
Angiotensin II Exaggerates SARS-CoV-2 Specific T-Cell Response in Convalescent Individuals following COVID-19

 , , ,
, , , 
Abstract
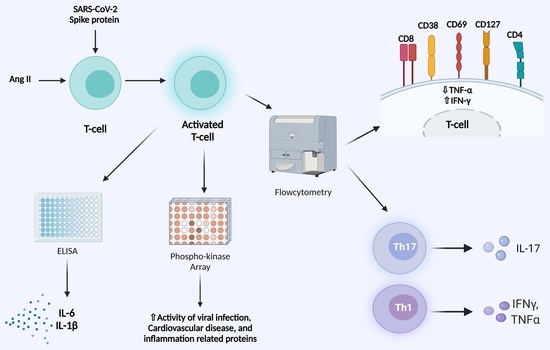
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Ang II Exaggerates SARS-CoV-2-Specific Memory T-cell Response in Recovered COVID-19 Patients
2.2. Ang II Alters the Inflammatory Response in Recovered COVID-19 Patients
2.3. Ang II Stimulates T-Helper Type1 (Th1) and Type17 (Th17) Phenotypes in Recovered COVID-19 Patients
2.4. Ang II Contributes to the Cytokine Storm Caused by SARS-CoV-2 Infection
2.5. Ang II Increases the Phosphorylation Level of Kinases in Recovered PBMCs
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.2. Peripheral Blood Mononuclear Cells (PBMCs) Isolation
4.3. T-Cell Stimulation and Treatment
4.4. Activation-Induced Cell Marker Assay (AIM)
4.5. Intracellular Cytokine Staining
4.6. Human Phospho-Kinase Array
4.7. Mitogenic Stimulation
4.8. ELISA Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Patidar, R.; Younis, K.; Desai, P.; Hosein, Z.; Padda, I.; Mangat, J.; Altaf, M. Comorbidity and Its Impact on Patients with COVID-19. SN Compr. Clin. Med. 2020, 2, 1069–1076. [Google Scholar] [CrossRef]
- Bosso, M.; Thanaraj, T.A.; Abu-Farha, M.; Alanbaei, M.; Abubaker, J.; Al-Mulla, F. The Two Faces of ACE2: The Role of ACE2 Receptor and Its Polymorphisms in Hypertension and COVID-19. Mol. Ther. Methods Clin. Dev. 2020, 18, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Tallant, E.A.; Clark, M.A. Molecular Mechanisms of Inhibition of Vascular Growth by Angiotensin-(1-7). Hypertension 2003, 42, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, I.O.; Melenotte, C.; Brouqui, P.; Million, M.; Lagier, J.C.; Parola, P.; Stein, A.; Scola, B.L.; Meddeb, L.; Mege, J.-L.; et al. Expression of ACE2 Receptor Soluble ACE2 Angiotensin I Angiotensin II and Angiotensin (1-7) Is Modulated in COVID-19 Patients. Front. Immunol. 2021, 12, 2350. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Y.; Zhang, C.; Huang, F.; Wang, F.; Yuan, J.; Wang, Z.; Li, J.; Li, J.; Feng, C.; et al. Clinical and Biochemical Indexes from 2019-NCoV Infected Patients Linked to Viral Loads and Lung Injury. Sci. China Life Sci. 2020, 63, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Almutlaq, M.; Alamro, A.A.; Alroqi, F.; Barhoumi, T. Classical and Counter-Regulatory Renin-Angiotensin System: Potential Key Roles in COVID-19 Pathophysiology. CJC Open 2021, 3, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Wiese, O.J.; Allwood, B.W.; Zemlin, A.E. COVID-19 and the Renin-Angiotensin System (RAS): A Spark That Sets the Forest Alight? Med. Hypotheses 2020, 144, 110231. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, T.; Kasal, D.A.; Li, M.W.; Shbat, L.; Laurant, P.; Neves, M.F.; Paradis, P.; Schiffrin, E.L. T Regulatory Lymphocytes Prevent Angiotensin II-Induced Hypertension and Vascular Injury. Hypertension 2011, 57, 469–476. [Google Scholar] [CrossRef] [Green Version]
- De Miguel, C.; Rudemiller, N.P.; Abais, J.M.; Mattson, D.L. Inflammation and Hypertension: New Understandings and Potential Therapeutic Targets. Curr. Hypertens. Rep. 2015, 17, 507. [Google Scholar] [CrossRef] [Green Version]
