- D’Amours, O.; Bordeleau, L.-J.; Frenette, G.; Blondin, P.; Leclerc, P.; Sullivan, R. Binder of sperm 1 and epididymal sperm binding protein 1 are associated with different bull sperm subpopulations. Reproduction 2012, 143, 759–771. [Google Scholar] [CrossRef]
- Cheng, G.-Z.; Li, J.-Y.; Li, F.; Wang, H.-Y.; Shi, G.-X. Human ribonuclease 9, a member of ribonuclease A superfamily, specifically expressed in the epididymis, is a novel sperm-binding protein. Asian J. Androl. 2009, 11, 240–251. [Google Scholar] [CrossRef]
- Liu, J.; Hou, Y.; Gong, L.; Gu, Y.; Duan, L.; Tang, Y.; Wang, W. A novel role for ribonuclease 9 in suppressing sperm capacitation and acrosome reaction. Science 2018, 44, 383–387. [Google Scholar] [CrossRef]
- Zhu, C.-F.; Liu, Q.; Zhang, L.; Yuan, H.-X.; Zhen, W.; Zhang, J.-S.; Chen, Z.-J.; Hall, S.H.; French, F.S.; Zhang, Y.-L. RNase9, an Androgen-Dependent Member of the RNase A Family, Is Specifically Expressed in the Rat Epididymis. Biol. Reprod. 2007, 76, 63–73. [Google Scholar] [CrossRef]
- Penttinen, J.; Pujianto, D.A.; Sipilä, P.; Huhtaniemi, I.; Poutanen, M. Discovery in Silico and Characterization in Vitro of Novel Genes Exclusively Expressed in the Mouse Epididymis. Mol. Endocrinol. 2003, 17, 2138–2151. [Google Scholar] [CrossRef]
- Fouchécourt, S.; Métayer, S.; Locatelli, A.; Dacheux, F.; Dacheux, J.-L. Stallion Epididymal Fluid Proteome: Qualitative and Quantitative Characterization; Secretion and Dynamic Changes of Major Proteins. Biol. Reprod. 2000, 62, 1790–1803. [Google Scholar] [CrossRef]
- Dacheux, J.L.; Belleannée, C.; Jones, R.; Labas, V.; Belghazi, M.; Guyonnet, B.; Druart, X.; Gatti, J.L.; Dacheux, F. Mammalian epididymal proteome. Mol. Cell. Endocrinol. 2009, 306, 45–50. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, J.K.; Edwards, C.A.; Xu, Z.; Taichman, R.; Wang, C.Y. Clusterin inhibits apoptosis by interacting with activated Bax. Nat. Cell Biol. 2005, 7, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.M.; Foster, D.N.; Crabo, B.G. Localization of clusterin on freeze-preserved bull spermatozoa before and after glass wool-sephadex filtration. J. Androl. 2001, 22, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Pawson, T. Protein modules and signalling networks. Nature 1995, 373, 573–580. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Atlas PLEKHH1. Available online: https://www.proteinatlas.org/ENSG00000054690-PLEKHH1/tissue (accessed on 20 June 2022).
- Cornwall, G.A.; Orgebin-Crist, M.C.; Hann, S.R. The CRES gene: A unique testis-regulated gene related to the cystatin family is highly restricted in its expression to the proximal region of the mouse epididymis. Mol. Endocrinol. 1992, 6, 1653–1664. [Google Scholar] [CrossRef]
- Sutton, H.G.; Fusco, A.; Cornwall, G.A. Cystatin-Related Epididymal Spermatogenic Protein Colocalizes with Luteinizing Hormone-β Protein in Mouse Anterior Pituitary Gonadotropes. Endocrinology 1999, 140, 2721–2732. [Google Scholar] [CrossRef]
- Yao, H.L.; Liu, M.; Wang, W.J.; Wang, X.L.; Song, J.; Song, Q.Q.; Han, J. Construction of miRNA-target networks using microRNA profiles of CVB3-infected HeLa cells. Sci. Rep. 2019, 9, 17876. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Atlas FAM135A. Available online: https://www.proteinatlas.org/ENSG00000082269-FAM135A/tissue (accessed on 20 June 2022).
