
Methyl Group Metabolism in Differentiation, Aging, and Cancer


Abstract
:1. Introduction
2. Methyl Group Metabolism
2.1. The One-Carbon Metabolism
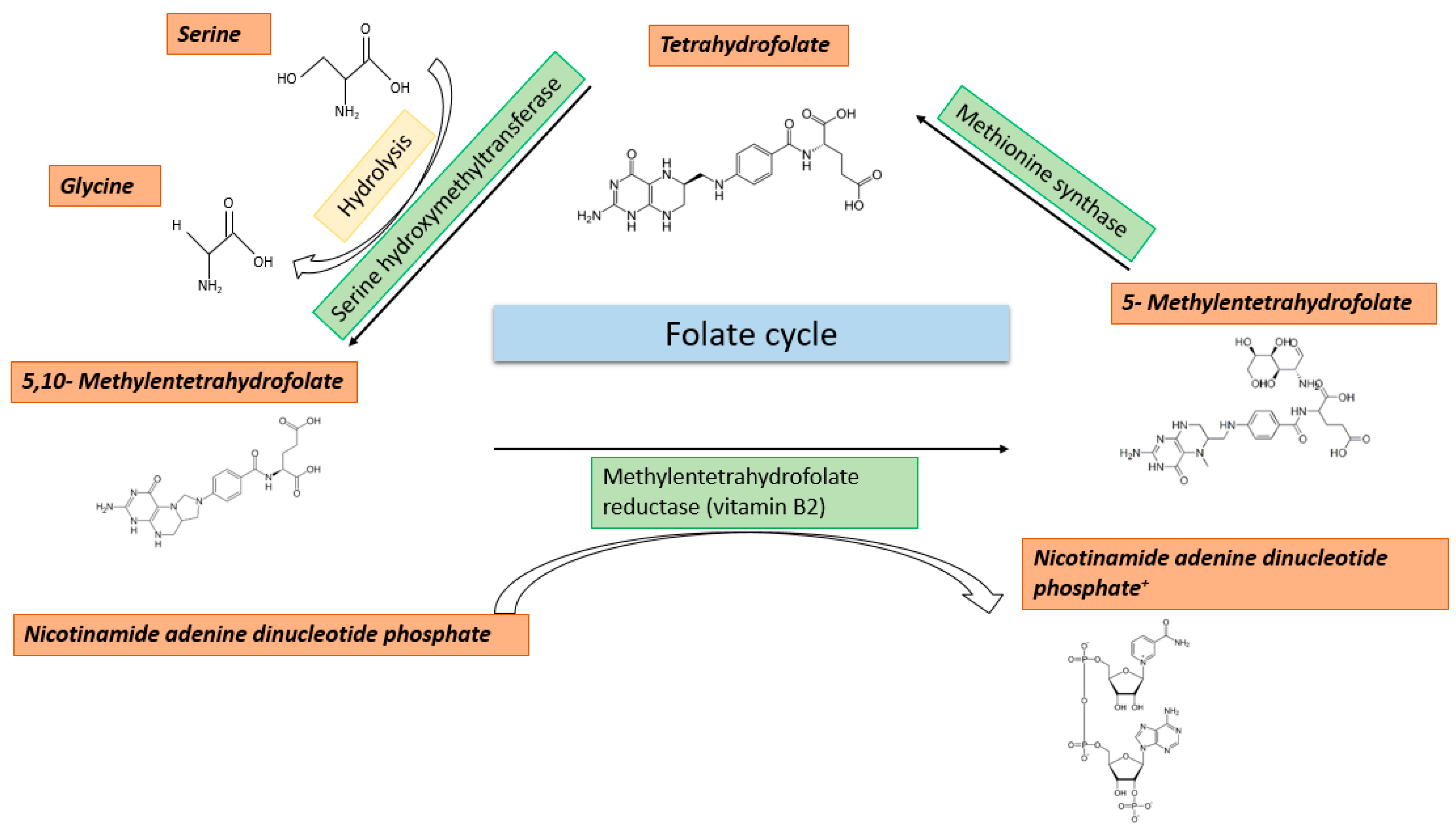
2.2. Folate Cycle
2.3. Methionine Cycle
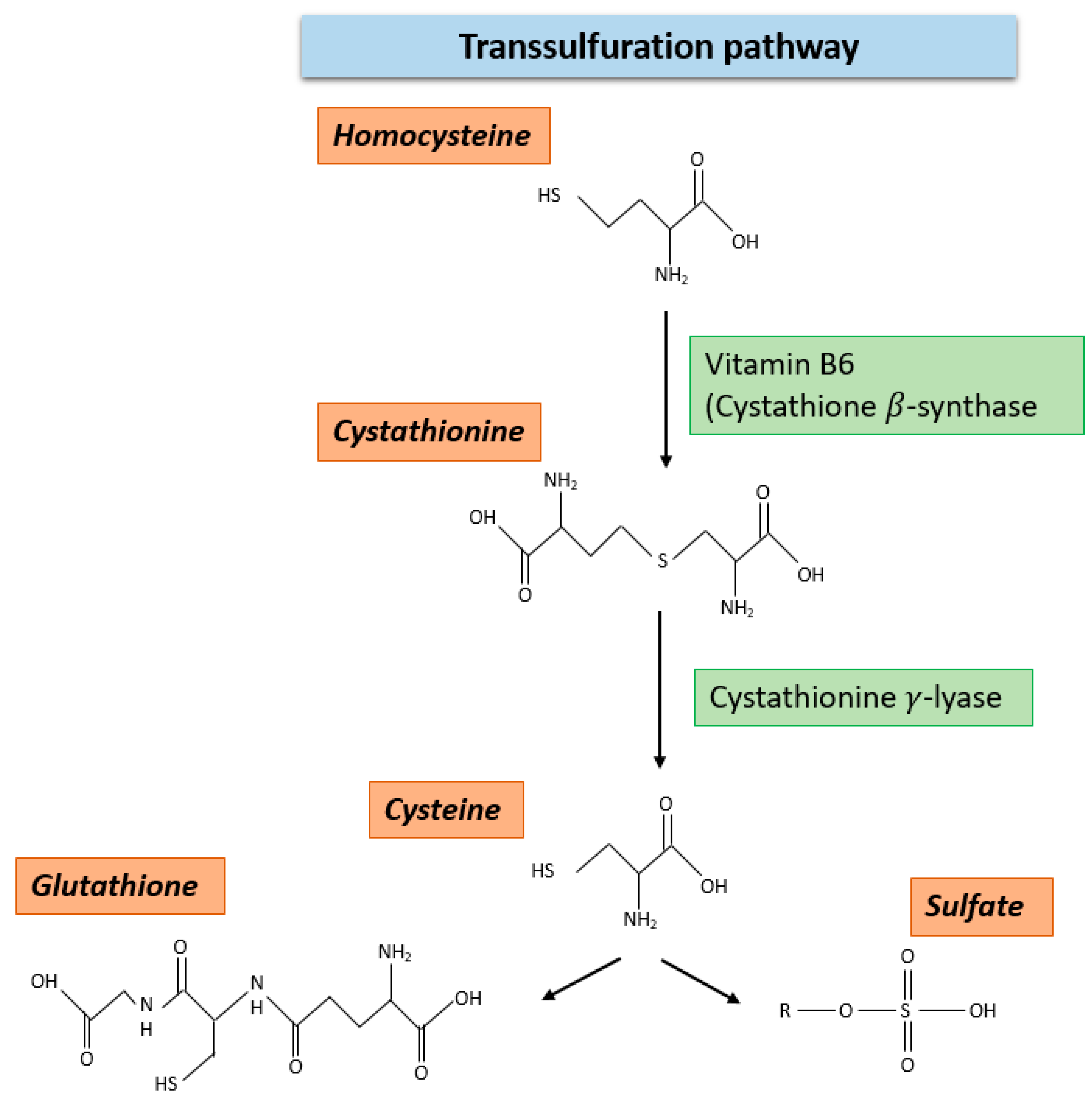
2.4. Transsulfuration Pathway
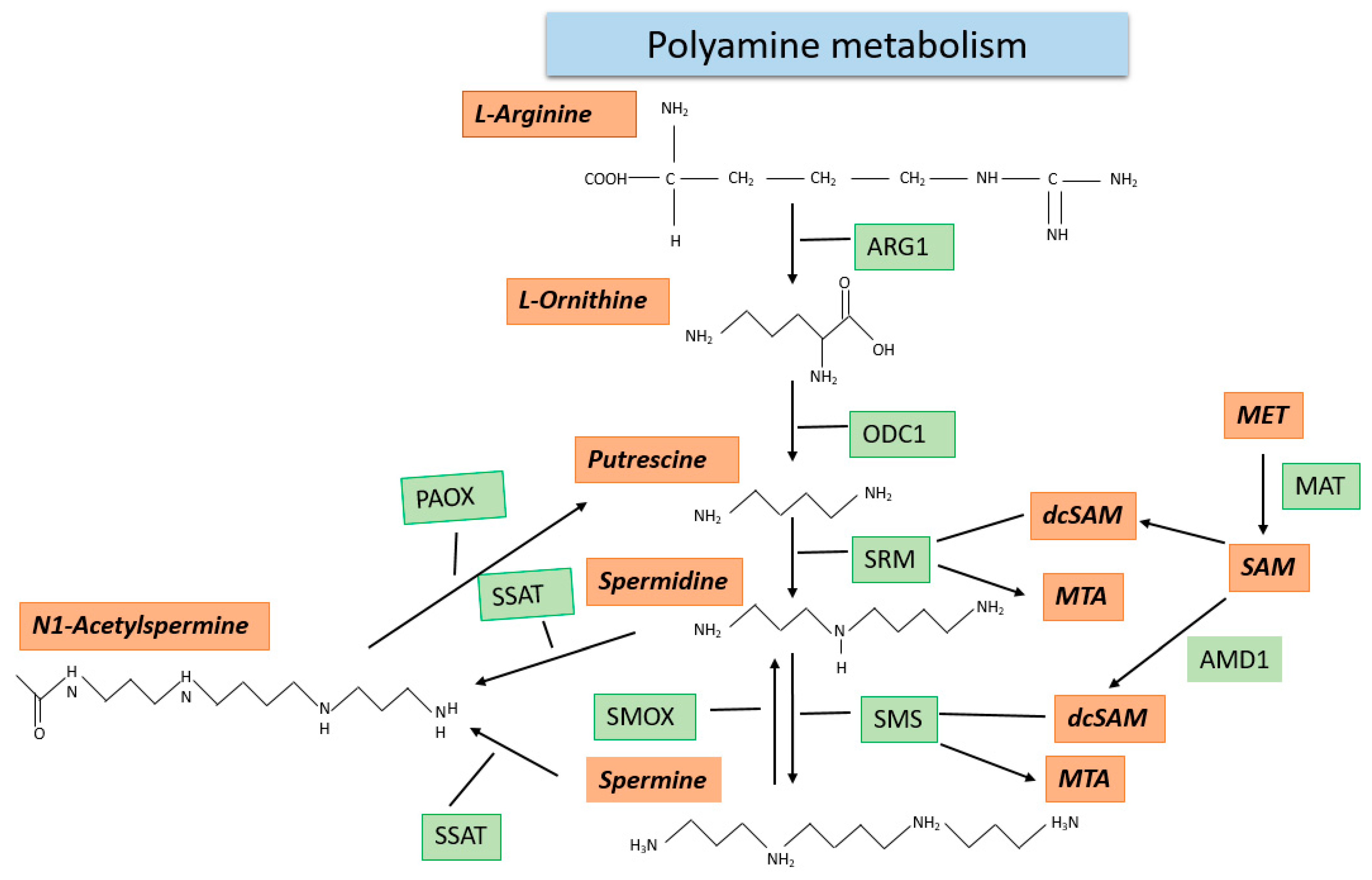
2.5. Polyamine Metabolism
3. Influence of the One-Carbon Metabolism and the Polyamine Metabolism on Epigenetics and Cancer
3.1. Metabolic Influence on Epigenetics
3.2. Metabolic Influence on Cancer Development and Progression
4. Nutrition and Plant-Extracted Compounds Influencing the One-Carbon- and Polyamine Metabolism
4.1. Nutrition
4.2. Naturally Existing Plant-Extracted Compounds with Impact on DNA Methylation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hemberger, M.; Dean, W.; Reik, W. Epigenetic dynamics of stem cells and cell lineage commitment: Digging Waddington’s canal. Nat. Rev. Mol. Cell Biol. 2009, 10, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yang, M.; Li, S.; Zhang, J.; Peng, B.; Wang, C.; Chang, Z.; Ong, J.; Du, P. Mouse totipotent stem cells captured and maintained through spliceosomal repression. Cell 2021, 184, 2843–2859. [Google Scholar] [CrossRef] [PubMed]
- Venter, J.C.; Adams, M.D.; Myers, E.W.; Li, P.W.; Mural, R.J.; Sutton, G.G.; Smith, H.O.; Yandell, M.; Evans, C.A.; Holt, R.A.; et al. The sequence of the human genome. Science 2001, 291, 1304–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.H. The epigenotype. Endeavour 1942, 1, 18–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, M.; Boubelik, M.; Lehnert, S. Temporal and regional changes in DNA methylation in the embryonic, extraembryonic and germ cell lineages during mouse embryo development. Development 1987, 99, 371–382. [Google Scholar] [CrossRef]
- Kafri, T.; Ariel, M.; Brandeis, M.; Shemer, R.; Urven, L.; McCarrey, J.; Cedar, H.; Razin, A. Developmental pattern of gene-specific DNA methylation in the mouse embryo and germ line. Genes Dev. 1992, 6, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Cedar, H.; Bergman, Y. Programming of DNA methylation patterns. Annu. Rev. Biochem. 2012, 81, 97–117. [Google Scholar] [CrossRef] [Green Version]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Ryall, J.G.; Cliff, T.; Dalton, S.; Sartorelli, V. Metabolic Reprogramming of Stem Cell Epigenetics. Cell Stem Cell 2015, 17, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Vousden, K.H. Serine and one-carbon metabolism in cancer. Nat. Rev. Cancer 2016, 16, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Shyh-Chang, N.; Locasale, J.W.; Lyssiotis, C.A.; Zheng, Y.; Teo, R.Y.; Ratanasirintrawoot, S.; Zhang, J.; Onder, T.; Unternaehrer, J.J.; Zhu, H.; et al. Influence of threonine metabolism on S-adenosylmethionine and histone methylation. Science 2013, 339, 222–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.C.; Shyh-Chang, N.; Yang, H.; Rai, A.; Umashankar, S.; Ma, S.; Soh, B.S.; Sun, L.L.; Tai, B.C.; Nga, M.E.; et al. Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis. Cell 2012, 148, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, M.E.; MacMillan, L.; Stevens, J.R.; Brosnan, J.T. Division of labour: How does folate metabolism partition between one-carbon metabolism and amino acid oxidation? Biochem. J. 2015, 472, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducker, G.S.; Chen, L.; Morscher, R.J.; Ghergurovich, J.M.; Esposito, M.; Teng, X.; Kang, Y.; Rabinowitz, J.D. Reversal of Cytosolic One-Carbon Flux Compensates for Loss of the Mitochondrial Folate Pathway. Cell Metab. 2016, 23, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Sunden, S.L.; Renduchintala, M.S.; Park, E.I.; Miklasz, S.D.; Garrow, T.A. Betaine-homocysteine methyltransferase expression in porcine and human tissues and chromosomal localization of the human gene. Arch. Biochem. Biophys. 1997, 345, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Poole, J.R. Labile methyl balances for normal humans on various dietary regimens. Metab. Clin. Exp. 1975, 24, 721–735. [Google Scholar] [CrossRef]
