
Unraveling the Role of Histone Variant CENP-A and Chaperone HJURP Expression in Thymic Epithelial Neoplasms

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
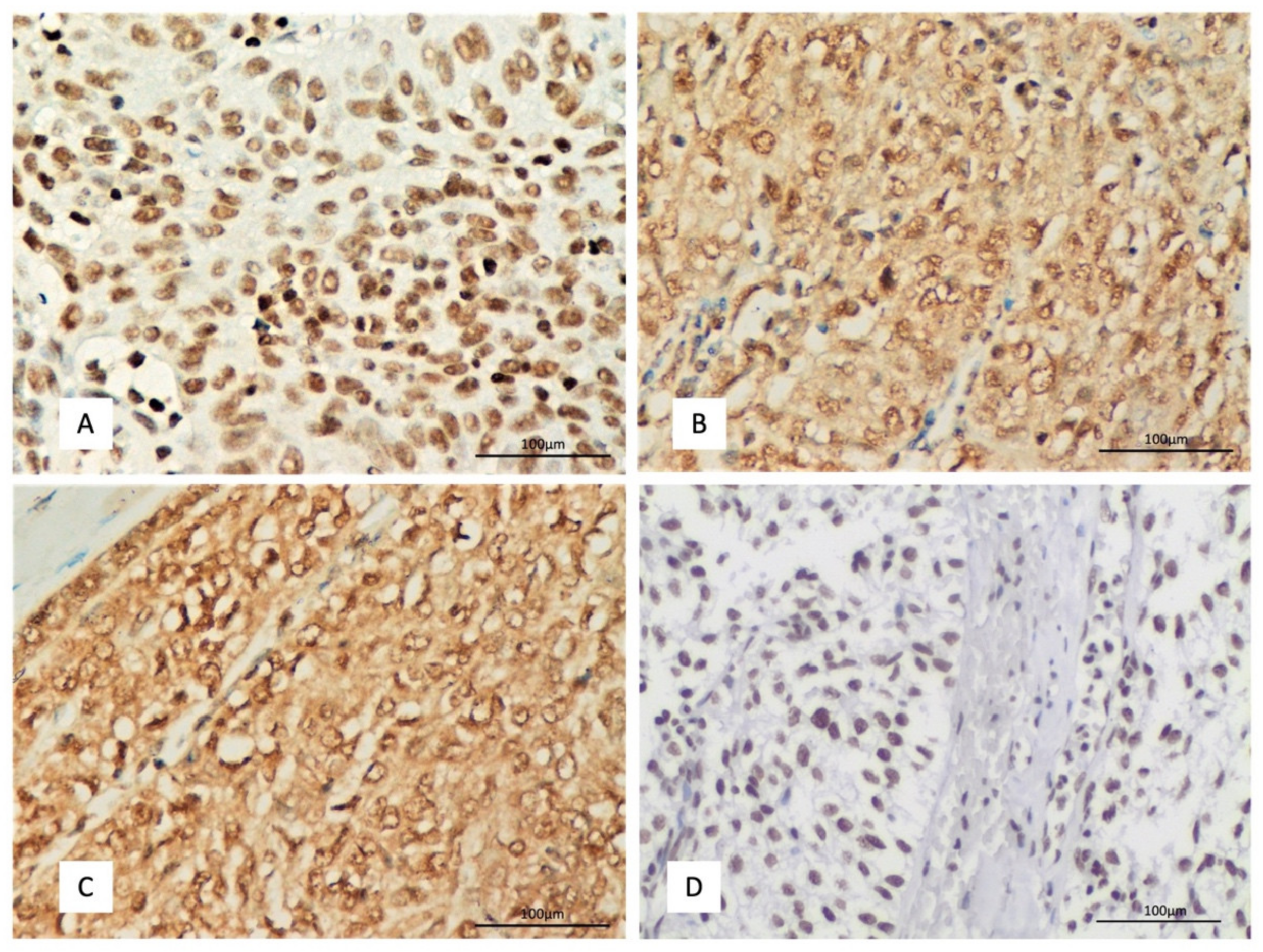
2.1. DAXX Expression and Associations with Clinicopathological Features
2.2. CENP-A Expression and Associations with Clinicopathological Features
2.3. HJURP Expression and Associations with Clinicopathological Features
2.4. H3K4me3 Expression and Associations with Clinicopathological Features
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. TMA Construction
4.3. Immunohistochemistry
4.4. Database and TCGA Analysis
4.4.1. Gene Expression Profiling Interactive Analysis (GEPIA)
4.4.2. Tumor Immune Estimation Resource (TIMER2.0) Analysis
4.4.3. Genetic-Copy Number Alteration and Mutation Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baylin, S.B.; Jones, P.A. A decade of exploring the cancer epigenome—Biological and translational implications. Nat. Cancer 2011, 11, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Ray-Gallet, D.; Almouzni, G. H3-H4 Histone Chaperones and Cancer. Curr. Opin. Genet. Dev. 2022, 73, 101900. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca, R.; Gurard-Levin, Z.A.; Berger, F.; Rehman, H.; Martel, E.; Corpet, A.; de Koning, L.; Vassias, I.; Wilson, L.O.W.; Meseure, D.; et al. The Histone Chaperone HJURP Is a New Independent Prognostic Marker for Luminal A Breast Carcinoma. Mol. Oncol. 2015, 9, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Arnaudo, A.M.; Garcia, B.A. Proteomic Characterization of Novel Histone Post-Translational Modifications. Epigenet. Chromatin 2013, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zang, C.; Rosenfeld, J.A.; Schones, D.E.; Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Peng, W.; Zhang, M.Q.; et al. Combinatorial Patterns of Histone Acetylations and Methylations in the Human Genome. Nat. Genet. 2008, 40, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Cooper, S.; Brockdorff, N. The Interplay of Histone Modifications-Writers That Read. EMBO Rep. 2015, 16, 1467–1481. [Google Scholar] [CrossRef]
- Franklin, S.G.; Zweidler, A. Non-Allelic Variants of Histones 2a, 2b and 3 in Mammals. Nature 1977, 266, 273–275. [Google Scholar] [CrossRef]
- Malik, H.S.; Henikoff, S. Phylogenomics of the Nucleosome. Nat. Struct. Mol. Biol. 2003, 10, 882–891. [Google Scholar] [CrossRef]
- Smith, S.; Stillman, B. Purification and Characterization of CAF-I, a Human Cell Factor Required for Chromatin Assembly during DNA Replication in Vitro. Cell 1989, 58, 15–25. [Google Scholar] [CrossRef]