- Kanellakis, P.; Dinh, T.N.; Agrotis, A.; Bobik, A. CD4+CD25+Foxp3+ Regulatory T Cells Suppress Cardiac Fibrosis in the Hypertensive Heart. J. Hypertens. 2011, 29, 1820–1828. [Google Scholar] [CrossRef]
- Sahiratmadja, E.; Alisjahbana, B.; de Boer, T.; Adnan, I.; Maya, A.; Danusantoso, H.; Nelwan, R.H.H.; Marzuki, S.; van der Meer, J.W.M.; van Crevel, R.; et al. Dynamic Changes in Pro- and Anti-Inflammatory Cytokine Profiles and Gamma Interferon Receptor Signaling Integrity Correlate with Tuberculosis Disease Activity and Response to Curative Treatment. Infect. Immun. 2007, 75, 820–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seder, R.A.; Darrah, P.A.; Roederer, M. T-Cell Quality in Memory and Protection: Implications for Vaccine Design. Nat. Rev. Immunol. 2008, 8, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Thevarajan, I.; Nguyen, T.H.O.; Koutsakos, M.; Druce, J.; Caly, L.; van de Sandt, C.E.; Jia, X.; Nicholson, S.; Catton, M.; Cowie, B.; et al. Breadth of Concomitant Immune Responses Prior to Patient Recovery: A Case Report of Non-Severe COVID-19. Nat. Med. 2020, 26, 453–455. [Google Scholar] [CrossRef] [Green Version]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.; Lugli, E. The Who’s Who of T-Cell Differentiation: Human Memory T-Cell Subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef]
- Barata, J.T.; Durum, S.K.; Seddon, B. Flip the Coin: IL-7 and IL-7R in Health and Disease. Nat. Immunol. 2019, 20, 1584–1593. [Google Scholar] [CrossRef]
- Neidleman, J.; Luo, X.; Frouard, J.; Xie, G.; Gill, G.; Stein, E.S.; McGregor, M.; Ma, T.; George, A.F.; Kosters, A.; et al. SARS-CoV-2-Specific T Cells Exhibit Phenotypic Features of Helper Function, Lack of Terminal Differentiation, and High Proliferation Potential. Cell Rep. Med. 2020, 1, 100081. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wang, E.H.C.; Petukhova, L.; Chang, Y.; Lee, E.Y.; Christiano, A.M. Blockade of IL-7 Signaling Suppresses Inflammatory Responses and Reverses Alopecia Areata in C3H/HeJ Mice. Sci. Adv. 2021, 7, eabd1866. [Google Scholar] [CrossRef]
- Madhur, M.S.; Lob, H.E.; McCann, L.A.; Iwakura, Y.; Blinder, Y.; Guzik, T.J.; Harrison, D.G. Interleukin 17 Promotes Angiotensin II-Induced Hypertension and Vascular Dysfunction. Hypertension 2010, 55, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.N.; King, C.A.; Bailey, S.R.; Holt, J.W.; Venkatachalam, K.; Agrawal, A.; Valente, A.J.; Chandrasekar, B. Interleukin-17 Stimulates C-Reactive Protein Expression in Hepatocytes and Smooth Muscle Cells via P38 MAPK and ERK1/2-Dependent NF-KappaB and C/EBPbeta Activation. J. Biol. Chem. 2007, 282, 27229–27238. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; Cohen, S.L.; et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Parackova, Z.; Bloomfield, M.; Klocperk, A.; Sediva, A. Neutrophils Mediate Th17 Promotion in COVID-19 Patients. J. Leukoc. Biol. 2021, 109, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Matsuishi, Y.; Mathis, B.J.; Shimojo, N.; Subrina, J.; Okubo, N.; Inoue, Y. Severe COVID-19 Infection Associated with Endothelial Dysfunction Induces Multiple Organ Dysfunction: A Review of Therapeutic Interventions. Biomedicines 2021, 9, 279. [Google Scholar] [CrossRef]