- Storch, K.J.; Wagner, D.A.; Burke, J.F.; Young, V.R. Quantitative study in vivo of methionine cycle in humans using methyl-2H3- and 1-13Cmethionine. Am. J. Physiol. 1988, 255, E322–E331. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Igarashi, K. Polyamines and their metabolites as diagnostic markers of human diseases. Biomol. Ther. 2013, 21, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, J.M.; Pegg, A.E.; McCloskey, D.E. Drugs affecting the cell cycle via actions on the polyamine metabolic pathway. Prog. Cell Cycle Res. 2003, 5, 461–468. [Google Scholar] [PubMed]
- Seiler, N.; Raul, F. Polyamines and apoptosis. J. Cell. Mol. Med. 2005, 9, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Tabor, C.W.; Tabor, H. Polyamines. Annu. Rev. Biochem. 1984, 53, 749–790. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, B.; Karlic, H.; Varga, F.; Fabianowska-Majewska, K.; Haslberger, A. Epigenetic mechanisms in anti-cancer actions of bioactive food components--the implications in cancer prevention. Br. J. Pharmacol. 2012, 167, 279–297. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, B.N.; Arnold, K.M.; Denu, J.M. KAT(ching) metabolism by the tail: Insight into the links between lysine acetyltransferases and metabolism. Chembiochem 2011, 12, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, J.L. Metabolic mechanisms of epigenetic regulation. ACS Chem. Biol. 2013, 8, 2607–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.V.; Carrer, A.; Shah, S.; Snyder, N.W.; Wei, S.; Venneti, S.; Worth, A.J.; Yuan, Z.-F.; Lim, H.-W.; Liu, S.; et al. Akt-dependent metabolic reprogramming regulates tumor cell histone acetylation. Cell Metab. 2014, 20, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruti, S.S.; Ulrich, C.M.; White, E. Folate and one-carbon metabolism nutrients from supplements and diet in relation to breast cancer risk. Am. J. Clin. Nutr. 2009, 89, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Akoglu, B.; Milovic, V.; Caspary, W.F.; Faust, D. Hyperproliferation of homocysteine-treated colon cancer cells is reversed by folate and 5-methyltetrahydrofolate. Eur. J. Nutr. 2004, 43, 93–99. [Google Scholar] [CrossRef]
- Pegg, A.E.; Michael, A.J. Spermine synthase. Cell. Mol. Life Sci. 2010, 67, 113–121. [Google Scholar] [CrossRef]
- Chen, Z.; Karaplis, A.C.; Ackerman, S.L.; Pogribny, I.P.; Melnyk, S.; Lussier-Cacan, S.; Chen, M.F.; Pai, A.; John, S.W.; Smith, R.S.; et al. Mice deficient in methylenetetrahydrofolate reductase exhibit hyperhomocysteinemia and decreased methylation capacity, with neuropathology and aortic lipid deposition. Hum. Mol. Genet. 2001, 10, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, L.A.; Wise, C.K.; Delongchamp, R.R.; Sinha, R. Blood determinations of S-adenosylmethionine, S-adenosylhomocysteine, and homocysteine: Correlations with diet. Cancer Epidemiol. Biomark. Prev. 2001, 10, 649–655. [Google Scholar]
- Lim, U.; Song, M.-A. Dietary and lifestyle factors of DNA methylation. Methods Mol. Biol. 2012, 863, 359–376. [Google Scholar] [CrossRef]
- Mentch, S.J.; Mehrmohamadi, M.; Huang, L.; Liu, X.; Gupta, D.; Mattocks, D.; Gómez Padilla, P.; Ables, G.; Bamman, M.M.; Thalacker-Mercer, A.E.; et al. Histone Methylation Dynamics and Gene Regulation Occur through the Sensing of One-Carbon Metabolism. Cell Metab. 2015, 22, 861–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, K.N.; Hodges, R.; Hanes, D.; Stack, E.; Cheishvili, D.; Szyf, M.; Henkel, J.; Twedt, M.W.; Giannopoulou, D.; Herdell, J.; et al. Potential reversal of epigenetic age using a diet and lifestyle intervention: A pilot randomized clinical trial. Aging 2021, 13, 9419–9432. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Kinnaird, A.; Zhao, S.; Wellen, K.E.; Michelakis, E.D. Metabolic control of epigenetics in cancer. Nat. Rev. Cancer 2016, 16, 694–707. [Google Scholar] [CrossRef]