- Müller, S.; Almouzni, G. Chromatin Dynamics during the Cell Cycle at Centromeres. Nat. Rev. Genet. 2017, 18, 192–208. [Google Scholar] [CrossRef]
- Ray-Gallet, D.; Almouzni, G. The Histone H3 Family and Its Deposition Pathways. Adv. Exp. Med. Biol. 2021, 1283, 17–42. [Google Scholar] [CrossRef]
- Dunleavy, E.M.; Roche, D.; Tagami, H.; Lacoste, N.; Ray-Gallet, D.; Nakamura, Y.; Daigo, Y.; Nakatani, Y.; Almouzni-Pettinotti, G. HJURP Is a Cell-Cycle-Dependent Maintenance and Deposition Factor of CENP-A at Centromeres. Cell 2009, 137, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Foltz, D.R.; Jansen, L.E.T.; Bailey, A.O.; Yates, J.R.; Bassett, E.A.; Wood, S.; Black, B.E.; Cleveland, D.W. Centromere-Specific Assembly of CENP-a Nucleosomes Is Mediated by HJURP. Cell 2009, 137, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Sitbon, D.; Podsypanina, K.; Yadav, T.; Almouzni, G. Shaping Chromatin in the Nucleus: The Bricks and the Architects. Cold Spring Harb. Symp. Quant. Biol. 2017, 82, 1–14. [Google Scholar] [CrossRef]
- Arimura, Y.; Shirayama, K.; Horikoshi, N.; Fujita, R.; Taguchi, H.; Kagawa, W.; Fukagawa, T.; Almouzni, G.; Kurumizaka, H. Crystal Structure and Stable Property of the Cancer-Associated Heterotypic Nucleosome Containing CENP-A and H3.3. Sci. Rep. 2014, 4, 7115. [Google Scholar] [CrossRef] [Green Version]
- Rashid, O.M.; Cassano, A.D.; Takabe, K. Thymic Neoplasm: A Rare Disease with a Complex Clinical Presentation. J. Thorac. Dis. 2013, 5, 173–183. [Google Scholar] [CrossRef]
- Marx, A.; Chan, J.K.C.; Chalabreysse, L.; Dacic, S.; Detterbeck, F.; French, C.A.; Hornick, J.L.; Inagaki, H.; Jain, D.; Lazar, A.J.; et al. The 2021 WHO Classification of Tumors of the Thymus and Mediastinum: What Is New in Thymic Epithelial, Germ Cell, and Mesenchymal Tumors? J. Thorac. Oncol. 2022, 17, 200–213. [Google Scholar] [CrossRef]
- WHO Classification of Tumours Editiorial Board. Thoracic Tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2021; Volume 5. [Google Scholar]
- Psilopatis, I.; Pergaris, A.; Vrettou, K.; Theocharis, S.; Troungos, C. Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations. Int. J. Mol. Sci. 2022, 23, 4045. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Z.; Zhang, S.; Yu, D.; Yu, H.; Liu, L.; Cao, X.; Wang, L.; Gao, H.; Zhu, M. ShRNA-Targeted Centromere Protein A Inhibits Hepatocellular Carcinoma Growth. PLoS ONE 2011, 6, e17794. [Google Scholar] [CrossRef] [Green Version]
