
Investigation of the Molecular Mechanisms Underlying the Antiatherogenic Actions of Kaempferol in Human THP-1 Macrophages


Abstract
:1. Introduction
2. Results
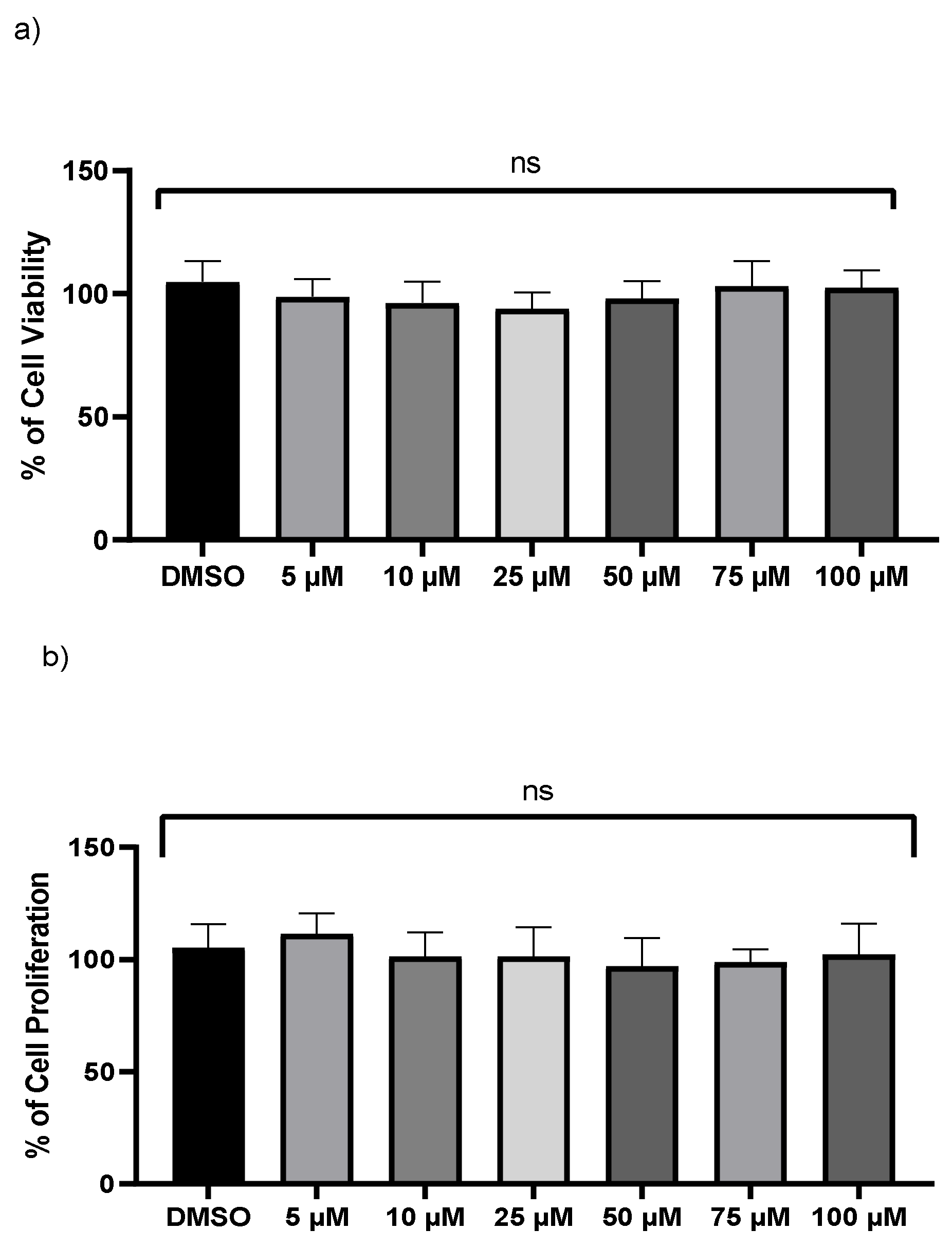
2.1. Influence of Kaempferol on the Survival of THP-1 Macrophages
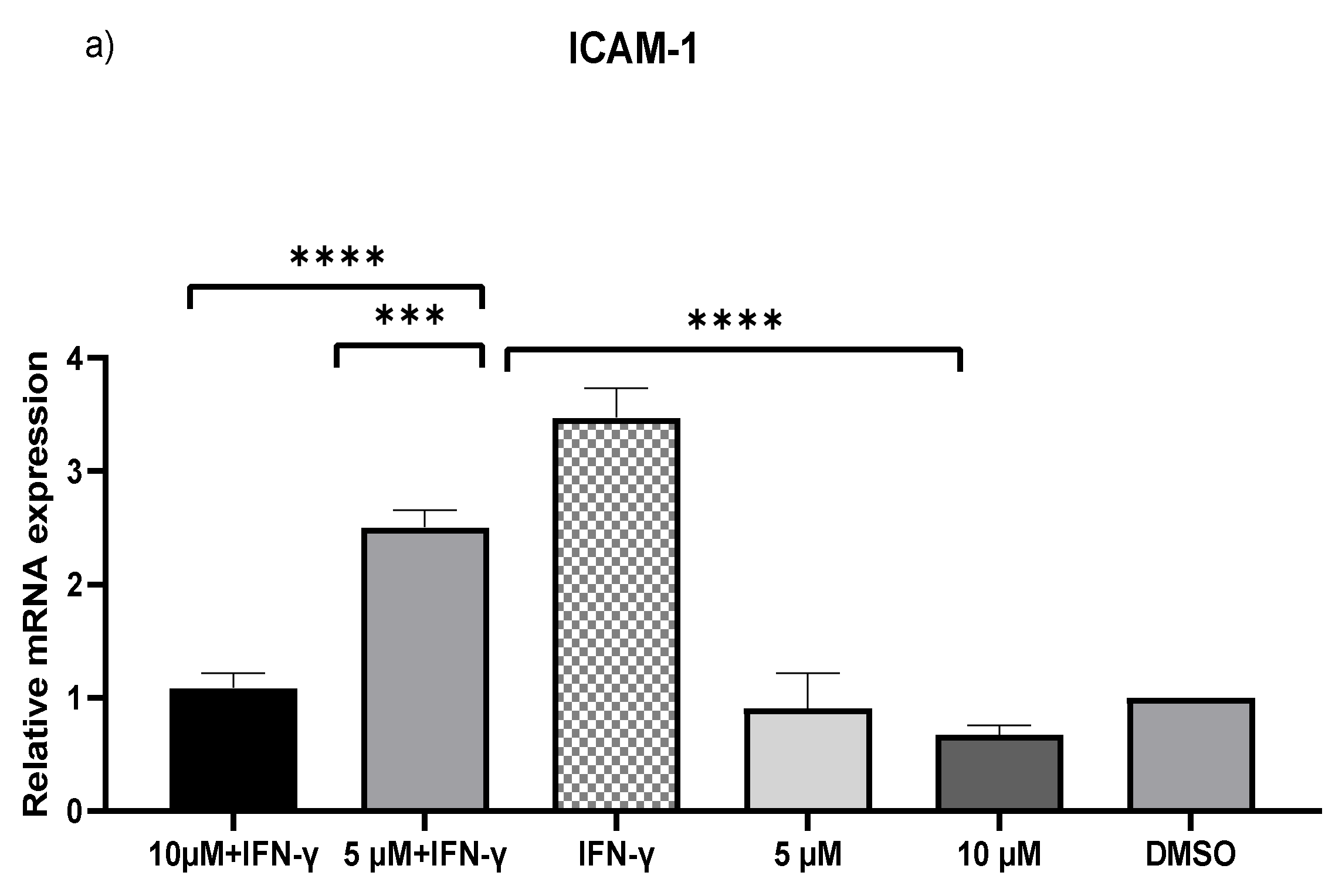
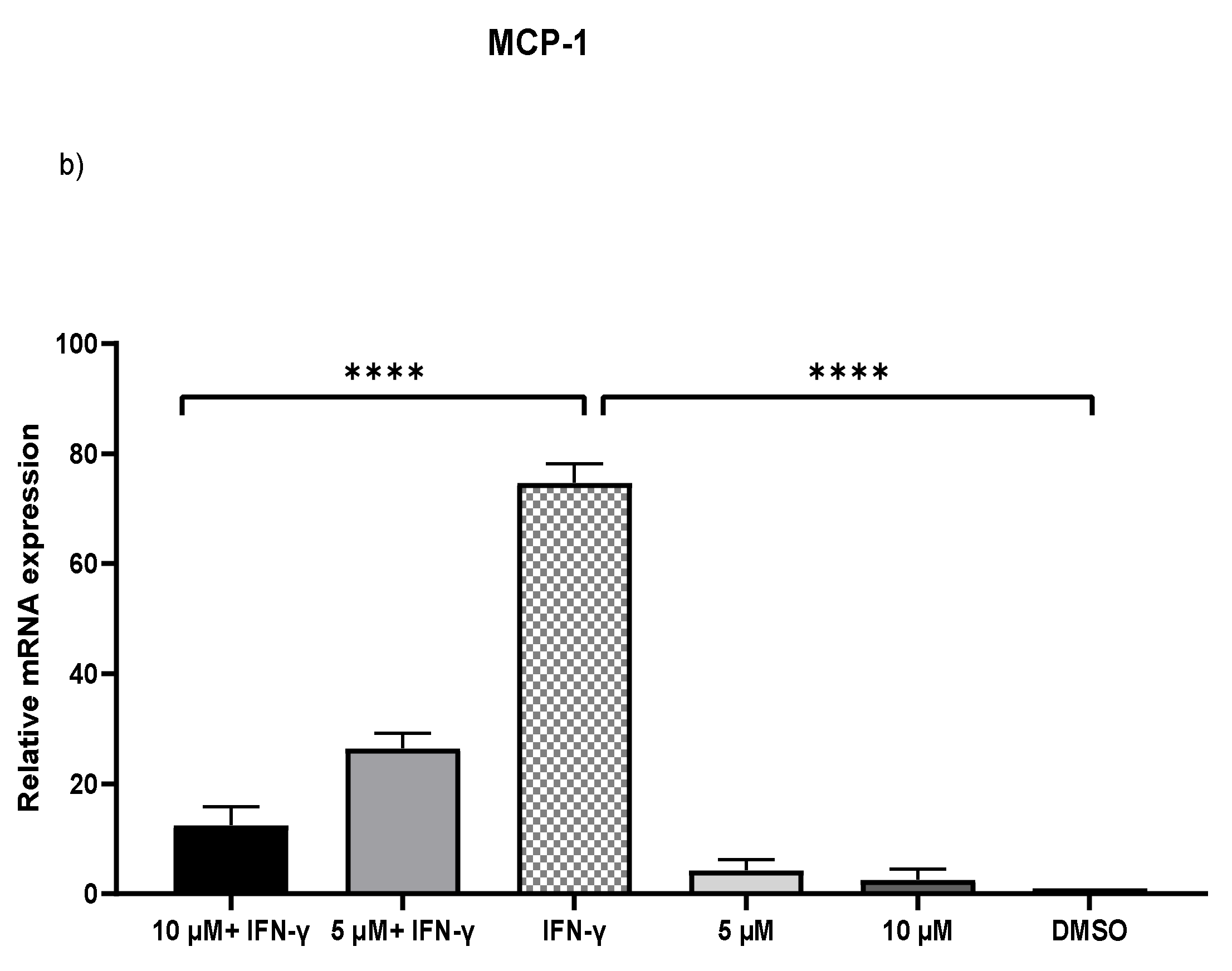
2.2. Downregulation of MCP-1 and ICAM-1 Expression by Kaempferol
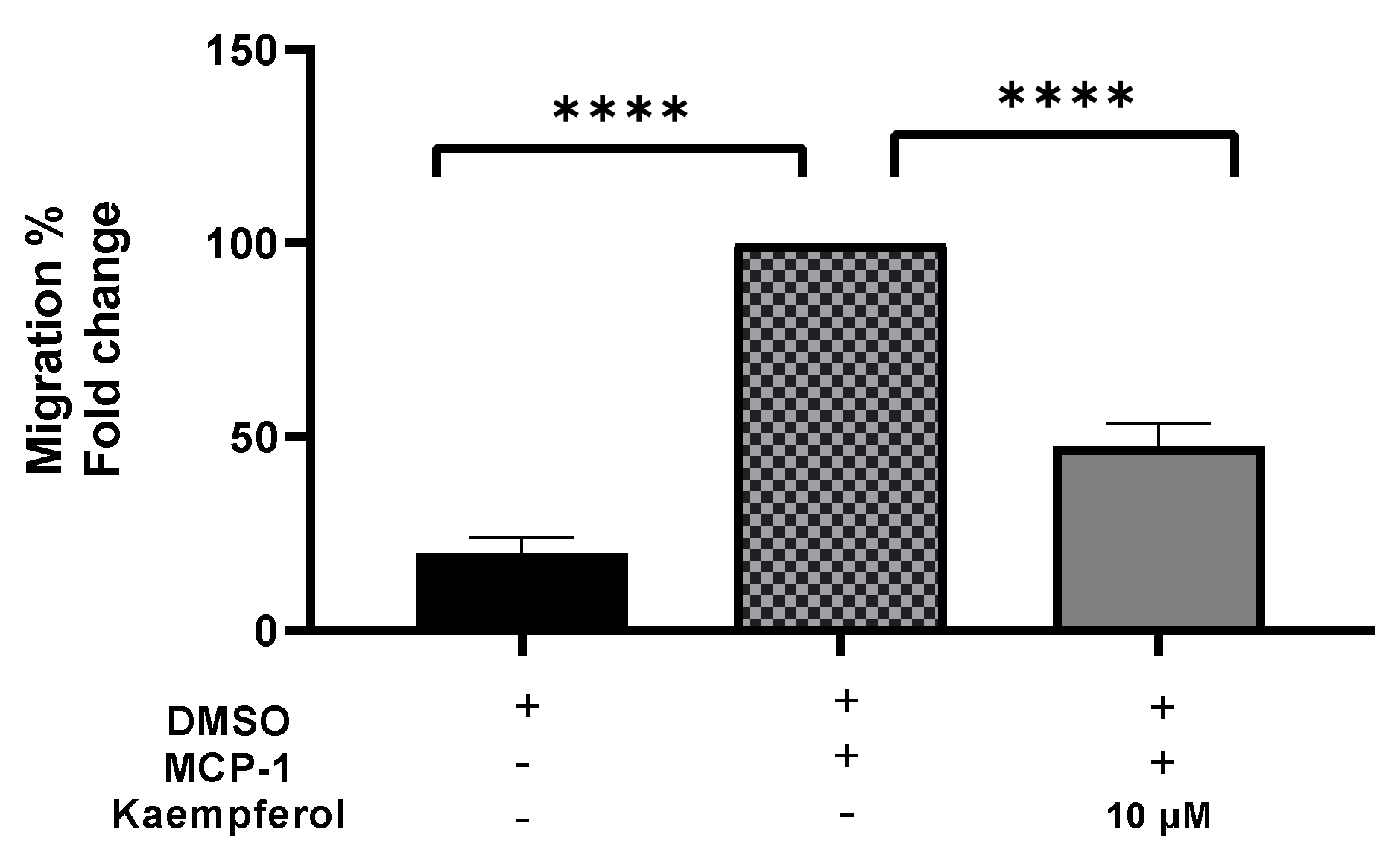
2.3. Inhibition of the Transferring of Monocytes by Kaempferol
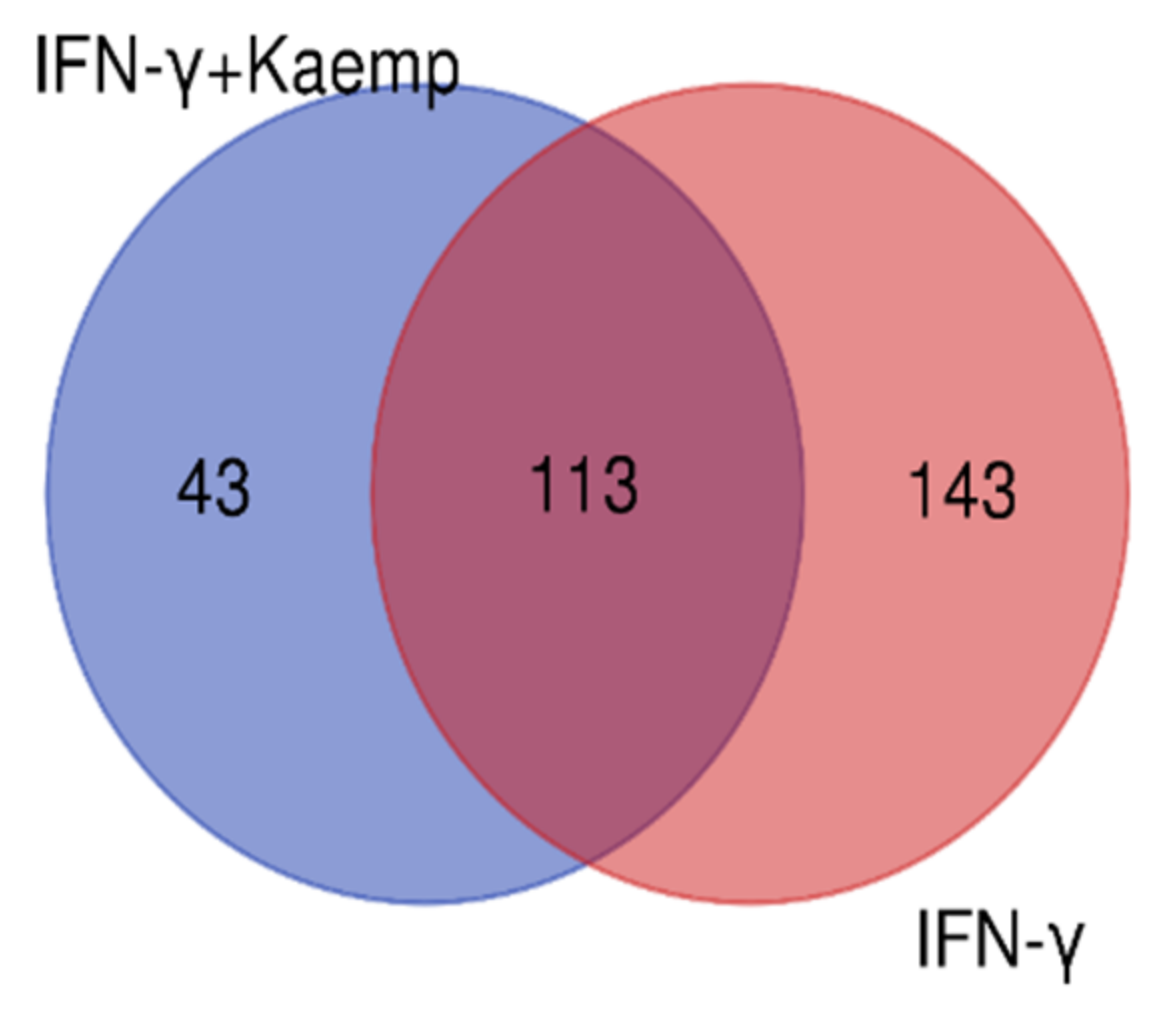
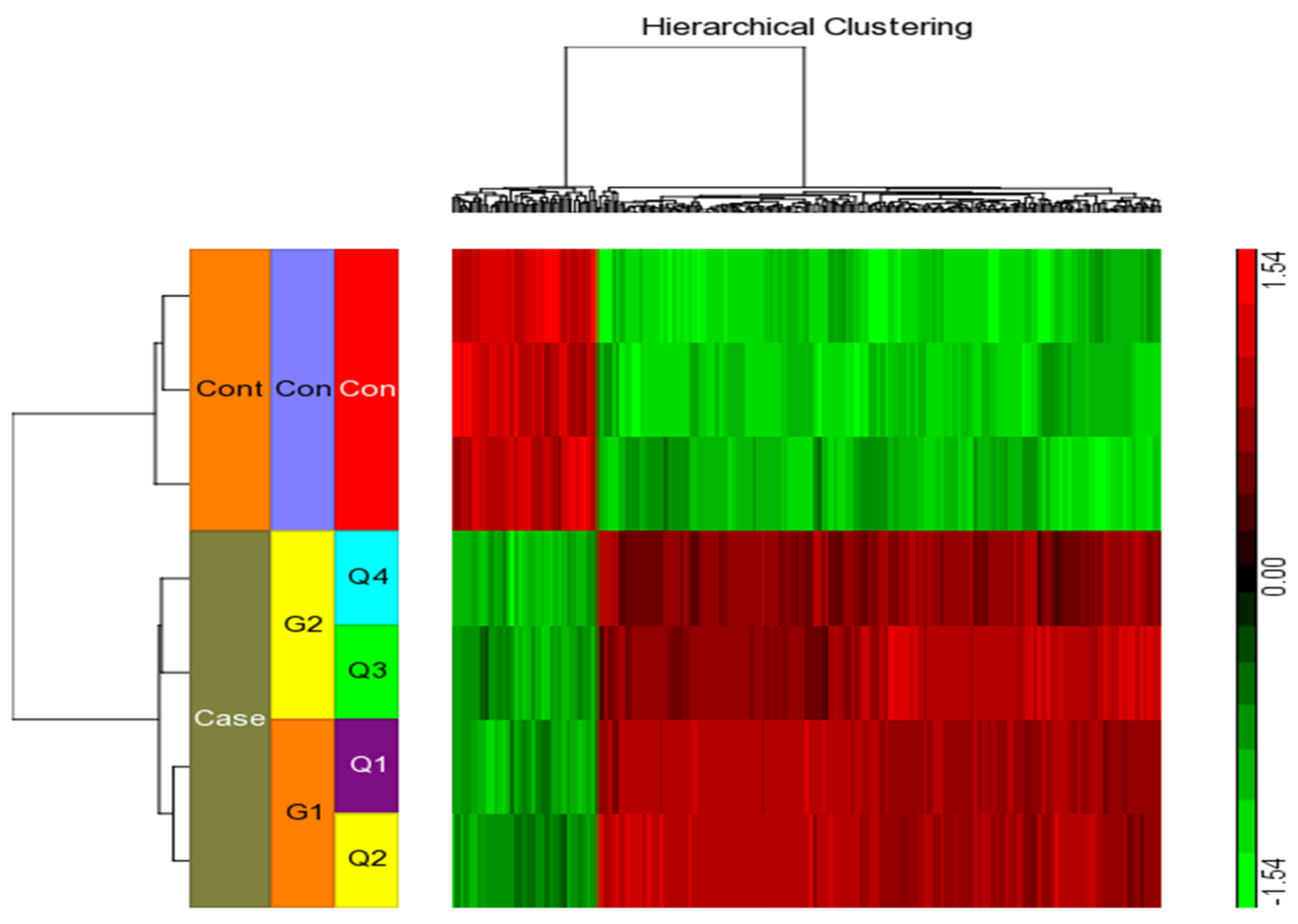
2.4. Differential Gene Expression Profiles in Differentiated THP-1 Are Induced by Kaempferol
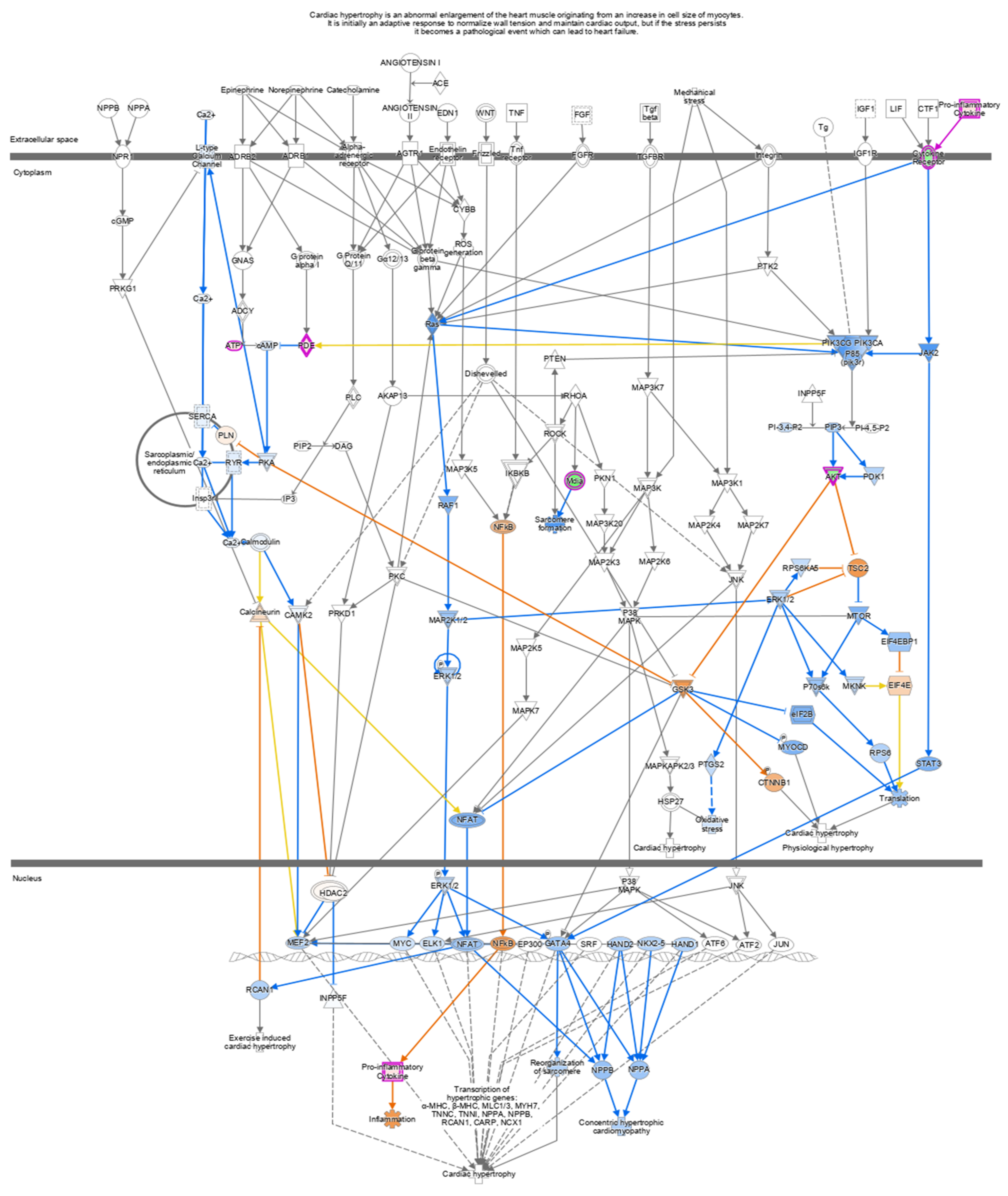
2.5. Ingenuity Pathway Analysis
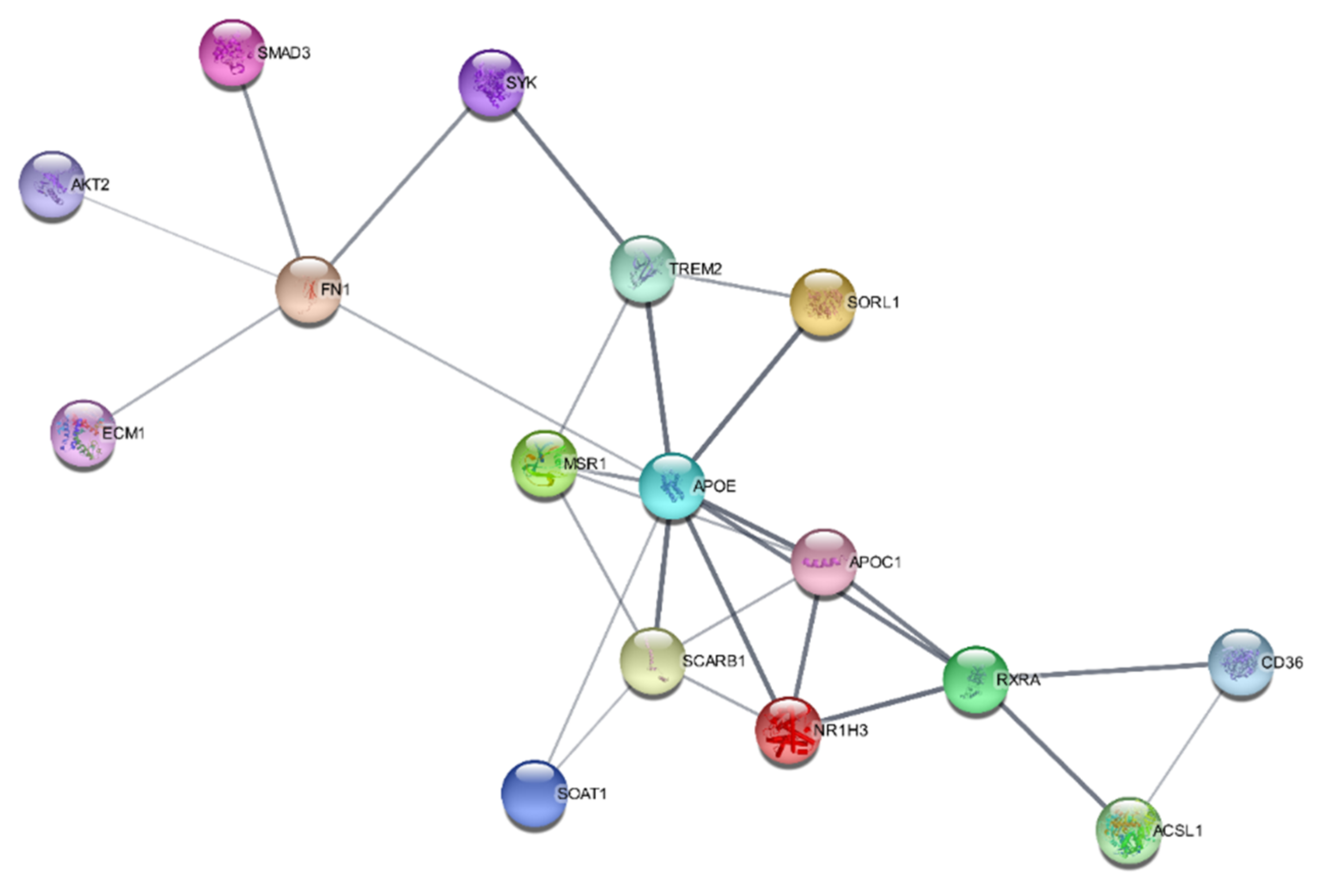
2.6. SwissTargetPrediction and Protein–Protein Interaction Networks
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Chemicals and Reagents
4.3. THP-1 Cell Culture
4.4. Lactate Dehydrogenase Assay
4.5. Crystal Violet Assay
4.6. In Vitro Monocyte Migration Assay
4.7. Microarray Processing and Gene Expression Analysis
4.8. Quantitative Real-Time Polymerase Chain Reaction
4.9. Ingenuity Pathway Analysis
4.10. In Silico Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lusis, A.J. Atherosclerosis. Nature 2002, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.; Michael, D.; Ashlin, T.; Ramji, D. Cytokines, macrophage lipid metabolism and foam cells: Implications for car-diovascular disease therapy. Prog. Lipid Res. 2011, 50, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Ramji, D.P.; Davies, T.S. Cytokines in atherosclerosis: Key players in all stages of disease and promising therapeutic targets. Cytokine Growth Factor Rev. 2015, 26, 673–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, D.K.; Pitts-Meek, S.; Keshav, S.; Figari, I.S.; Bradley, A.; Stewart, T.A. Multiple defects of immune cell function in mice with disrupted interferon-gamma genes. Science 1993, 259, 1739–1742. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K.; Leducq Transatlantic Network on Atherothrombosis. Inflammation in Atherosclerosis: From Pathophysiology to Practice. J. Am. Coll. Cardiol. 2009, 54, 2129–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delavary, B.M.; van der Veer, W.M.; van Egmond, M.; Niessen, F.B.; Beelen, R.H. Macrophages in skin injury and repair. Immunobiology 2011, 216, 753–762. [Google Scholar] [CrossRef]