- Erichsen, L.; Beermann, A.; Arauzo-Bravo, M.J.; Hassan, M.; Dkhil, M.A.; Al-Quraishy, S.; Hafiz, T.A.; Fischer, J.C.; Santourlidis, S. Genome-wide hypomethylation of LINE-1 and Alu retroelements in cell-free DNA of blood is an epigenetic biomarker of human aging. Saudi J. Biol. Sci. 2018, 25, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, Y.-I.; Fang, J.; Erdjument-Bromage, H.; Warren, M.E.; Borchers, C.H.; Tempst, P.; Zhang, Y. Histone demethylation by a family of JmjC domain-containing proteins. Nature 2006, 439, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Yang, H.; Xu, W.; Ma, S.; Lin, H.; Zhu, H.; Liu, L.; Liu, Y.; Yang, C.; Xu, Y.; et al. Inhibition of α-KG-dependent histone and DNA demethylases by fumarate and succinate that are accumulated in mutations of FH and SDH tumor suppressors. Genes Dev. 2012, 26, 1326–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killian, J.K.; Kim, S.Y.; Miettinen, M.; Smith, C.; Merino, M.; Tsokos, M.; Quezado, M.; Smith, W.I.; Jahromi, M.S.; Xekouki, P.; et al. Succinate dehydrogenase mutation underlies global epigenomic divergence in gastrointestinal stromal tumor. Cancer Discov. 2013, 3, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, Y.; Ikura, T.; Hoshikawa, Y.; Tashiro, S.; Ito, T.; Ohta, M.; Kera, Y.; Noda, T.; Igarashi, K. Methionine adenosyltransferase II serves as a transcriptional corepressor of Maf oncoprotein. Mol. Cell 2011, 41, 554–566. [Google Scholar] [CrossRef]
- Li, S.; Swanson, S.K.; Gogol, M.; Florens, L.; Washburn, M.P.; Workman, J.L.; Suganuma, T. Serine and SAM Responsive Complex SESAME Regulates Histone Modification Crosstalk by Sensing Cellular Metabolism. Mol. Cell 2015, 60, 408–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Qian, X.; Shen, J.; Wang, Y.; Li, X.; Liu, R.; Xia, Y.; Chen, Q.; Peng, G.; Lin, S.-Y.; et al. Local generation of fumarate promotes DNA repair through inhibition of histone H3 demethylation. Nat. Cell Biol. 2015, 17, 1158–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. Über den Stoffwechsel der Carcinomzelle. Naturwissenschaften 1924, 12, 1131–1137. [Google Scholar] [CrossRef]
- Lu, J.; Tan, M.; Cai, Q. The Warburg effect in tumor progression: Mitochondrial oxidative metabolism as an anti-metastasis mechanism. Cancer Lett. 2015, 356, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farber, S.; Diamond, L.K. Temporary remissions in acute leukemia in children produced by folic acid antagonist, 4-aminopteroyl-glutamic acid. N. Engl. J. Med. 1948, 238, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.C.; Maddocks, O.D.K. One-carbon metabolism in cancer. Br. J. Cancer 2017, 116, 1499–1504. [Google Scholar] [CrossRef] [Green Version]
- Gonen, N.; Assaraf, Y.G. Antifolates in cancer therapy: Structure, activity and mechanisms of drug resistance. Drug Resist. Updat. 2012, 15, 183–210. [Google Scholar] [CrossRef]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Goulian, M.; Bleile, B.; Tseng, B.Y. Methotrexate-induced misincorporation of uracil into DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 1956–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blount, B.C.; Mack, M.M.; Wehr, C.M.; MacGregor, J.T.; Hiatt, R.A.; Wang, G.; Wickramasinghe, S.N.; Everson, R.B.; Ames, B.N. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: Implications for cancer and neuronal damage. Proc. Natl. Acad. Sci. USA 1997, 94, 3290–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schernhammer, E.S.; Giovannucci, E.; Kawasaki, T.; Rosner, B.; Fuchs, C.S.; Ogino, S. Dietary folate, alcohol and B vitamins in relation to LINE-1 hypomethylation in colon cancer. Gut 2010, 59, 794–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, R.A.; Mak, T.W. Oncogenic isocitrate dehydrogenase mutations: Mechanisms, models, and clinical opportunities. Cancer Discov. 2013, 3, 730–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, M.E.; Abdel-Wahab, O.; Lu, C.; Ward, P.S.; Patel, J.; Shih, A.; Li, Y.; Bhagwat, N.; Vasanthakumar, A.; Fernandez, H.F.; et al. Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell 2010, 18, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Losman, J.-A.; Looper, R.E.; Koivunen, P.; Lee, S.; Schneider, R.K.; McMahon, C.; Cowley, G.S.; Root, D.E.; Ebert, B.L.; Kaelin, W.G. (R)-2-hydroxyglutarate is sufficient to promote leukemogenesis and its effects are reversible. Science 2013, 339, 1621–1625. [Google Scholar] [CrossRef]