- Guzik, T.J.; Hoch, N.E.; Brown, K.A.; McCann, L.A.; Rahman, A.; Dikalov, S.; Goronzy, J.; Weyand, C.; Harrison, D.G. Role of the T Cell in the Genesis of Angiotensin II Induced Hypertension and Vascular Dysfunction. J. Exp. Med. 2007, 204, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Smail, S.W.; Saeed, M.; Alkasalias, T.; Khudhur, Z.O.; Younus, D.A.; Rajab, M.F.; Abdulahad, W.H.; Hussain, H.I.; Niaz, K.; Safdar, M. Inflammation, Immunity and Potential Target Therapy of SARS-CoV-2: A Total Scale Analysis Review. Food Chem. Toxicol. 2021, 150, 112087. [Google Scholar] [CrossRef]
- Rowaiye, A.B.; Okpalefe, O.A.; Onuh Adejoke, O.; Ogidigo, J.O.; Hannah Oladipo, O.; Ogu, A.C.; Oli, A.N.; Olofinase, S.; Onyekwere, O.; Rabiu Abubakar, A.; et al. Attenuating the Effects of Novel COVID-19 (SARS-CoV-2) Infection-Induced Cytokine Storm and the Implications. J. Inflamm. Res. 2021, 14, 1487–1510. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.O. TH17 Responses in Cytokine Storm of COVID-19: An Emerging Target of JAK2 Inhibitor Fedratinib. J. Microbiol. Immunol. Infect. 2020, 53, 368–370. [Google Scholar] [CrossRef]
- Pacha, O.; Sallman, M.A.; Evans, S.E. COVID-19: A Case for Inhibiting IL-17? Nat. Rev. Immunol. 2020, 20, 345–346. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A. Heterogeneity of CD4+ Memory T Cells: Functional Modules for Tailored Immunity. Eur. J. Immunol. 2009, 39, 2076–2082. [Google Scholar] [CrossRef]
- Lane, N.; Robins, R.A.; Corne, J.; Fairclough, L. Regulation in Chronic Obstructive Pulmonary Disease: The Role of Regulatory T-Cells and Th17 Cells. Clin. Sci. 2010, 119, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, A.; Tahmasebi, S.; Mahmood, A.; Kuznetsova, M.; Valizadeh, H.; Taghizadieh, A.; Nazemiyeh, M.; Aghebati-Maleki, L.; Jadidi-Niaragh, F.; Abbaspour-Aghdam, S.; et al. Th17 and Treg Cells Function in SARS-CoV2 Patients Compared with Healthy Controls. J. Cell. Physiol. 2021, 236, 2829–2839. [Google Scholar] [CrossRef]
- Hausding, M.; Jurk, K.; Daub, S.; Kröller-Schön, S.; Stein, J.; Schwenk, M.; Oelze, M.; Mikhed, Y.; Kerahrodi, J.G.; Kossmann, S.; et al. CD40L Contributes to Angiotensin II-Induced pro-Thrombotic State, Vascular Inflammation, Oxidative Stress and Endothelial Dysfunction. Basic Res. Cardiol. 2013, 108, 386. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Dai, S.-M.; Tong, Q. COVID-19: A Recommendation to Examine the Effect of Hydroxychloroquine in Preventing Infection and Progression. J. Antimicrob. Chemother. 2020, 75, 1667–1670. [Google Scholar] [CrossRef]
- Kawabe, T.; Matsushima, M.; Hashimoto, N.; Imaizumi, K.; Hasegawa, Y. CD40/CD40 Ligand Interactions in Immune Responses and Pulmonary Immunity. Nagoya J. Med. Sci. 2011, 73, 69–78. [Google Scholar]
- Ara, A.; Ahmed, K.A.; Xiang, J. Multiple Effects of CD40-CD40L Axis in Immunity against Infection and Cancer. ImmunoTargets Ther. 2018, 7, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Tuazon Kels, M.J.; Ng, E.; Al Rumaih, Z.; Pandey, P.; Ruuls, S.R.; Korner, H.; Newsome, T.P.; Chaudhri, G.; Karupiah, G. TNF Deficiency Dysregulates Inflammatory Cytokine Production, Leading to Lung Pathology and Death during Respiratory Poxvirus Infection. Proc. Natl. Acad. Sci. USA 2020, 117, 15935–15946. [Google Scholar] [CrossRef] [PubMed]