- Komohara, Y.; Fujiwara, Y.; Ohnishi, K.; Shiraishi, D.; Takeya, M. Contribution of Macrophage Polarization to Metabolic Diseases. J. Atheroscler. Thromb. 2016, 23, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Zhang, X.; Ma, Z.; Niu, J.; Ma, S.; Wenjie, W.; Chen, J. Combination of tanshinone IIA and astragaloside IV attenuate atherosclerotic plaque vulnerability in ApoE(−/−) mice by activating PI3K/AKT signaling and suppressing TRL4/NF-κB signaling. Biomed. Pharmacother 2020, 123, 109729. [Google Scholar] [CrossRef]
- Chen, F.; Guo, N.; Cao, G.; Zhou, J.; Yuan, Z. Molecular Analysis of Curcumin-induced Polarization of Murine RAW264.7 Macrophages. J. Cardiovasc. Pharmacol. 2014, 63, 544–552. [Google Scholar] [CrossRef]
- Endo, A.; Tsujita, Y.; Kuroda, M.; Tanzawa, K. Inhibition of cholesterol synthesis in vitro and in vivo by ML 236A and ML 236B, competitive inhibitors of 3 hydroxy 3 methylglutaryl Coenzyme A reductase. Eur. J. Biochem. 1977, 77, 31–36. [Google Scholar] [CrossRef]
- Klingenberg, R.; Hansson, G.K. Treating inflammation in atherosclerotic cardiovascular disease: Emerging therapies. Eur. Heart J. 2009, 30, 2838–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, D.; Lu, G.; Cai, N.; Wu, Z.G.; Li, Y.H.; Chen, H.; Zhu, J.Q.; Jin, X.J.; Wouters, B.C.; Zhao, J. Cholesterol-lowering effect of a theaflavin-enriched green tea extract: A randomized controlled trial. Arch. Intern. Med. 2003, 163, 1448–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huddy, K.; Dhesi, P.; Thompson, P.D. Do the frequencies of adverse events increase, decrease, or stay the same with long-term use of statins? Curr. Atheroscler. Rep. 2013, 15, 301. [Google Scholar] [CrossRef]
- Ayoub, M.; de Camargo, A.C.; Shahidi, F. Antioxidants and bioactivities of free, esterified, and insoluble-bound phenolics from berry seed meals. Food Chem. 2016, 197, 221–232. [Google Scholar] [CrossRef]
- Jaime, L.; Santoyo, S. The Health Benefits of the Bioactive Compounds in Foods. Foods 2021, 10, 325. [Google Scholar] [CrossRef]
- Pieretti, S.; Saviano, A.; Mollica, A.; Stefanucci, A.; Aloisi, A.M.; Nicoletti, M. Calceolarioside A, a Phenylpropanoid Glycoside from Calceolaria spp., Displays Antinociceptive and Anti-Inflammatory Properties. Molecules 2022, 27, 2183. [Google Scholar] [CrossRef]
- Dimmito, M.P.; Stefanucci, A.; Della Valle, A.; Scioli, G.; Cichelli, A.; Mollica, A. An overview on plants cannabinoids endorsed with cardiovascular effects. Biomed. Pharmacother. 2021, 142, 111963. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef]
