
Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Identification of Several Inhibitors Targeting the IgE-Mediated Activation of MCs from More Than 60 Types of Compounds
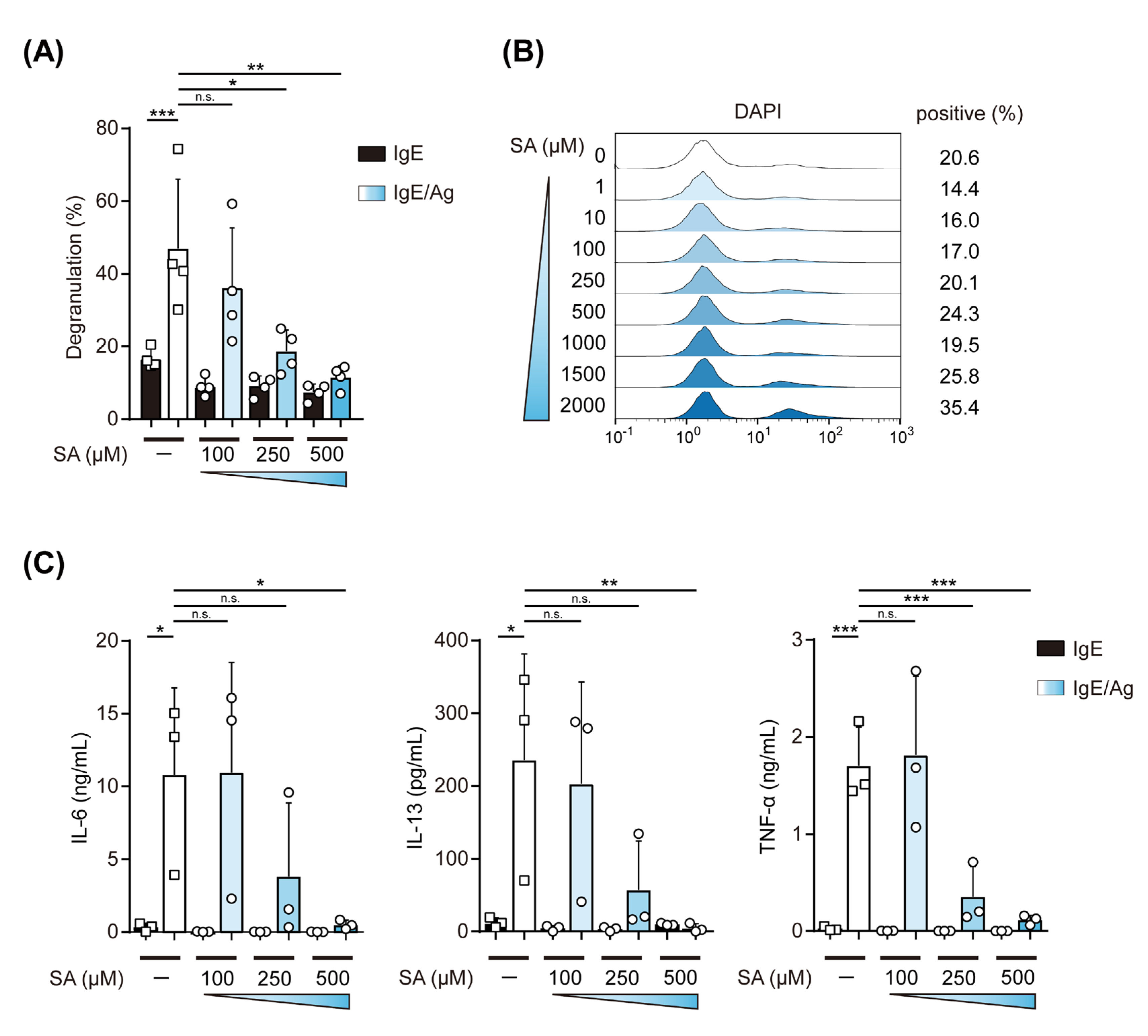
2.2. Effects of Salicylaldehyde on the IgE-Induced Degranulation of and Cytokine Production by MCs
2.3. Salicylaldehyde Suppressed the Expression of FcεRI and the IgE-Mediated Phosphorylation of Kinase Proteins in MC
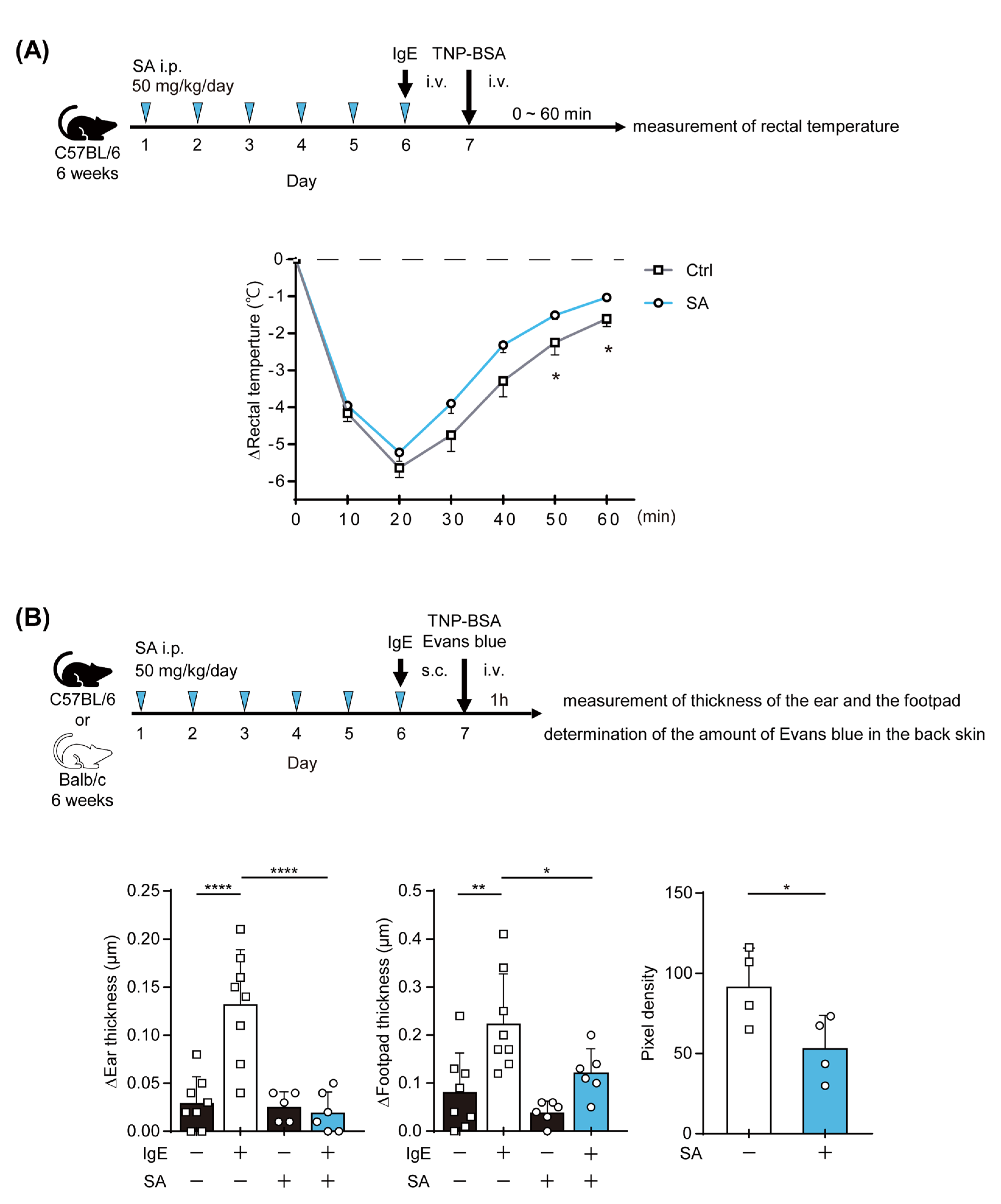
2.4. The Salicylaldehyde Treatment Ameliorated IgE-Mediated Anaphylaxis in Mice
3. Discussion
4. Materials and Methods
4.1. Mice and Cells
4.2. IgE-Induced Degranulation of MCs
4.3. Flow Cytometry
4.4. ELISA
4.5. Quantification of mRNA
4.6. Western Blot Analysis
4.7. PSA and PCA
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kubo, M. Mast cells and basophils in allergic inflammation. Curr. Opin. Immunol. 2018, 54, 74–79. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, U.; Huang, H.; Kawakami, T. The high affinity IgE receptor: A signaling update. Curr. Opin. Immunol. 2021, 72, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Okayama, Y.; Matsumoto, H.; Odajima, H.; Takahagi, S.; Hide, M.; Okubo, K. Roles of omalizumab in various allergic diseases. Allergol. Int. 2020, 69, 167–177. [Google Scholar] [CrossRef]
- Trinh, H.K.T.; Lee, S.H.; Cao, T.B.T.; Park, H.S. Asthma pharmacotherapy: An update on leukotriene treatments. Exp. Rev. Respir. Med. 2019, 13, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, L.; Jiang, J.; Li, J.; Xu, C.; Jin, S.; Zhu, L.; Yan, G. Pterostilbene Inhibits FcεRI Signaling through Activation of the LKB1/AMPK Pathway in Allergic Response. J. Agric. Food Chem. 2020, 68, 3456–3465. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Hu, S.; Ge, S.; Jia, M.; Wang, N. Resveratrol inhibits MRGPRX2-mediated mast cell activation via Nrf2 pathway. Int. Immunopharmacol. 2021, 93, 107426. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Yamamoto, M.; Inagaki, N.; Kitaura, J.; Chikumoto, T.; Kawahara, H.; Kawakami, Y.; Sano, M.; Miyase, T.; Tachibana, H.; Nagai, H.; et al. O-methylated catechins from tea leaves inhibit multiple protein kinases in mast cells. J. Immunol. 2004, 172, 4486–4492. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Lantz, C.S.; Oettgen, H.C.; Katona, I.M.; Fleming, T.; Miyajima, I.; Kinet, J.P.; Galli, S.J. IgE enhances mouse mast cell Fc(epsilon)RI expression in vitro and in vivo: Evidence for a novel amplification mechanism in IgE-dependent reactions. J. Exp. Med. 1997, 185, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Boesiger, J.; Tsai, M.; Maurer, M.; Yamaguchi, M.; Brown, L.F.; Claffey, K.P.; Dvorak, H.F.; Galli, S.J. Mast cells can secrete vascular permeability factor/ vascular endothelial cell growth factor and exhibit enhanced release after immunoglobulin E-dependent upregulation of fc epsilon receptor I expression. J. Exp. Med. 1998, 188, 1135–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Routy, B.; Gopalakrishnan, V.; Daillère, R.; Zitvogel, L.; Wargo, J.A.; Kroemer, G. The gut microbiota influences anticancer immunosurveillance and general health. Nat. Rev. Clin. Oncol. 2018, 15, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Holvoet, S.; Mercenier, A. Dietary polyphenols in the prevention and treatment of allergic diseases. Clin. Exp. Allergy 2011, 41, 1346–1359. [Google Scholar] [CrossRef] [PubMed]
- Hagenlocher, Y.; Feilhauer, K.; Schäffer, M.; Bischoff, S.C.; Lorentz, A. Citrus peel polymethoxyflavones nobiletin and tangeretin suppress LPS- and IgE-mediated activation of human intestinal mast cells. Eur. J. Nutr. 2017, 56, 1609–1620. [Google Scholar] [CrossRef]
- Kim, J.H.; Campbell, B.C.; Mahoney, N.; Chan, K.L.; Molyneux, R.J. Chemosensitization of aflatoxigenic fungi to antimycin A and strobilurin using salicylaldehyde, a volatile natural compound targeting cellular antioxidation system. Mycopathologia 2011, 171, 291–298. [Google Scholar] [CrossRef]
- Liu, C.; Liu, X.; Ge, X.; Wang, Q.; Zhang, L.; Shang, W.; Zhang, Y.; Yuan, X.A.; Tian, L.; Liu, Z.; et al. Fluorescent iridium(iii) coumarin-salicylaldehyde Schiff base compounds as lysosome-targeted antitumor agents. Dalton Trans. 2020, 49, 5988–5998. [Google Scholar] [CrossRef]
- Petrovic, Z.D.; Dorovic, J.; Simijonovic, D.; Petrovic, V.P.; Markovic, Z. Experimental and theoretical study of antioxidative properties of some salicylaldehyde and vanillic Schiff bases. Rsc. Adv. 2015, 5, 24094–24100. [Google Scholar] [CrossRef]
- Li, S.; Zhu, G.; Yang, Y.; Jian, Z.; Guo, S.; Dai, W.; Shi, Q.; Ge, R.; Ma, J.; Liu, L.; et al. Oxidative stress drives CD8. J. Allergy Clin. Immunol. 2017, 140, 177–189.e179. [Google Scholar] [CrossRef] [Green Version]