- Tomonaga, T.; Matsushita, K.; Yamaguchi, S.; Oohashi, T.; Shimada, H.; Ochiai, T.; Yoda, K.; Nomura, F. Overexpression and Mistargeting of Centromere Protein-A in Human Primary Colorectal Cancer. Cancer Res. 2003, 63, 3511–3516. [Google Scholar]
- McGovern, S.L.; Qi, Y.; Pusztai, L.; Symmans, W.F.; Buchholz, T.A. Centromere Protein-A, an Essential Centromere Protein, Is a Prognostic Marker for Relapse in Estrogen Receptor-Positive Breast Cancer. Breast Cancer Res. 2012, 14, R72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Qian, Y.-M.; Zhao, X.-L.; Wang, S.-M.; Feng, X.-J.; Chen, X.-F.; Zhang, S.-H. Expression and Prognostic Significance of Centromere Protein A in Human Lung Adenocarcinoma. Lung Cancer 2012, 77, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.-J.; Guo, J.-J.; Lv, T.-J.; Jin, H.-Y.; Ding, J.-X.; Feng, W.-W.; Zhang, Y.; Hua, K.-Q. Prognostic Value of Centromere Protein-A Expression in Patients with Epithelial Ovarian Cancer. Tumour Biol. 2013, 34, 2971–2975. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Huang, G.; Sadanandam, A.; Gu, S.; Lenburg, M.E.; Pai, M.; Bayani, N.; Blakely, E.A.; Gray, J.W.; Mao, J.-H. The Expression Level of HJURP Has an Independent Prognostic Impact and Predicts the Sensitivity to Radiotherapy in Breast Cancer. Breast Cancer Res. 2010, 12, R18. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-F.; Liang, Y.-X.; Yang, J.-A.; Yuan, D.-Z.; Li, J.; Zheng, S.-S.; Wan, Y.-P.; Wang, B.; Han, Z.-D.; Zhong, W.-D. Upregulation of Holliday Junction Recognition Protein Predicts Poor Prognosis and Biochemical Recurrence in Patients with Prostate Cancer. Oncol. Lett. 2019, 18, 6697–6703. [Google Scholar] [CrossRef] [Green Version]
- Valente, V.; Serafim, R.B.; de Oliveira, L.C.; Adorni, F.S.; Torrieri, R.; Tirapelli, D.P.D.C.; Espreafico, E.M.; Oba-Shinjo, S.M.; Marie, S.K.N.; Paçó-Larson, M.L.; et al. Modulation of HJURP (Holliday Junction-Recognizing Protein) Levels Is Correlated with Glioblastoma Cells Survival. PLoS ONE 2013, 8, e62200. [Google Scholar] [CrossRef]
- Mahmud, I.; Liao, D. DAXX in Cancer: Phenomena, Processes, Mechanisms and Regulation. Nucleic Acids Res. 2019, 47, 7734–7752. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.; Zhu, W.; Xiao, C.; Li, X.; Wang, Y.; Han, Y.; Zheng, J.; Li, Y.; Li, M.; Wen, X. HJURP Promotes Proliferation in Prostate Cancer Cells through Increasing CDKN1A Degradation via the GSK3β/JNK Signaling Pathway. Cell Death Dis. 2021, 12, 583. [Google Scholar] [CrossRef]