- Ochiai, A.; Miyata, S.; Iwase, M.; Shimizu, M.; Inoue, J.; Sato, R. Kaempferol stimulates gene expression of low-density lipo-protein receptor through activation of Sp1 in cultured hepatocytes. Sci. Rep. 2016, 6, 24940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Sun, J.; Wei, J.; Zhang, X.; Chen, B.; Lin, Y. Kaempferol protects blood vessels from damage induced by oxidative stress and inflammation in association with the Nrf2/HO-1 signaling pathway. Front. Pharmacol. 2021, 22, 1118. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, S.; Geisler, S.; Lisandrelli, R.; Ngoc, H.N.; Ganzera, M.; Schennach, H.; Fuchs, D.; Fuchs, J.E.; Gostner, J.M.; Kurz, K. Pharmacological Targets of Kaempferol Within Inflammatory Pathways—A Hint Towards the Central Role of Tryptophan Metabolism. Antioxidants 2020, 9, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, J.; Ramji, D. Nutraceutical therapies for atherosclerosis. Nat. Rev. Cardiol. 2016, 13, 513–532. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; McLaren, J.E.; Michael, D.R.; Clement, M.; Fielding, C.A.; Ramji, D.P. ERK Is Integral to the IFN-γ–Mediated Activation of STAT1, the Expression of Key Genes Implicated in Atherosclerosis, and the Uptake of Modified Lipoproteins by Human Macrophages. J. Immunol. 2010, 185, 3041–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, J.; Ramji, P. Interferon gamma: A master regulator of atherosclerosis. Cytokine Growth Factor Rev. 2009, 20, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, J.; Samojedny, A.; Paul, M.; Pietsz, G.; Wilczok, T. Effect of apigenin, kaempferol and resveratrol on the expression of interleukin-1beta and tumor necrosis factor-alpha genes in J774.2 macrophages. Pharmacol. Rep. 2005, 57, 390–394. [Google Scholar]
- Kakkar, V.; Lu, X. Impact of MCP-1 in Atherosclerosis. Curr. Pharm. Des. 2014, 20, 4580–4588. [Google Scholar]
- Kang, H.; Li, X.; Xiong, K.; Song, Z.; Tian, J.; Wen, Y.; Sun, A.; Deng, X. The Entry and Egress of Monocytes in Atherosclerosis: A Biochemical and Biomechanical Driven Process. Cardiovasc. Ther. 2021, 2021, 6642927. [Google Scholar] [CrossRef]
- Bot, I.; Ortiz, N.; de Witte, W.; de Vries, H.; van Santbrink, P.J.; van der Velden, D.; Kröner, M.J.; van der Berg, D.J.; Stamos, D.; de Lange, E.C.; et al. A novel CCR2 antagonist inhibits atherogenesis in apoE deficient mice by achieving high receptor occupancy. Sci. Rep. 2017, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Zhang, W.; Wang, Z. Laquinimod Protects Against TNF-α-Induced Attachment of Monocytes to Human Aortic Endothelial Cells (HAECs) by Increasing the Expression of KLF2. Drug Des. Dev. Ther. 2020, 14, 1683–1691. [Google Scholar] [CrossRef]