- Croft, A.; Tay, K.H.; Boyd, S.C.; Guo, S.T.; Jiang, C.C.; Lai, F.; Tseng, H.Y.; Jin, L.; Rizos, H.; Hersey, P.; et al. Oncogenic activation of MEK/ERK primes melanoma cells for adaptation to endoplasmic reticulum stress. J. Investig. Dermatol. 2014, 134, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Volkmann, K.; Lucas, J.L.; Vuga, D.; Wang, X.; Brumm, D.; Stiles, C.; Kriebel, D.; Der-Sarkissian, A.; Krishnan, K.; Schweitzer, C.; et al. Potent and selective inhibitors of the inositol-requiring enzyme 1 endoribonuclease. J. Biol. Chem. 2011, 286, 12743–12755. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Nishiyama, C.; Nishiyama, M.; Akizawa, Y.; Takahashi, K.; Ito, T.; Furukawa, S.; Ra, C.; Okumura, K.; Ogawa, H. Regulation of the human Fc epsilon RI alpha-chain distal promoter. J. Immunol. 2003, 170, 3732–3738. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kasakura, K.; Fujigaki, I.; Kageyama, A.; Okumura, K.; Ogawa, H.; Yashiro, T.; Nishiyama, C. The effect of PU.1 knockdown on gene expression and function of mast cells. Sci. Rep. 2018, 8, 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Hou, D.X. Multiple regulations of Keap1/Nrf2 system by dietary phytochemicals. Mol. Nutr. Food Res. 2016, 60, 1731–1755. [Google Scholar] [CrossRef] [PubMed]
- Kode, A.; Rajendrasozhan, S.; Caito, S.; Yang, S.R.; Megson, I.L.; Rahman, I. Resveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cells. Am. J. Physiol. Lung Cell. Mol. Phys. 2008, 294, L478–L488. [Google Scholar] [CrossRef] [Green Version]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.G.; Chen, T.Y.; Fahey, J.W.; Talalay, P. Keap1-nrf2 signaling: A target for cancer prevention by sulforaphane. Top. Curr. Chem. 2013, 329, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Eisenstein, A.; Hilliard, B.K.; Pope, S.D.; Zhang, C.; Taskar, P.; Waizman, D.A.; Israni-Winger, K.; Tian, H.; Luan, H.H.; Wang, A. Activation of the transcription factor NRF2 mediates the anti-inflammatory properties of a subset of over-the-counter and prescription NSAIDs. Immunity 2022, 55, 1082–1095.e1085. [Google Scholar] [CrossRef]
- Kasakura, K.; Nagata, K.; Miura, R.; Iida, M.; Nakaya, H.; Okada, H.; Arai, T.; Kawakami, Y.; Kawakami, T.; Yashiro, T.; et al. Cooperative Regulation of the Mucosal Mast Cell-Specific Protease Genes. J. Immunol. 2020, 204, 1641–1649. [Google Scholar] [CrossRef]