- Kouprina, N.; Pavlicek, A.; Collins, N.K.; Nakano, M.; Noskov, V.N.; Ohzeki, J.-I.; Mochida, G.H.; Risinger, J.I.; Goldsmith, P.; Gunsior, M.; et al. The microcephaly ASPM gene is expressed in proliferating tissues and encodes for a mitotic spindle protein. Hum. Mol. Genet. 2005, 14, 2155–2165. [Google Scholar] [CrossRef]
- Lüers, G.H.; Michels, M.; Schwaab, U.; Franz, T. Murine calmodulin-binding protein 1 (Calmbp1): Tissue-specific expression during development and in adult tissues. Mech. Dev. 2002, 118, 229–232. [Google Scholar] [CrossRef]
- Mogielnicka-Brzozowska, M.; Kordan, W. Characteristics of selected seminal plasma proteins and their application in the improvement of the reproductive processes in mammals. Pol. J. Vet. Sci. 2011, 14, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Nyathi, Y.; Wilkinson, B.M.; Pool, M.R. Co-translational targeting and translocation of proteins to the endoplasmic reticulum. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2392–2402. [Google Scholar] [CrossRef]
- NagDas, S.K.; Winfrey, V.P.; Olson, G.E. Identification of ras and its downstream signaling elements and their potential role in hamster sperm motility. Biol. Reprod. 2002, 67, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-W.; Kim, S.-H.; Kim, D.-H.; Ha, J.J.; Yi, J.K.; Hwang, S.; Ryu, B.-Y.; Pang, M.-G.; Kwon, W.-S. Ras-related proteins (Rab) are key proteins related to male fertility following a unique activation mechanism. Reprod. Biol. 2019, 19, 356–362. [Google Scholar] [CrossRef]
- Bae, J.-W.; Yi, J.K.; Jeong, E.-J.; Lee, W.-J.; Hwang, J.-M.; Kim, D.-H.; Ha, J.J.; Kwon, W.-S. Ras-related proteins (Rab) play significant roles in sperm motility and capacitation status. Reprod. Biol. 2022, 22, 100617. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. Cell–cell interactions at the ectoplasmic specialization in the testis. Trends Endocrinol. Metab. 2004, 15, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.H.N.; Mruk, D.D.; Wong, E.W.P.; Lee, W.M.; Cheng, C.Y. An autocrine axis in the testis that coordinates spermiation and blood–testis barrier restructuring during spermatogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 8950–8955. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.T.; Ten Hagen, K.G. Mucin-type O-Glycosylation during Development. J. Biol. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef]
- An, G.; Wei, B.; Xia, B.; McDaniel, J.M.; Ju, T.; Cummings, R.D.; Braun, J.; Xia, L. Increased susceptibility to colitis and colorectal tumors in mice lacking core 3–derived O-glycans. J. Exp. Med. 2007, 204, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Liang, J.; Lv, C.; Memon, S.; Fang, Y.; Wu, G.; Quan, G. The characteristics of proteome and metabolome associated with contrasting sperm motility in goat seminal plasma. Sci. Rep. 2021, 11, 15562. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Castillo, J.; Ramalho-Santos, J.; Oliva, R. The combined human sperm proteome: Cellular pathways and implications for basic and clinical science. Hum. Reprod. Update 2014, 20, 40–62. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Pacheco, A.; Segura-Correa, J.C.; Bolio-Gonzalez, M.E.; Jiménez-Coello, M.; Linde Forsberg, C. Reproductive patterns of stray male dogs in the tropics. Theriogenology 2006, 66, 2084–2090. [Google Scholar] [CrossRef] [PubMed]
- Ramos Angrimani, D.S.; Nichi, M.; Losano, J.D.A.; Lucio, C.F.; Lima Veiga, G.A.; Junqueira, M.V.M.; Vannucchi, C.I. Fatty acid content in epididymal fluid and spermatozoa during sperm maturation in dogs. J. Animal Sci. Biotechnol. 2017, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Mogielnicka-Brzozowska, M.; Prochowska, S.; Niżański, W.; Bromke, M.A.; Wiśniewski, J.; Olejnik, B.; Kuzborska, A.; Fraser, L.; Młynarz, P.; Kordan, W. Proteome of cat semen obtained after urethral catheterization. Theriogenology 2020, 141, 68–81. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010; ISBN 978-92-4-154778-9. [Google Scholar]
- Cebi, C.; Faundez, R.; Jurka, P.; Akcay, E.; Petrajtis-Golobow, M.; Ambarcioglu, P. Evaluation of the canine epididymal sperm morphology with two different staining methods, one fixative solution and Motile Sperm Organelle Morphology Examination (MSOME). Kafkas Univ. Vet. Fak. Derg. 2016, 22, 57–62. [Google Scholar] [CrossRef]
- Intasqui, P.; Agarwal, A.; Sharma, R.; Samanta, L.; Bertolla, R.P. Towards the identification of reliable sperm biomarkers for male infertility: A sperm proteomic approach. Andrologia 2018, 50, e12919. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for an-notation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | G1 (n = 10) | G2 (n = 10) | G3 (n = 10) | p-Value |
|---|---|---|---|---|
| Concentration (×108/mL) | 38.2 ± 3.2 a | 27.7 ± 3.7 ab | 24.8 ± 3.9 b | 0.018 |
| Total motility (TMOT, %) | 92.8 ± 0.3 a | 89.1 ± 0.9 a | 81.3 ± 2.4 b | 0.002 |
| Progressive motility (PMOT, %) | 57.7 ± 2.7 a | 53.8 ± 2.3 a | 38.5 ± 5.8 b | 0.007 |
| Average path velocity (VAP, µm/s) | 137.7 ± 3.4 a | 132.5 ± 3.0 a | 113.5 ± 7.3 b | 0.002 |
| Straight line velocity (VSL, µm/s) | 114.0 ± 3.5 a | 110.1 ± 2.5 ab | 93.0 ± 7.5 b | 0.003 |
| Curvilinear velocity (VCL, µm/s) | 206.6 ± 5.9 a | 198.0 ± 8.0 ab | 174.2 ± 9.3 b | 0.014 |
| Amplitude of lateral head displacement (ALH, µm) | 7.1 ± 0.3 a | 6.8 ± 0.3 a | 6.5 ± 0.4 a | 0.430 |
| Beat cross frequency (BCF, Hz) | 16.2 ± 1.0 a | 17.2 ± 1.4 a | 19.8 ± 2.0 a | 0.115 |
| Straightness (STR, %) | 81.9 ± 1.3 a | 82.1 ± 1.1 a | 80.5 ± 2.2 a | 0.354 |
| Linearity (LIN, %) | 57.8 ± 2.3 a | 58.2 ± 2.2 a | 54.5 ± 3.4 a | 0.234 |
| Age Groups | Protein Bands | MW (kDa) | OD ± SE |
|---|---|---|---|
| G1 | A | 68.48 | 0.32 ± 0.01 a |
| G2 | B | 68.37 | 0.25 ± 0.01 b |
| G3 | C | 68.07 | 0.33 ± 0.02 a |