- Yin, M.; Li, C.; Jiang, J.; Le, J.; Luo, B.; Yang, F.; Fang, Y.; Yang, M.; Deng, Z.; Ni, W.; et al. Cell adhesion molecule-mediated therapeutic strategies in atherosclerosis: From a biological basis and molecular mechanism to drug delivery nanosystems. Biochem. Pharmacol. 2021, 186, 114471. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Luo, C.; Li, X.; Zhou, Y.; He, H. The anti-inflammatory effect of kaempferol on early atherosclerosis in high cholesterol fed rabbits. Lipids Health Dis. 2013, 12, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielewski, S.; Olejnik, A.; Sikorski, K.; Pelisek, J.; Błaszczyk, K.; Aoqui, C.; Nowicka, H.; Zernecke, A.; Heemann, U.; Wesoly, J.; et al. STAT1-Dependent Signal Integration between IFNc and TLR4 in Vascular Cells Reflect Pro-Atherogenic Responses in Human Atherosclerosis. PLoS ONE 2014, 9, e113318. [Google Scholar] [CrossRef] [PubMed]
- Segers, D.; Lipton, J.A.; Leenen, P.J.M.; Cheng, C.; Tempel, D.; Pasterkamp, G.; Moll, F.L.; de Crom, R.; Krams, R. Atherosclerotic Plaque Stability Is Affected by the Chemokine CXCL10 in Both Mice and Humans. Int. J. Inflamm. 2011, 2011, 936109. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.J.; Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Heller, E.A.; Liu, E.; Tager, A.M.; Yuan, Q.; Lin, A.Y.; Ahluwalia, N.; Jones, K.; Koehn, S.L.; Lok, V.M.; Aikawa, E.; et al. Chemokine CXCL10 promotes atherogenesis by modulating the local balance of effector and reg-ulatory T cells. Circulation 2006, 113, 2301–2312. [Google Scholar] [CrossRef]
- Watanabe, Y.; Koyama, S.; Yamashita, A.; Matsuura, Y.; Nishihira, K.; Kitamura, K.; Asada, Y. Indoleamine 2,3-dioxygenase 1 in coronary atherosclerotic plaque enhances tissue factor expression in activated macrophages. Res. Pr. Thromb. Haemost. 2018, 2, 726–735. [Google Scholar] [CrossRef]
- Bekhouche, M.; Leduc, C.; Dupont, L.; Janssen, L.; Delolme, F.; Vadon-Le Goff, S.; Smargiasso, N.; Baiwir, D.; Mazzucchelli, G.; Zanella-Cleon, I.; et al. Determination of the substrate rep-ertoire of ADAMTS2, 3, and 14 significantly broadens their functions and identifies extracellular matrix organization and TGF-β signaling as primary targets. FASEB J. 2016, 30, 1741–1756. [Google Scholar] [CrossRef] [Green Version]
- Hardy, S.A.; Mabotuwana, N.S.; Murtha, L.A.; Coulter, B.; Sanchez-Bezanilla, S.; Al-Omary, M.S.; Senanayake, T.; Loering, S.; Starkey, M.; Lee, R.J.; et al. Novel role of extracellular matrix protein 1 (ECM1) in cardiac aging and myocardial infarction. PLoS ONE 2019, 14, e0212230. [Google Scholar] [CrossRef] [Green Version]
- Auclair, M.; Guénantin, A.C.; Fellahi, S.; Garcia, M.; Capeau, J. HIV antiretroviral drugs, dolutegravir, maraviroc and ri-tonavir-boosted atazanavir use different pathways to affect inflammation, senescence, and insulin sensitivity in human cor-onary endothelial cells. PLoS ONE 2020, 15, e0226924. [Google Scholar] [CrossRef]