- Inage, E.; Kasakura, K.; Yashiro, T.; Suzuki, R.; Baba, Y.; Nakano, N.; Hara, M.; Tanabe, A.; Oboki, K.; Matsumoto, K.; et al. Critical Roles for PU.1, GATA1, and GATA2 in the expression of human FcεRI on mast cells: PU.1 and GATA1 transactivate FCER1A, and GATA2 transactivates FCER1A and MS4A2. J. Immunol. 2014, 192, 3936–3946. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, N.; Yokoyama, H.; Yashiro, T.; Nakano, N.; Nishiyama, M.; Kanada, S.; Fukai, T.; Hara, M.; Ikeda, S.; Ogawa, H.; et al. Role of PU.1 in MHC class II expression through transcriptional regulation of class II transactivator pI in dendritic cells. J. Allergy Clin. Immunol. 2012, 129, 814–824.e816. [Google Scholar] [CrossRef]
- Nakamura, Y.; Harama, D.; Shimokawa, N.; Hara, M.; Suzuki, R.; Tahara, Y.; Ishimaru, K.; Katoh, R.; Okumura, K.; Ogawa, H.; et al. Circadian clock gene Period2 regulates a time-of-day-dependent variation in cutaneous anaphylactic reaction. J. Allergy Clin. Immunol. 2011, 127, 1038–1045.E3. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | 0.01% | 0.001% | No. | Compound | 0.01% | 0.001% |
|---|---|---|---|---|---|---|---|
| 1 | Furfural | − | 33 | Anisole | − | ||
| 2 | 2-Piperonylpropanal | X | − | 34 | Benzaldehyde propylene glycol acetal | − | |
| 3 | Salicylaldehyde | X | + | 35 | Β-Caryophyllene | X | − |
| 4 | Phenylmethanol (benzylalchol) | − | 36 | p-Cymene | − | ||
| 5 | Furfuryl alcohol | X | − | 37 | Diacetyl | X | + |
| 6 | 2-Acetyl-3-octanone | − | 38 | 1.8-Cineole | − | ||
| 7 | Triacetin | − | 39 | Acetic acid | − | ||
| 8 | Isobutyl acetate | − | 40 | Hexanoic acid | − | ||
| 9 | Butyl butyryllactate | − | 41 | Hydroxycitronellal diethyl acetal | − | ||
| 10 | Diethyl succinate | − | 42 | Levulinic acid | − | ||
| 11 | δ-Dodecalactone | − | 43 | α-Pinene | X | − | |
| 12 | Ethyl acetoacetate | − | 44 | Terpinolene | − | ||
| 13 | Ethyl dodecanoate | − | 45 | 2-Pentylfuran | − | ||
| 14 | Ethyl nonanoate | − | 46 | Bisabolene | X | − | |
| 15 | Ambrettolide | + | 47 | Hexanal propylene glycol acetal | − | ||
| 16 | Methyl phenylacetate | − | 48 | 1.4-Cineole | − | ||
| 17 | Triethyl citrate | − | 49 | Isovaleraldehyde diethyl acetal | − | ||
| 18 | γ-Pentlactone | − | 50 | Carvacrol | X | − | |