- Yousef, A.-M.; Shomaf, M.; Berger, S.; Ababneh, N.; Bobali, Y.; Ali, D.; Al-Hasan, S.; Diab, O.; Ismail, S. Allele and genotype frequencies of the polymorphic methylenetetrahydrofolate reductase and colorectal cancer among Jordanian population. Asian Pac. J. Cancer Prev. 2013, 14, 4559–4565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senses, K.M.; Gonen, M.; Barutcu, A.R.; Kalaylioglu, Z.; Isbilen, M.; Konu, O.; Chen, Y.T.; Altorki, N.K.; Gure, A.O. Cancer-testis gene expression is associated with the methylenetetrahydrofolate reductase 677 CT polymorphism in non-small cell lung carcinoma. BMC Med. Genet. 2013, 14, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Xu, D.; Wang, P.; Wang, P.; Liu, C.; Hua, C.; Jiang, T.; Zhang, B.; Li, Z.; Lu, L.; et al. MTHFR C677T polymorphism contributes to the risk for gastric cancer. Tumour Biol. 2014, 35, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Ulrey, C.L.; Liu, L.; Andrews, L.G.; Tollefsbol, T.O. The impact of metabolism on DNA methylation. Hum. Mol. Genet. 2005, 14 (Suppl. S1), R139–R147. [Google Scholar] [CrossRef]
- Llanos, A.A.M.; Marian, C.; Brasky, T.M.; Dumitrescu, R.G.; Liu, Z.; Mason, J.B.; Makambi, K.H.; Spear, S.L.; Kallakury, B.V.S.; Freudenheim, J.L.; et al. Associations between genetic variation in one-carbon metabolism and LINE-1 DNA methylation in histologically normal breast tissues. Epigenetics 2015, 10, 727–735. [Google Scholar] [CrossRef]
- Ouerhani, S.; Oliveira, E.; Marrakchi, R.; Ben Slama, M.R.; Sfaxi, M.; Ayed, M.; Chebil, M.; Amorim, A.; El Gaaied, A.B.; Prata, M.J. Methylenetetrahydrofolate reductase and methionine synthase polymorphisms and risk of bladder cancer in a Tunisian population. Cancer Genet. Cytogenet. 2007, 176, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Erichsen, L.; Ghanjati, F.; Beermann, A.; Poyet, C.; Hermanns, T.; Schulz, W.A.; Seifert, H.-H.; Wild, P.J.; Buser, L.; Kröning, A.; et al. Aberrant methylated key genes of methyl group metabolism within the molecular etiology of urothelial carcinogenesis. Sci. Rep. 2018, 8, 3477. [Google Scholar] [CrossRef] [PubMed]
- Vaissière, T.; Hung, R.J.; Zaridze, D.; Moukeria, A.; Cuenin, C.; Fasolo, V.; Ferro, G.; Paliwal, A.; Hainaut, P.; Brennan, P.; et al. Quantitative analysis of DNA methylation profiles in lung cancer identifies aberrant DNA methylation of specific genes and its association with gender and cancer risk factors. Cancer Res. 2009, 69, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medici, V.; Shibata, N.M.; Kharbanda, K.K.; LaSalle, J.M.; Woods, R.; Liu, S.; Engelberg, J.A.; Devaraj, S.; Török, N.J.; Jiang, J.X.; et al. Wilson’s disease: Changes in methionine metabolism and inflammation affect global DNA methylation in early liver disease. Hepatology 2013, 57, 555–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, J.F.; Ferrer, I.; Blanco-Aparicio, C.; Hernández-Losa, J.; Ramón, Y.; Cajal, S.; Carnero, A.; Lleonart, M.E. S-adenosylhomocysteine hydrolase downregulation contributes to tumorigenesis. Carcinogenesis 2008, 29, 2089–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belužić, L.; Grbeša, I.; Belužić, R.; Park, J.H.; Kong, H.K.; Kopjar, N.; Espadas, G.; Sabidó, E.; Lepur, A.; Rokić, F.; et al. Knock-down of AHCY and depletion of adenosine induces DNA damage and cell cycle arrest. Sci. Rep. 2018, 8, 14012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heljasvaara, R.; Veress, I.; Halmekytö, M.; Alhonen, L.; Jänne, J.; Laajala, P.; Pajunen, A. Transgenic mice overexpressing ornithine and S-adenosylmethionine decarboxylases maintain a physiological polyamine homoeostasis in their tissues. Biochem. J. 1997, 323 Pt 2, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiou, G.A.; Lianos, G.D.; Ragos, V.; Galani, V.; Kyritsis, A.P. Difluoromethylornithine in cancer: New advances. Future Oncol. 