- Patel, A. Does the Role of Angiogenesis play a Role in Atherosclerosis and Plaque Instability? Anat. Physiol. 2014, 4, 147. [Google Scholar]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Kothapalli, D.; Liu, S.L.; Bae, Y.H.; Monslow, J.; Xu, T.; Hawthorne, E.A.; Byfield, F.J.; Castagnino, P.; Rao, S.; Rader, D.J.; et al. Cardiovascular protection by ApoE and ApoE-HDL linked to sup-pression of ECM gene expression and arterial stiffening. Cell Rep. 2012, 2, 1259–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedin, L.L.; Gustafsson, J.A.; Steffensen, K.R. The oxysterol receptors LXRalpha and LXRbeta suppress proliferation in the colon. Mol. Carcinog. 2013, 52, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzu, O.F.; Noory, M.A.; Robertson, G.P. The Role of Cholesterol in Cancer. Cancer Res. 2016, 76, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.K.; Rame, J.E.; Khera, A.; Murphy, S.A.; Canham, R.M.; Peshock, R.M.; de Lemos, J.A.; Drazner, M.H. Left ventricular hypertrophy, subclinical atherosclerosis, and inflammation. Hypertension 2007, 49, 1385–1391. [Google Scholar] [CrossRef] [Green Version]
- Torii, S.; Nakayama, K.; Yamamoto, T.; Nishida, E. Regulatory Mechanisms and Function of ERK MAP Kinases. J. Biochem. 2004, 136, 557–561. [Google Scholar] [CrossRef]
- Thorburn, J.; Frost, J.; Thorburn, A. Mitogen-activated protein kinases mediate changes in gene expression, but not cytoskeletal organization associated with cardiac muscle cell hypertrophy. J. Cell Biol. 1994, 126, 1565–1572. [Google Scholar] [CrossRef]
- Zhang, G.; Ohmori, K.; Nagai, Y.; Fujisawa, Y.; Nishiyama, A.; Abe, Y.; Kimura, S. Role of AT1 receptor in isopro-terenol-induced cardiac hypertrophy and oxidative stress in mice. J. Mol. Cell. Cardiol. 2007, 42, 804–811. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, A.; Wang, X.; Sun, H. A kaempferol-3-O-β-d-glucoside, intervention effect of astragalin on estradiol metab-olism. Steroids 2019, 149, 108413. [Google Scholar] [CrossRef]
- Carrera, A.N.; Grant, M.K.; Zordoky, B.N. CYP1B1 as a therapeutic target in cardio-oncology. Clin. Sci. 2020, 134, 2897–2927. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sukhova, G.K.; Wolters, P.J.; Yang, M.; Kitamoto, S.; Libby, P.; Macfarlane, L.A.; Clair, J.M.-S.; Shi, G.-P. Mast cells promote atherosclerosis by releasing proinflammatory cytokines. Nat. Med. 2007, 13, 719–724. [Google Scholar] [CrossRef]
- Kinet, J.P. The high-affinity IgE receptor (Fc epsilon RI): From physiology to pathology. Annu. Rev. Immunol. 1999, 17, 931–972. [Google Scholar] [CrossRef]