| 19 | Methyl epi-dihydrojasmonate | − | 51 | 4-Ethylguaiacol | X | − | |
| 20 | Hexyl phenylacetate | − | 52 | Nerolidol | X | − | |
| 21 | Ethylene brassylate | − | 53 | Hotrienol | − | ||
| 22 | Cis-3-Hexenyllactate | − | 54 | α-Hexylcinnamaldehyde | X | − | |
| 23 | Cis-3-Hexenylpyruvate | − | 55 | trans-2-Heptenal | X | X | |
| 24 | Methyl 3-hydroxybutyrate | X | − | 56 | 5-Methyl-2-phenyl-2-hexenal | X | X |
| 25 | Isoamyl salicylate | + | 57 | 2-Phenylcrotonaldehyde | X | − | |
| 26 | Benzyl salicylate | + | 58 | 2-Isopropyl-5-methyl-2-hexenal | − | ||
| 27 | 2-Hydroxy-3-methyl-2-hexen-1,4-olide | − | 59 | Linalool-3.6-oxide | − | ||
| 28 | Allyl cinnamate | − | 60 | α-Irone | X | X | |
| 29 | Allyl phenoxyacetate | − | 61 | cis-Jasmone | + | ||
| 30 | Furaneol acetate | − | 62 | 4-Oxoisophorone | − | ||
| 31 | Mintlactone | + | 63 | Mesifurane | − | ||
| 32 | Isoamyl alcohol | − |
| Compound | MW | v/v (%) | Concentration (μM) |
|---|---|---|---|
| Salicylaldehyde | 122.1 | 0.001 | 95.8 |
| 0.002 | 191.6 | ||
| Diacetyl | 86.1 | 0.001 | 115.0 |
| 0.002 | 230.0 | ||
| Compound | MW | v/v (%) | Concentration (μM) |
| Ambrettolide | 252.4 | 0.002 | 75.8 |
| 0.010 | 378.8 | ||
| Isoamyl salicylate | 208.3 | 0.002 | 100.8 |
| 0.010 | 504.2 | ||
| Benzyl salicylate | 228.2 | 0.002 | 102.5 |
| 0.010 | 512.6 | ||
| Mintlactone | 166.2 | 0.002 | 127.3 |
| 0.010 | 636.5 | ||
| cis-Jasmone | 164.2 | 0.002 | 114.5 |
| 0.010 | 572.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashikari, T.; Hachisu, M.; Nagata, K.; Ando, D.; Iizuka, Y.; Ito, N.; Ito, K.; Ikeda, Y.; Matsubara, H.; Yashiro, T.; et al. Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice. Int. J. Mol. Sci. 2022, 23, 8826. https://doi.org/10.3390/ijms23158826
Ashikari T, Hachisu M, Nagata K, Ando D, Iizuka Y, Ito N, Ito K, Ikeda Y, Matsubara H, Yashiro T, et al. Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice. International Journal of Molecular Sciences. 2022; 23(15):8826. https://doi.org/10.3390/ijms23158826
Chicago/Turabian StyleAshikari, Tsubasa, Masakazu Hachisu, Kazuki Nagata, Daisuke Ando, Yuki Iizuka, Naoto Ito, Kandai Ito, Yuki Ikeda, Hiroki Matsubara, Takuya Yashiro, and et al. 2022. "Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice" International Journal of Molecular Sciences 23, no. 15: 8826. https://doi.org/10.3390/ijms23158826