- Chen, T.; Huang, H.; Zhou, Y.; Geng, L.; Shen, T.; Yin, S.; Zhou, L.; Zheng, S. HJURP Promotes Hepatocellular Carcinoma Proliferation by Destabilizing P21 via the MAPK/ERK1/2 and AKT/GSK3β Signaling Pathways. J. Exp. Clin. Cancer Res. 2018, 37, 193. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Zhou, L.; Zhou, Y.; Zhou, W.; Huang, H.; Yin, S.; Xie, H.; Zhou, L.; Zheng, S. HJURP Promotes Epithelial-to-Mesenchymal Transition via Upregulating SPHK1 in Hepatocellular Carcinoma. Int. J. Biol. Sci. 2019, 15, 1139–1147. [Google Scholar] [CrossRef]
- De Tayrac, M.; Saikali, S.; Aubry, M.; Bellaud, P.; Boniface, R.; Quillien, V.; Mosser, J. Prognostic Significance of EDN/RB, HJURP, P60/CAF-1 and PDLI4, Four New Markers in High-Grade Gliomas. PLoS ONE 2013, 8, e73332. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-J.; Li, X.; Shi, P.; Ding, H.-Y.; Liu, Y.-P.; Li, T.; Lin, P.-P.; Wang, Y.-S.; Zhang, G.-Q.; Cao, Y. Holliday Junction Recognition Protein Promotes Pancreatic Cancer Growth and Metastasis via Modulation of the MDM2/P53 Signaling. Cell Death Dis. 2020, 11, 386. [Google Scholar] [CrossRef]
- Kang, D.H.; Woo, J.; Kim, H.; Kim, S.Y.; Ji, S.; Jaygal, G.; Ahn, T.S.; Kim, H.J.; Kwak, H.J.; Kim, C.-J.; et al. Prognostic Relevance of HJURP Expression in Patients with Surgically Resected Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 7928. [Google Scholar] [CrossRef]
- Verrelle, P.; Meseure, D.; Berger, F.; Forest, A.; Leclère, R.; Nicolas, A.; Fortas, E.; Sastre-Garau, X.; Lae, M.; Boudjemaa, S.; et al. CENP-A Subnuclear Localization Pattern as Marker Predicting Curability by Chemoradiation Therapy for Locally Advanced Head and Neck Cancer Patients. Cancers 2021, 13, 3928. [Google Scholar] [CrossRef]
- Jing, R.; Xi, J.; Leng, Y.; Chen, W.; Wang, G.; Jia, W.; Kang, J.; Zhu, S. Motifs in the Amino-Terminus of CENP-A Are Required for Its Accumulation within the Nucleus and at the Centromere. Oncotarget 2017, 8, 40654–40667. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Xu, J.; Zhang, J.; Xie, D.; Ye, H.; Xiao, Z.; Cai, M.; Xu, K.; Zeng, Y.; Li, H.; et al. High Expression of Trimethylated Histone H3 Lysine 4 Is Associated with Poor Prognosis in Hepatocellular Carcinoma. Hum. Pathol. 2012, 43, 1425–1435. [Google Scholar] [CrossRef]