- Yu, Y.; Blokhuis, B.R.; Garssen, J.; Redegeld, F.A. Non-IgE mediated mast cell activation. Immunol. Rev. 2018, 282, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, X.; Xiang, M.X.; Alanne-Kinnunen, M.; Wang, J.A.; Chen, H.; He, A.; Sun, X.; Lin, Y.; Tang, T.T.; et al. IgE stimulates human and mouse arterial cell apoptosis and cytokine expression and promotes atherogenesis in Apoe−/− mice. J. Clin. Investig. 2011, 121, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhu, W.; Gonzalez, F.J. Potential role of CYP1B1 in the development and treatment of metabolic diseases. Pharmacol. Ther. 2017, 178, 18–30. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | p-Value | Fold-Change |
|---|---|---|---|
| Upregulated differentially expressed genes | |||
| CXCL9, C-X-C Motif Chemokine Ligand 9 | CXCL9 | 2.08 × 10−6 | 197.457 |
| Major Histocompatibility Complex Class II DR Alpha | HLA-DRA | 1.63 × 10−5 | 137.562 |
| Tryptophan 2,3-Dioxygenase | TDO2 | 0.000551137 | 133.681 |
| Indoleamine 2,3-Dioxygenase 1 | IDO1 | 1.18 × 10−5 | 108.503 |
| C-X-C Motif Chemokine Ligand 10 | CXCL10 | 6.52 × 10−5 | 86.0006 |
| Interferon Induced Protein 44 Like | IFI44L | 0.000593426 | 70.7593 |
| Serpin Family G Member 1 | SERPING1 | 1.73 × 10−7 | 65.1139 |
| Guanylate Binding Protein 5 | GBP5 | 1.24 × 10−6 | 59.8055 |
| Downregulated differentially expressed genes | |||
| Sortilin Related Receptor 1 | SORL1 | 0.000414781 | −8.41001 |
| S100 Calcium Binding Protein A4 | S100A4 | 0.000298601 | −5.95322 |
| Sugen kinase 223 | SGK223 | 0.000583494 | −4.32646 |
| SMAD Family Member 3 | SMAD3 | 9.74 × 10−5 | −3.4329 |
| Vasohibin 1 | VASH1 | 0.000277198 | −3.30646 |
| Long Intergenic Non-Protein Coding RNA 1001 | LINC01001 | 8.77 × 10−5 | −3.21825 |
| Filamin B | FLNB | 2.46 × 10−5 | −3.1648 |
| Proteolipid Protein 2 | PLP2 | 0.000246883 | −3.0025 |
| glutathione reductase | GSR | 0.000199734 | −2.9929 |
| Neurotensin Receptor 1 | NTSR1 | 6.73 × 10−5 | −2.85795 |
| Gene Name | Gene Symbol | p-Value | Fold-Change |
|---|---|---|---|
| Upregulated differentially expressed genes | |||
| C-X-C Motif Chemokine Ligand 9 | CXCL9 | 2.35 × 10−6 | 167.766 |
| Major Histocompatibility Complex, Class II, DR Alpha | HLA-DRA | 2.06 × 10−5 | 103.719 |
| Tryptophan 2,3-Dioxygenase | AIM2/TDO2 | 2.71 × 10−6 | 66.5112 |
| Indoleamine 2,3-Dioxygenase 1 | IDO1 | 1.91 × 10−5 | 63.8001 |
| C-X-C Motif Chemokine Ligand 10 | CXCL10 | 9.54 × 10−5 | 57.1592 |
| Interferon Induced Protein 44 Like | IFI44 | 1.59 × 10−6 | 47.0317 |
| Guanylate Binding Protein 5 | GBP5 | 1.59 × 10−6 | 46.794 |