2017, 13, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Erichsen, L.; Seifert, H.-H.; Schulz, W.A.; Hoffmann, M.J.; Niegisch, G.; Araúzo-Bravo, M.J.; Bendhack, M.L.; Poyet, C.; Hermanns, T.; Beermann, A.; et al. Basic Hallmarks of Urothelial Cancer Unleashed in Primary Uroepithelium by Interference with the Epigenetic Master Regulator ODC1. Sci. Rep. 2020, 10, 3808. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, D.; Shima, K.; Matsuo, K.; Nishioka, T.; Chen, C.Y.; Hu, G.-F.; Sasaki, A.; Tsuji, T. Ornithine decarboxylase antizyme induces hypomethylation of genome DNA and histone H3 lysine 9 dimethylation (H3K9me2) in human oral cancer cell line. PLoS ONE 2010, 5, e12554. [Google Scholar] [CrossRef] [Green Version]
- Florl, A.R.; Löwer, R.; Schmitz-Dräger, B.J.; Schulz, W.A. DNA methylation and expression of LINE-1 and HERV-K provirus sequences in urothelial and renal cell carcinomas. Br. J. Cancer 1999, 80, 1312–1321. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, M. DNA hypomethylation in cancer cells. Epigenomics 2009, 1, 239–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quach, A.; Levine, M.E.; Tanaka, T.; Lu, A.T.; Chen, B.H.; Ferrucci, L.; Ritz, B.; Bandinelli, S.; Neuhouser, M.L.; Beasley, J.M.; et al. Epigenetic clock analysis of diet, exercise, education, and lifestyle factors. Aging 2017, 9, 419–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sybesma, W.; Starrenburg, M.; Tijsseling, L.; Hoefnagel, M.H.N.; Hugenholtz, J. Effects of cultivation conditions on folate production by lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 4542–4548. [Google Scholar] [CrossRef] [Green Version]
- Hariri, M.; Salehi, R.; Feizi, A.; Mirlohi, M.; Ghiasvand, R.; Habibi, N. A randomized, double-blind, placebo-controlled, clinical trial on probiotic soy milk and soy milk: Effects on epigenetics and oxidative stress in patients with type II diabetes. Genes Nutr. 2015, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (U.S.). Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B₆, Folate, Vitamin B₁₂, Pantothenic Acid, Biotin, and Choline; National Academy Press: Washington, DC, USA, 1998; ISBN 9780309065542. [Google Scholar]
- Shao, A.; Hathcock, J.N. Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regul. Toxicol. Pharmacol. 2008, 50, 376–399. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of betaine as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, e05658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momose, Y.; Maeda-Yamamoto, M.; Nabetani, H. Systematic review of green tea epigallocatechin gallate in reducing low-density lipoprotein cholesterol levels of humans. Int. J. Food Sci. Nutr. 2016, 67, 606–613. [Google Scholar] [CrossRef]
- Formica, J.V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academy Press: Washington, DC, USA, 2005; ISBN 978-0-309-08525-0. [Google Scholar]
- Christensen, K.E.; Wu, Q.; Wang, X.; Deng, L.; Caudill, M.A.; Rozen, R. Steatosis in mice is associated with gender, folate intake, and expression of genes of one-carbon metabolism. J. Nutr. 2010, 140, 1736–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolders, R.G.V.; de Meer, K.; Kenemans, P.; Jakobs, C.; Kulik, W.; van der Mooren, M.J. Oral estradiol decreases plasma homocysteine, vitamin B6, and albumin in postmenopausal women but does not change the whole-body homocysteine remethylation and transmethylation flux. J. Clin. Endocrinol. Metab. 2005, 90, 2218–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, S.H.; Mar, M.H.; Zhou, Z.; da Costa, K.A. Pregnancy and lactation are associated with diminished concentrations of choline and its metabolites in rat liver. J. Nutr. 1995, 125, 3049–3054. [Google Scholar] [CrossRef] [PubMed]