- Beyer, S.; Zhu, J.; Mayr, D.; Kuhn, C.; Schulze, S.; Hofmann, S.; Dannecker, C.; Jeschke, U.; Kost, B.P. Histone H3 Acetyl K9 and Histone H3 Tri Methyl K4 as Prognostic Markers for Patients with Cervical Cancer. Int. J. Mol. Sci. 2017, 18, 477. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, H.; Chen, H.; Zavadil, J.; Kluz, T.; Arita, A.; Costa, M. Hypoxia Induces Trimethylated H3 Lysine 4 by Inhibition of JARID1A Demethylase. Cancer Res. 2010, 70, 4214–4221. [Google Scholar] [CrossRef] [Green Version]
- Berger, L.; Kolben, T.; Meister, S.; Kolben, T.M.; Schmoeckel, E.; Mayr, D.; Mahner, S.; Jeschke, U.; Ditsch, N.; Beyer, S. Expression of H3K4me3 and H3K9ac in Breast Cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 2017–2027. [Google Scholar] [CrossRef]
- Ellinger, J.; Kahl, P.; Mertens, C.; Rogenhofer, S.; Hauser, S.; Hartmann, W.; Bastian, P.J.; Büttner, R.; Müller, S.C.; von Ruecker, A. Prognostic Relevance of Global Histone H3 Lysine 4 (H3K4) Methylation in Renal Cell Carcinoma. Int. J. Cancer 2010, 127, 2360–2366. [Google Scholar] [CrossRef]
- Schneider, A.-C.; Heukamp, L.C.; Rogenhofer, S.; Fechner, G.; Bastian, P.J.; von Ruecker, A.; Müller, S.C.; Ellinger, J. Global Histone H4K20 Trimethylation Predicts Cancer-Specific Survival in Patients with Muscle-Invasive Bladder Cancer. BJU Int. 2011, 108, E290–E296. [Google Scholar] [CrossRef]
- Shrestha, R.L.; Rossi, A.; Wangsa, D.; Hogan, A.K.; Zaldana, K.S.; Suva, E.; Chung, Y.J.; Sanders, C.L.; Difilippantonio, S.; Karpova, T.S.; et al. CENP-A Overexpression Promotes Aneuploidy with Karyotypic Heterogeneity. J. Cell Biol. 2021, 220, e202007195. [Google Scholar] [CrossRef]
- Mahlke, M.A.; Nechemia-Arbely, Y. Guarding the Genome: CENP-A-Chromatin in Health and Cancer. Genes 2020, 11, E810. [Google Scholar] [CrossRef]
- Arbour, K.C.; Naidoo, J.; Steele, K.E.; Ni, A.; Moreira, A.L.; Rekhtman, N.; Robbins, P.B.; Karakunnel, J.; Rimner, A.; Huang, J.; et al. Expression of PD-L1 and Other Immunotherapeutic Targets in Thymic Epithelial Tumors. PLoS ONE 2017, 12, e0182665. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for Analysis of Tumor-Infiltrating Immune Cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- John, G.T.; Sally, B.; Harry, C.J.; Zbyslaw, S.; David, M.B.; Nidhi, B.; Harry, B.; Charlotte, G.C.; Celestino, C.; Elisabeth, D.; et al. COSMIC: The catalogue of somatic mutations in cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Positivity Rate | H-Score, Median | H-Score, Range | |
|---|---|---|---|
| Epithelial cells | |||
| DAXX nuclear expression | 93.6% | 90 | 0–300 |
| CENP-A cytoplasmic expression | 90% | 50 | 0–300 |
| CENP-A nuclear expression | 39.8% | 0 | 0–160 |