| G1 | ||||
| ID | Pathway Name | Protein Counts | −Log10 (Adjusted p-Value) | Protein Names |
| cfa03060 | Protein export | 3 | 5.63 × 10−4 | SRP14|SEC61A1|SRPRA |
| cfa04141 | Protein processing in endoplasmic reticulum | 6 | 6.22 × 10−4 | TUSC3|SEC24D|MAPK10| RRBP1|SEC61A1|ERN1 |
| cfa04014 | Ras signaling pathway | 6 | 3.58 × 10−3 | RGL2|TIAM1|GRB2| MAPK10|ZAP70|ABL1 |
| cfa04930 | Type II diabetes mellitus | 3 | 3.59 × 10−3 | ABCC8|HK1|MAPK10 |
| cfa00330 | Arginine and proline metabolism | 3 | 3.80 × 10−3 | MAOB|PRODH2|NOS3 |
| cfa04979 | Cholesterol metabolism | 3 | 4.02 × 10−3 | LIPA|ABCG8|ABCA1 |
| cfa02010 | ABC transporters | 3 | 4.02 × 10−3 | ABCG8|ABCC8|ABCA1 |
| cfa04530 | Tight junction | 5 | 5.10 × 10−3 | TIAM1|SLC9A3R1|MAPK10| RAB13|ROCK2 |
| cfa04360 | Axon guidance | 5 | 6.14 × 10−3 | PLXNC1|ABL1|ENAH| ROCK2|ABLIM2 |
| cfa00360 | Phenylalanine metabolism | 2 | 6.20 × 10−3 | MAOB|IL4I1 |
| cfa05135 | Yersinia infection | 4 | 7.34 × 10−3 | ROCK2|MAPK10|ZAP70|NLRP3 |
| G2 | ||||
| ID | Pathway Name | Protein Counts | −Log10 (Adjusted p-Value) | Protein Names |
| cfa05132 | Salmonella infection | 3 | 5.55 × 10−4 | NFKB1|TJP1|DYNC2H1 |
| cfa05202 | Transcriptional misregulation in cancer | 3 | 4.54 × 10−3 | MET|MLLT1|NFKB1 |
| cfa04520 | Adherens junction | 2 | 8.47 × 10−3 | MET|TJP1 |
| G3 | ||||
| ID | Pathway Name | Protein Counts | −Log10 (Adjusted p-Value) | Protein Names |
| cfa00512 | Mucin-type O-glycan biosynthesis | 2 | 3.37 × 10−3 | GALNT6|GALNT18 |
| cfa00860 | Porphyrin and chlorophyll metabolism | 2 | 3.79 × 10−3 | BLVRA|CP |
| cfa04110 | Cell cycle | 3 | 4.43 × 10−3 | SMC1B|ANAPC11|MCM2 |
| cfa04120 | Ubiquitin mediated proteolysis | 3 | 6.18 × 10−3 | ANAPC11|MGRN1|UBR5 |
| cfa02010 | ABC transporters | 2 | 7.61 × 10−3 | ABCA12|ABCC1 |
| cfa05225 | Hepatocellular carcinoma | 3 | 9.79 × 10−3 | PLCG2|TERT|WNT6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zmudzinska, A.; Wisniewski, J.; Mlynarz, P.; Olejnik, B.; Mogielnicka-Brzozowska, M. Age-Dependent Variations in Functional Quality and Proteomic Characteristics of Canine (Canis lupus familiaris) Epididymal Spermatozoa. Int. J. Mol. Sci. 2022, 23, 9143. https://doi.org/10.3390/ijms23169143
Zmudzinska A, Wisniewski J, Mlynarz P, Olejnik B, Mogielnicka-Brzozowska M. Age-Dependent Variations in Functional Quality and Proteomic Characteristics of Canine (Canis lupus familiaris) Epididymal Spermatozoa. International Journal of Molecular Sciences. 2022; 23(16):9143. https://doi.org/10.3390/ijms23169143
Chicago/Turabian StyleZmudzinska, Anna, Jerzy Wisniewski, Piotr Mlynarz, Beata Olejnik, and Marzena Mogielnicka-Brzozowska. 2022. "Age-Dependent Variations in Functional Quality and Proteomic Characteristics of Canine (Canis lupus familiaris) Epididymal Spermatozoa" International Journal of Molecular Sciences 23, no. 16: 9143. https://doi.org/10.3390/ijms23169143