- Sadre-Marandi, F.; Dahdoul, T.; Reed, M.C.; Nijhout, H.F. Sex differences in hepatic one-carbon metabolism. BMC Syst. Biol. 2018, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, J.D.; Kyle, W.E.; Harris, B.J. Methionine metabolism in mammals. Regulation of homocysteine methyltransferases in rat tissue. Arch. Biochem. Biophys. 1971, 146, 84–92. [Google Scholar] [CrossRef]
- Schwahn, B.C.; Chen, Z.; Laryea, M.D.; Wendel, U.; Lussier-Cacan, S.; Genest, J.; Mar, M.-H.; Zeisel, S.H.; Castro, C.; Garrow, T.; et al. Homocysteine-betaine interactions in a murine model of 5,10-methylenetetrahydrofolate reductase deficiency. FASEB J. 2003, 17, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Välimäki, M.J.; Härkönen, M.; Peter Eriksson, C.J.; Ylikahri, R.H. Sex hormones and adrenocortical steroids in men acutely intoxicated with ethanol. Alcohol 1984, 1, 89–93. [Google Scholar] [CrossRef]
- Frias, J.; Torres, J.M.; Miranda, M.T.; Ruiz, E.; Ortega, E. Effects of acute alcohol intoxication on pituitary-gonadal axis hormones, pituitary-adrenal axis hormones, beta-endorphin and prolactin in human adults of both sexes. Alcohol Alcohol. 2002, 37, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Sierksma, A.; Sarkola, T.; Eriksson, C.J.P.; van der Gaag, M.S.; Grobbee, D.E.; Hendriks, H.F.J. Effect of moderate alcohol consumption on plasma dehydroepiandrosterone sulfate, testosterone, and estradiol levels in middle-aged men and postmenopausal women: A diet-controlled intervention study. Alcohol. Clin. Exp. Res. 2004, 28, 780–785. [Google Scholar] [CrossRef] [Green Version]
- Hämäläinen, E.; Adlercreutz, H.; Puska, P.; Pietinen, P. Diet and serum sex hormones in healthy men. J. Steroid Biochem. 1984, 20, 459–464. [Google Scholar] [CrossRef]
- Reed, M.J.; Cheng, R.W.; Simmonds, M.; Richmond, W.; James, V.H. Dietary lipids: An additional regulator of plasma levels of sex hormone binding globulin. J. Clin. Endocrinol. Metab. 1987, 64, 1083–1085. [Google Scholar] [CrossRef]
- Goldin, B.R.; Woods, M.N.; Spiegelman, D.L.; Longcope, C.; Morrill-LaBrode, A.; Dwyer, J.T.; Gualtieri, L.J.; Hertzmark, E.; Gorbach, S.L. The effect of dietary fat and fiber on serum estrogen concentrations in premenopausal women under controlled dietary conditions. Cancer 1994, 74, 1125–1131. [Google Scholar] [CrossRef]
- Hill, P.B.; Wynder, E.L. Effect of a vegetarian diet and dexamethasone on plasma prolactin, testosterone and dehydroepiandrosterone in men and women. Cancer Lett. 1979, 7, 273–282. [Google Scholar] [CrossRef]
- Howie, B.J.; Shultz, T.D. Dietary and hormonal interrelationships among vegetarian Seventh-Day Adventists and nonvegetarian men. Am. J. Clin. Nutr. 1985, 42, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélanger, A.; Locong, A.; Noel, C.; Cusan, L.; Dupont, A.; Prévost, J.; Caron, S.; Sévigny, J. Influence of diet on plasma steroid and sex plasma binding globulin levels in adult men. J. Steroid Biochem. 1989, 32, 829–833. [Google Scholar] [CrossRef]
- Key, T.J.; Roe, L.; Thorogood, M.; Moore, J.W.; Clark, G.M.; Wang, D.Y. Testosterone, sex hormone-binding globulin, calculated free testosterone, and oestradiol in male vegans and omnivores. Br. J. Nutr. 1990, 64, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, L.W.; Cureton, K.J.; Thompson, F.N. Comparison of serum testosterone and androstenedione responses to weight lifting in men and women. Europ. J. Appl. Physiol. 1983, 50, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, K.; Pakarinen, A. Acute hormonal responses to two different fatiguing heavy-resistance protocols in male athletes. J. Appl. Physiol. 1993, 74, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Chandler, R.M.; Byrne, H.K.; Patterson, J.G.; Ivy, J.L. Dietary supplements affect the anabolic hormones after weight-training exercise. J. Appl. Physiol. 1994, 76, 839–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jotova, I.; Wang, C.; Tabib, A.; Dimitrov, O.; Bachrach, U. Effects of testosterone and 17, beta-estradiol on the polyamine metabolism in cultivated normal rat kidney epithelial cells. Amino Acids 2000, 18, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Levillain, O.; Diaz, J.