| HJURP cytoplasmic expression | 69% | 35 | 0–300 |
| HJURP nuclear expression | 99% | 120 | 0–300 |
| H3K4me3 nuclear expression | 100% | 100 | 10–294 |
| Lymphoid cells | |||
| DAXX | 95.7% | 200 | 0–300 |
| CENP-A cytoplasmic expression | 98.6% | 140 | 0–300 |
| HJURP nuclear expression | 98.3% | 235 | 0–300 |
| H3K4me3 nuclear expression | 100% | 270 | 0–300 |
| Parameter | Median | Range |
|---|---|---|
| Age | 63 | 27–88 years |
| Number | % | |
| Gender | ||
| Male | 42/95 | 44.2% |
| Female | 53/95 | 55.8% |
| WHO subtypes | ||
| Type A | 11/95 | 11.6% |
| Type AB | 22/95 | 23.2% |
| Type B1 | 17/95 | 17.9% |
| Type B2 | 18/95 | 19.0% |
| Type B3 | 13/95 | 13.7% |
| Micronodular with lymphoid stroma | 2/95 | 2% |
| Type C | 12/95 | 12.6% |
| Masaoka–Koga stage | ||
| I | 17/85 | 20% |
| IIa | 34/85 | 40% |
| IIb | 14/85 | 16.5% |
| III | 14/85 | 16.5% |
| IVa | 3/85 | 3.5% |
| IVb | 3/85 | 3.5% |
| Positive surgical margins | 13/46 | 28.3% |
| Presence of myasthenia Gravis | 33/56 | 58.9% |
| Presence of chemotherapy | 10/37 | 27% |
| Presence of radiotherapy | 18/36 | 50% |
| Event | ||
| Alive disease free | 25/38, follow-up 5–134 months | 71.4% |
| Alive with disease | 3/38, follow-up 28–65 months | 8.6% |
| Dead of disease | 7/38, within 11–65 months | 20% |
| Presence of relapse | 3/33, within 58–65 months | 9.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levidou, G.; Palamaris, K.; Sykaras, A.G.; Andreadakis, G.; Masaoutis, C.; Theochari, I.; Korkolopoulou, P.; Rontogianni, D.; Theocharis, S. Unraveling the Role of Histone Variant CENP-A and Chaperone HJURP Expression in Thymic Epithelial Neoplasms. Int. J. Mol. Sci. 2022, 23, 8339. https://doi.org/10.3390/ijms23158339
Levidou G, Palamaris K, Sykaras AG, Andreadakis G, Masaoutis C, Theochari I, Korkolopoulou P, Rontogianni D, Theocharis S. Unraveling the Role of Histone Variant CENP-A and Chaperone HJURP Expression in Thymic Epithelial Neoplasms. International Journal of Molecular Sciences. 2022; 23(15):8339. https://doi.org/10.3390/ijms23158339
Chicago/Turabian StyleLevidou, Georgia, Konstantinos Palamaris, Alexandros G. Sykaras, Georgios Andreadakis, Christos Masaoutis, Irene Theochari, Penelope Korkolopoulou, Dimitra Rontogianni, and Stamatios Theocharis. 2022. "Unraveling the Role of Histone Variant CENP-A and Chaperone HJURP Expression in Thymic Epithelial Neoplasms" International Journal of Molecular Sciences 23, no. 15: 8339. https://doi.org/10.3390/ijms23158339