- Chava, K.R.; Karpurapu, M.; Wang, D.; Bhanoori, M.; Kundumani-Sridharan, V.; Zhang, Q.; Ichiki, T.; Glasgow, W.C.; Rao, G.N. CREB-Mediated IL-6 Expression Is Required for 15(S)-Hydroxyeicosatetraenoic Acid-Induced Vascular Smooth Muscle Cell Migration. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Chiappara, G.; Chanez, P.; Bruno, A.; Pace, E.; Pompeo, F.; Bousquet, J.; Bonsignore, G.; Gjomarkaj, M. Variable P-CREB Expression Depicts Different Asthma Phenotypes. Allergy 2007, 62, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.; Ichiki, T.; Fukuyama, K.; Iino, N.; Masuda, S.; Egashira, K.; Takeshita, A. CAMP-Response Element-Binding Protein Mediates Tumor Necrosis Factor-Alpha-Induced Vascular Smooth Muscle Cell Migration. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1634–1639. [Google Scholar] [CrossRef] [Green Version]
- Hondermarck, H.; Bartlett, N.W.; Nurcombe, V. The Role of Growth Factor Receptors in Viral Infections: An Opportunity for Drug Repurposing against Emerging Viral Diseases Such as COVID-19? FASEB BioAdvances 2020, 2, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, Y.; Ichiki, T.; Takeda, K.; Tokuno, T.; Iino, N.; Takeshita, A. Critical Role of CAMP-Response Element-Binding Protein for Angiotensin II-Induced Hypertrophy of Vascular Smooth Muscle Cells. J. Biol. Chem. 2002, 277, 18710–18717. [Google Scholar] [CrossRef] [Green Version]
- Haack, K.K.V.; Mitra, A.K.; Zucker, I.H. NF-ΚB and CREB Are Required for Angiotensin II Type 1 Receptor Upregulation in Neurons. PLoS ONE 2013, 8, e78695. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.; Behm, D.J.; Nerurkar, S.; Eybye, M.E.; Haimbach, R.E.; Olzinski, A.R.; Douglas, S.A.; Willette, R.N. P38 MAPK Inhibitors Ameliorate Target Organ Damage in Hypertension: Part 1. P38 MAPK-Dependent Endothelial Dysfunction and Hypertension. J. Pharmacol. Exp. Ther. 2003, 307, 932–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor CREB in Immune Function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Song, Y.; Xing, R.; Yu, H.; Zhang, Y.; Li, Z.; Gao, W. Heat Shock Protein 70 Acts as a Potential Biomarker for Early Diagnosis of Heart Failure. PLoS ONE 2013, 8, e67964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abaspour, A.R.; Taghikhani, M.; Parizadeh, S.M.R.; Seyedi, S.M.R.; Ghazizadeh, H.; Kazemi, E.; Moohebati, M.; Ghafoori, F.; Mardannik, M.; Avan, A.; et al. HSP27 Expression in the Human Peripheral Blood Mononuclear Cells as an Early Prognostic Biomarker in Coronary Artery Disease Patients. Diabetes Metab. Syndr. 2019, 13, 1791–1795. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Song, D.; Li, H.; He, M.-L. Stress Proteins: The Biological Functions in Virus Infection, Present and Challenges for Target-Based Antiviral Drug Development. Signal. Transduct. Target. Ther. 2020, 5, 125. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Cox, A.J.; Gow, R.M.; Zhang, Y.; Kemp, B.E.; Gilbert, R.E. Platelet-Derived Growth Factor Receptor Transactivation Mediates the Trophic Effects of Angiotensin II in Vivo. Hypertension 2004, 44, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Rajaiya, J.; Yousuf, M.A.; Singh, G.; Stanish, H.; Chodosh, J. Heat Shock Protein 27 Mediated Signaling in Viral Infection. Biochemistry 2012, 51, 5695–5702. [Google Scholar] [CrossRef]