-J.; Blanchard, O.; Déchaud, H. Testosterone down-regulates ornithine aminotransferase gene and up-regulates arginase II and ornithine decarboxylase genes for polyamines synthesis in the murine kidney. Endocrinology 2005, 146, 950–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, L.K.; Friso, S.; Choi, S.-W. Nutritional influences on epigenetics and age-related disease. Proc. Nutr. Soc. 2012, 71, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Ghazi, T.; Arumugam, T.; Foolchand, A.; Chuturgoon, A.A. The Impact of Natural Dietary Compounds and Food-Borne Mycotoxins on DNA Methylation and Cancer. Cells 2020, 9, 2004. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Park, B.; Goel, A.; Aggarwal, B.B. Epigenetic changes induced by curcumin and other natural compounds. Genes Nutr. 2011, 6, 93–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xie, Z.; Jones, W.; Pavlovicz, R.E.; Liu, S.; Yu, J.; Li, P.-K.; Lin, J.; Fuchs, J.R.; Marcucci, G.; et al. Curcumin is a potent DNA hypomethylation agent. Bioorg. Med. Chem. Lett. 2009, 19, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Wargovich, M.J. Experimental evidence for cancer preventive elements in foods. Cancer Lett. 1997, 114, 11–17. [Google Scholar] [CrossRef]
- Paluszczak, J.; Krajka-Kuźniak, V.; Baer-Dubowska, W. The effect of dietary polyphenols on the epigenetic regulation of gene expression in MCF7 breast cancer cells. Toxicol. Lett. 2010, 192, 119–125. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M.; Mazzone, M.G.; Giuliano, F.; Basile, G.; Agodi, A. Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation. Int. J. Mol. Sci. 2018, 19, 2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tili, E.; Michaille, J.-J.; Alder, H.; Volinia, S.; Delmas, D.; Latruffe, N.; Croce, C.M. Resveratrol modulates the levels of microRNAs targeting genes encoding tumor-suppressors and effectors of TGFβ signaling pathway in SW480 cells. Biochem. Pharmacol. 2010, 80, 2057–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majid, S.; Dar, A.A.; Ahmad, A.E.; Hirata, H.; Kawakami, K.; Shahryari, V.; Saini, S.; Tanaka, Y.; Dahiya, A.V.; Khatri, G.; et al. BTG3 tumor suppressor gene promoter demethylation, histone modification and cell cycle arrest by genistein in renal cancer. Carcinogenesis 2009, 30, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majid, S.; Dar, A.A.; Shahryari, V.; Hirata, H.; Ahmad, A.; Saini, S.; Tanaka, Y.; Dahiya, A.V.; Dahiya, R. Genistein reverses hypermethylation and induces active histone modifications in tumor suppressor gene B-Cell translocation gene 3 in prostate cancer. Cancer 2010, 116, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Food | µg/100 g | Recommended Dietary Intake |
|---|---|---|---|
| Folate | Chicken liver | 578 | 330 μg/day [77] |
| Calf liver | 331 | ||
| Peanuts | 246 | ||
| Sunflower seed kernels | 238 | ||
| Lentils | 181 | ||
| Chickpeas | 172 | ||
| Asparagus | 149 | ||
| Spinach | 146 | ||
| Lettuce | 136 | ||
| Peanuts (oil roasted) | 125 | ||
| Soybeans | 111 | ||
| Broccoli | 108 | ||
| Walnuts | 98 | ||
| Peanut butter | 92 | ||
| Hazelnuts | 88 | ||
| Avocados | 81 | ||
| Beets | 80 | ||
| Kale | 65 | ||
| Bread | 65 | ||
| Cheese | 20–60 | ||
| Cabbage | 46 | ||
| Red bell peppers | 46 | ||
| Cauliflower | 44 | ||
| Chicken eggs | 44 | ||
| Salmon | 35 | ||
| Tofu | 29 | ||
| Potatoes | 28 | ||
| Chicken | 12 | ||
| Beef | 12 | ||
| Yoghurt | 8–11 | ||
| Pork | 8 | ||
| Milk | 5 | ||
| Butter | 3 | ||
| mg/100 g | |||
| Arginine | Pumpkin seeds | 5353 | 20 grams per day [78] |
| Peanuts (roasted) | 2832 | ||
| Pine nuts | 2413 | ||
| Walnuts | 2278 | ||
| Peas (dried) | 2278 | ||
| Chicken breast (raw) | 1436 | ||
| Pork (raw) | 1394 | ||
| Salmon (raw) | 1221 | ||
| Buckwheat grains | 982 | ||
| Egg | 820 | ||
| Wheat flour | 642 | ||