| Serpin Family G Member 1 | SERPING1 | 2.81 × 10−7 | 40.4335 |
| Purinergic Receptor P2X 7 | P2RX7 | 1.25 × 10−5 | 38.8665 |
| Guanylate Binding Protein 1 | GBP1 | 2.67 × 10−5 | 36.667 |
| Downregulated differentially expressed genes | |||
| Sortilin Related Receptor 1 | SORL1 | 0.0004207 | −8.34514 |
| S100 Calcium Binding Protein A4 | S100A4 | 0.000280278 | −6.12928 |
| Proteolipid Protein 2 | PLP2 | 4.19 × 10−5 | −5.60529 |
| NAD(P)H Quinone Dehydrogenase 1 | NQO1 | 0.000382832 | −5.44158 |
| Uronyl-2-sulfotransferase | UST | 0.000405202 | −5.24763 |
| CD37 molecule | CD37 | 3.09 × 10−5 | −5.01109 |
| xylosyltransferase I | XYLT1 | 0.000445907 | −4.90686 |
| Rho GDP dissociation inhibitor (GDI) beta | ARHGDIB | 0.000154905 | −4.55132 |
| Quiescin Q6 sulfhydryl oxidase 1 | QSOX1 | 5.81 × 10−5 | −3.92237 |
| Cytochrome b561 family, member A3 | CYB561A3 | 6.47 × 10−5 | −3.68936 |
| Ingenuity Canonical Pathways | −log (p-Value) | z-Score | Molecules |
|---|---|---|---|
| Cardiac Hypertrophy Signaling (Enhanced) | 12.19 | −1.225 | ACVR2A, ADCY8, ADCY9, AKT2, CALM1, CAMK2G, DIAPH1, DIAPH2, GNA13, HSPB1, IKBKE, IL17RA, IL1R1, IL31RA, ITGA2, ITGB3, JAK2, MKNK1, MRAS, PLCG2, PNPLA8, RCAN1, SMPDL3A, SRF, TGFBR1, TGFBR2, TNFSF10, TNFSF13B, TSC2 |
| Role of NFAT in Cardiac Hypertrophy | 1.995 | −2.111 | ADCY8, ADCY9, AKT2, CALM1, CAMK1, CAMK2G, MRAS, PLCG2, RCAN1, TGFBR1, TGFBR2 |
| Cardiac Hypertrophy Signaling | 1.893 | −1.541 | ADCY8, ADCY9, CALM1, GNA13, HSPB1, MRAS, MYL6, PLCG2, RND3, SRF, TGFBR1, TGFBR2 |
| Xenobiotic Metabolism General Signaling Pathway | 4.813 | −1.541 | |
| Role of Hypercytokinemia/hyperchemokinemia in the Pathogenesis of Influenza | 12.8 | 3.464 | CXCL10, DDX58, EIF2AK2, IFIT2, IFIT3, ISG20, MX1, OAS1, OAS3, RSAD2, STAT1, STAT2 |
| Interferon Signaling | 10.5 | 2.828 | IFI35, IFIT3, IFITM1, IRF1, MX1, OAS1, STAT1, STAT2 |
| Antigen Presentation Pathway | 10.2 | NP | B2M, CD74, HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA, TAP2 |
| Th1 and Th2 Activation Pathway | 7.04 | NP | CD274, HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA, IL4R, IRF1, mir-29, STAT1 |
| Fc Epsilon RI Signaling | 6.41 | 1.71 | AKT2, INPP5D, LCP2, LYN, MRAS, PLCG2, SYK, VAV3 |
| Pyroptosis Signaling Pathway | 6.95 | 2.828 | AIM2, GBP1, GBP2, GBP3, GBP4, GBP5, GBP7, P2RX7 |
| Role of PKR in Interferon Induction and Antiviral Response | 3.76 | 2.449 | DDX58, EIF2AK2, IFIH1, IRF1, STAT1, STAT2 |
| Activation of IRF by Cytosolic Pattern Recognition Receptors | 4.36 | 2.236 | DDX58, IFIH1, IFIT2, STAT1, STAT2 |
| Necroptosis Signaling Pathway | 3.38 | 2.236 | ATP, AXL, EIF2AK2, STAT1, STAT2, TNFSF10 |