- Alford, K.A.; Glennie, S.; Turrell, B.R.; Rawlinson, L.; Saklatvala, J.; Dean, J.L.E. Heat Shock Protein 27 Functions in Inflammatory Gene Expression and Transforming Growth Factor-β-Activated Kinase-1 (TAK1)-Mediated Signaling. J. Biol. Chem. 2007, 282, 6232–6241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Maat, S.; de Mast, Q.; Danser, A.H.J.; van de Veerdonk, F.L.; Maas, C. Impaired Breakdown of Bradykinin and Its Metabolites as a Possible Cause for Pulmonary Edema in COVID-19 Infection. Semin. Thromb. Hemost. 2020, 46, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Yayama, K.; Hiyoshi, H.; Imazu, D.; Okamoto, H. Angiotensin II Stimulates Endothelial NO Synthase Phosphorylation in Thoracic Aorta of Mice with Abdominal Aortic Banding via Type 2 Receptor. Hypertension 2006, 48, 958–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ping, S.; Chee, R.; Seet, S.; Kennedy, B.K. Does ENOS Derived Nitric Oxide Protect the Young from Severe COVID-19 Complications? Ageing Res. Rev. 2020, 64, 101201. [Google Scholar] [CrossRef]
- Kumar, A.; Nilkanth, S.; Kumar, A.; Singh, D. Glycogen Synthase Kinase-3: A Putative Target to Combat Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Pandemic. Cytokine Growth Factor Rev. 2020, 58, 92–101. [Google Scholar] [CrossRef]
- Caillon, A.; Mian, M.O.R.; Fraulob-Aquino, J.C.; Huo, K.G.; Barhoumi, T.; Ouerd, S.; Sinnaeve, P.R.; Paradis, P.; Schiffrin, E.L. Γδ T Cells Mediate Angiotensin II-Induced Hypertension and Vascular Injury. Circulation 2017, 135, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, T.; Nashabat, M.; Alghanem, B.; Alhallaj, A.; Boudjelal, M.; Umair, M.; Alarifi, S.; Alfares, A.; Mohrij, S.A.A.; Alfadhel, M. Delta Like-1 Gene Mutation: A Novel Cause of Congenital Vertebral Malformation. Front. Genet. 2019, 10, 534. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almutlaq, M.; Mansour, F.A.; Alghamdi, J.; Alhendi, Y.; Alamro, A.A.; Alghamdi, A.A.; Alamri, H.S.; Alroqi, F.; Barhoumi, T. Angiotensin II Exaggerates SARS-CoV-2 Specific T-Cell Response in Convalescent Individuals following COVID-19. Int. J. Mol. Sci. 2022, 23, 8669. https://doi.org/10.3390/ijms23158669
Almutlaq M, Mansour FA, Alghamdi J, Alhendi Y, Alamro AA, Alghamdi AA, Alamri HS, Alroqi F, Barhoumi T. Angiotensin II Exaggerates SARS-CoV-2 Specific T-Cell Response in Convalescent Individuals following COVID-19. International Journal of Molecular Sciences. 2022; 23(15):8669. https://doi.org/10.3390/ijms23158669
Chicago/Turabian StyleAlmutlaq, Moudhi, Fatmah A. Mansour, Jahad Alghamdi, Yassen Alhendi, Abir Abdullah Alamro, Amani Ahmed Alghamdi, Hassan S. Alamri, Fayhan Alroqi, and Tlili Barhoumi. 2022. "Angiotensin II Exaggerates SARS-CoV-2 Specific T-Cell Response in Convalescent Individuals following COVID-19" International Journal of Molecular Sciences 23, no. 15: 8669. https://doi.org/10.3390/ijms23158669