| Rice | 602 | ||
| Corn flour | 345 | ||
| Milk | 119 | ||
| Betaine | Quinoa | 630 | 6 mg/kg body weight per day in addition to the intake from the background diet [79] |
| Wheat germ | 410 | ||
| Lamb’s quarters | 330 | ||
| Wheat bran | 320 | ||
| Canned beetroot | 260 | ||
| Dark rye flour | 150 | ||
| Spinach | 110–130 | ||
| Red wine | 0.76 | ||
| Fish (shrimp) | 0.75 | ||
| Fish (tuna) | 0.75 | ||
| Fish (salmon) | 0.35 | ||
| White wine | 0.12 | ||
| Grapes | 0.11 | ||
| Epigallocatechin gallate | Green tea (brewed) | 70 | 107 to 856 mg/day [80] |
| White tea (brewed) | 42.45 | ||
| Black tea (brewed) | 9.36 | ||
| Green tea | 3.96 | ||
| Pecans | 2.3 | ||
| Hazelnut | 1.06 | ||
| Cranberries | 0.97 | ||
| Blackberries | 0.68 | ||
| Raspberries | 0.54 | ||
| Black tea | 0.51 | ||
| Pistachios | 0.4 | ||
| Plums | 0.4 | ||
| Peaches | 0.3 | ||
| Apples | 0.24 | ||
| Glutamic acid | Wheat flour | 4328 | |
| Peas (dried) | 4196 | ||
| Chicken breat (raw) | 3458 | ||
| Beef (raw) | 3191 | ||
| Salmon (raw) | 2830 | ||
| Walnuts | 2816 | ||
| Egg | 1676 | ||
| Rice | 1618 | ||
| Corn flour | 1300 | ||
| Milk | 687 | ||
| Tomato puree | 685 | ||
| Luteolin | Juniper berries | 69.05 | |
| Paprika (green) | 4.71 | ||
| Celery hearts (green) | 3.5 | ||
| Artichokes | 2.3 | ||
| Chicorée | 2.08 | ||
| Lemon | 1.9 | ||
| Pumpkin | 1.63 | ||
| Grapes (red) | 1.3 | ||
| Kohlrabi (raw) | 1.3 | ||
| Parsley (fresh) | 1.09 | ||
| Paprika (yellow) | 1.02 | ||
| Kiwi | 0.74 | ||
| Paprika (red) | 0.61 | ||
| Quercetin | Capers (raw) | 234 | Daily consumption of 25–50 mg [81] |
| Capers (canned) | 173 | ||
| Lovage leaves (raw) | 170 | ||
| Buckwheat seeds | 90 | ||
| Dock-like sorrel | 86 | ||
| Radish leaves | 70 | ||
| Carob fiber | 58 | ||
| Dill | 55 | ||
| Cilantro | 53 | ||
| Hungarian wax pepper | 51 | ||
| Fennel leaves | 49 | ||
| Onion (red) | 32 | ||
| Radicchio | 32 | ||
| Watercress | 30 | ||
| Kale | 23 | ||
| Chokeberry | 19 | ||
| Bog blueberry | 18 | ||
| Cranberry | 15 | ||
| Lingonberry | 13 | ||
| Plums (black) | 12 | ||
| Serine | Peanuts | 1862 | |
| Cheese (emmentaler) | 1749 | ||
| Soybeans | 1690 | ||
| Cheese (gouda) | 1570 | ||
| Lima beans | 1520 | ||
| Lentils | 1510 | ||
| Fish (plaice) | 1210 | ||
| Fish (tuna) | 1050 | ||
| Bacon | 1020 | ||
| Walnuts | 898 | ||
| g/100g | |||
| Methionine | Egg (white, dried, powder) | 3.204 | 19 mg/kg body weight/day [82] |
| Sesame seed flour | 1.656 | ||
| Brazil nuts | 1.124 | ||
| Cheese (parmesan) | 1.114 | ||
| Hemp seed | 0.933 | ||
| Soy protein concentrate | 0.814 | ||
| Chicken | 0.801 | ||
| Fish (tuna) | 0.755 | ||
| Beef | 0.749 | ||
| Bacon | 0.593 | ||
| Chia seed | 0.588 | ||
| Beef | 0.565 | ||
| Pork | 0.564 | ||
| Soybeans | 0.547 | ||
| Wheat germ | 0.456 | ||
| Egg (cooked) | 0.392 | ||
| Oat | 0.312 | ||
| Peanuts | 0.309 | ||
| Chickpea | 0.253 | ||
| Corn (yellow) | 0.197 | ||
| Almonds | 0.151 | ||
| Beans (pinto, cooked) | 0.117 | ||
| Lentils (cooked) | 0.077 | ||
| Rice (brown, cooked) | 0.052 | ||
| mg/kg | |||
| Spermidine | Wheat germ | 243 | |
| Soybean (dried) | 207 | ||
| Cheese (cheddar) | 199 | ||
| Mushroom | 89 | ||
| Rice bran | 50 | ||
| Chicken liver | 48 | ||
| Green peas | 46 | ||
| Mango | 30 | ||
| Chickpea | 29 | ||
| Cauliflower (cooked) | 25 | ||
| Broccoli (cooked) | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erichsen, L.; Thimm, C.; Santourlidis, S. Methyl Group Metabolism in Differentiation, Aging, and Cancer. Int. J. Mol. Sci. 2022, 23, 8378. https://doi.org/10.3390/ijms23158378
Erichsen L, Thimm C, Santourlidis S. Methyl Group Metabolism in Differentiation, Aging, and Cancer. International Journal of Molecular Sciences. 2022; 23(15):8378. https://doi.org/10.3390/ijms23158378
Chicago/Turabian StyleErichsen, Lars, Chantelle Thimm, and Simeon Santourlidis. 2022. "Methyl Group Metabolism in Differentiation, Aging, and Cancer" International Journal of Molecular Sciences 23, no. 15: 8378. https://doi.org/10.3390/ijms23158378