| MSP-RON Signaling In Macrophages Pathway | 4.08 | −2.449 | HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA, STAT1 |
| Neuroinflammation Signaling Pathway | 6.14 | 1.897 | AKT2, ATP, B2M, CXCL10, HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA, P2RX7, STAT1, SYK |
| IL-4 Signaling | 5.79 | NP | AKT2, HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA, IL4R |
| B Cell Development | 5.2 | NP | HLA-DPB1, HLA-DQA1, HLA-DQA2, HLA-DQB1, HLA-DRA |
| Biological Process | Strength | p Value | Molecules |
|---|---|---|---|
| Positive regulation of cholesterol storage | 2.72 | 2.29 × 10−5 | MSR1, SCARB1, CD36 |
| Very-low-density lipoprotein particle clearance | 2.61 | 0.0021 | APOE, APOC |
| Cholesterol efflux | 2.33 | 4.75 × 10−5 | SOAT1, SCARB1, APOE, APOC1 |
| Phospholipid efflux | 2.31 | 0.0049 | APOE, APOC1 |
| Low density lipoprotein particle clearance | 2.18 | 0.00028 | SOAT1, SCARB1, CD36 |
| Reverse cholesterol transport | 2.16 | 0.0075 | SCARB1, APOE |
| Cholesterol homeostasis | 1.76 | 0.00016 | SOAT1, SCARB1, APOE, NR1H3 |
| Lipid transport | 1.52 | 8.04 × 10−8 | SOAT1, MSR1, APOE, SCARB1, APOC1, RXRA, CD36, ACSL1 |
| Regulation of hydrolase activity | 0.88 | 0.0050 | SORL1, SYK, FN1, ECM1, AKT2, SMAD3, N1RH3, APOC1 |
| Gene Function | IFN-γ | IFN-γ + Kaempferol |
|---|---|---|
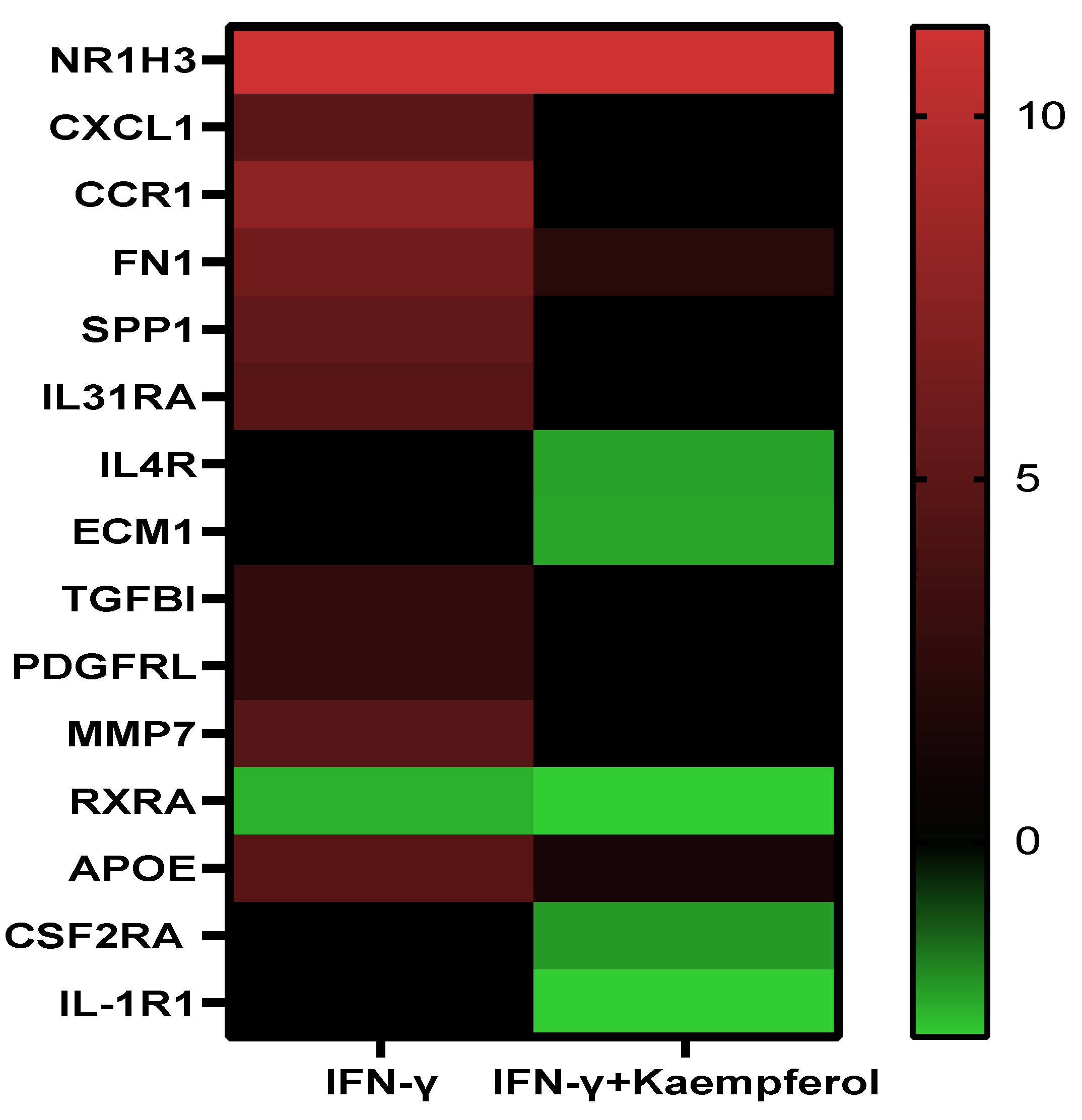
| Stress response | CCR1↑ SPP1↑ CXCl1↑ FN1↑ APOE↑ | IL4R↓ FN1↑ APOE↑ CSF2RA ↓ IL-1R1↓ |
| Apoptosis | TGFBI↑ SPP1↑ | |
| Blood coagulation and circulation | PDGFRL↑ | |
| Cell adhesion | SPP1↑ FN1↑ | FN1↑ |
| Extracellular Matrix Molecules | MMP7 ↑ FN1↑ | ECM1↓ IL4R↓ FN1↑ RXRA↓ |
| Lipid transport and metabolism | RXRA↓ NR1H3↑ | IL4R↓ NR1H3↑ RXRA↓ |
| Cell growth and proliferation | PDGFRL↑ SPP1↑ IL31RA↑ TGFBI↑ | IL4R↓ |
| Transcription regulation | RXRA↓ NR1H3↑ | RXRA↓ NR1H3↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huwait, E.; Ayoub, M.; Karim, S. Investigation of the Molecular Mechanisms Underlying the Antiatherogenic Actions of Kaempferol in Human THP-1 Macrophages. Int. J. Mol. Sci. 2022, 23, 7461. https://doi.org/10.3390/ijms23137461
Huwait E, Ayoub M, Karim S. Investigation of the Molecular Mechanisms Underlying the Antiatherogenic Actions of Kaempferol in Human THP-1 Macrophages. International Journal of Molecular Sciences. 2022; 23(13):7461. https://doi.org/10.3390/ijms23137461
Chicago/Turabian StyleHuwait, Etimad, Maha Ayoub, and Sajjad Karim. 2022. "Investigation of the Molecular Mechanisms Underlying the Antiatherogenic Actions of Kaempferol in Human THP-1 Macrophages" International Journal of Molecular Sciences 23, no. 13: 7461. https://doi.org/10.3390/ijms23137461