
Dipotassium Glycyrrhizinate on Melanoma Cell Line: Inhibition of Cerebral Metastases Formation by Targeting NF-kB Genes-Mediating MicroRNA-4443 and MicroRNA-3620—Dipotassium Glycyrrhizinate Effect on Melanoma †

 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Melanoma Cell Line and Reagents
2.2. Determination of Cellular Metabolic Activity (Cell Viability)
2.3. Measurement of the Effect of DPG on Cell Viability In Vitro
2.4. Agarose Gel Electrophoresis Analysis for DNA Fragmentation
2.5. Apoptosis by TUNEL (Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling) Assay
2.6. Wound-Healing Assay
2.7. DPG Effect on MicroRNAs (miRs)
2.8. Reverse-Transcriptase Polymerase Chain Reaction (RT-PCR) Analysis
2.9. Sphere-Cultured Stem Cell-Enriched Melanoma Populations
2.10. Statistical Tests
3. Results
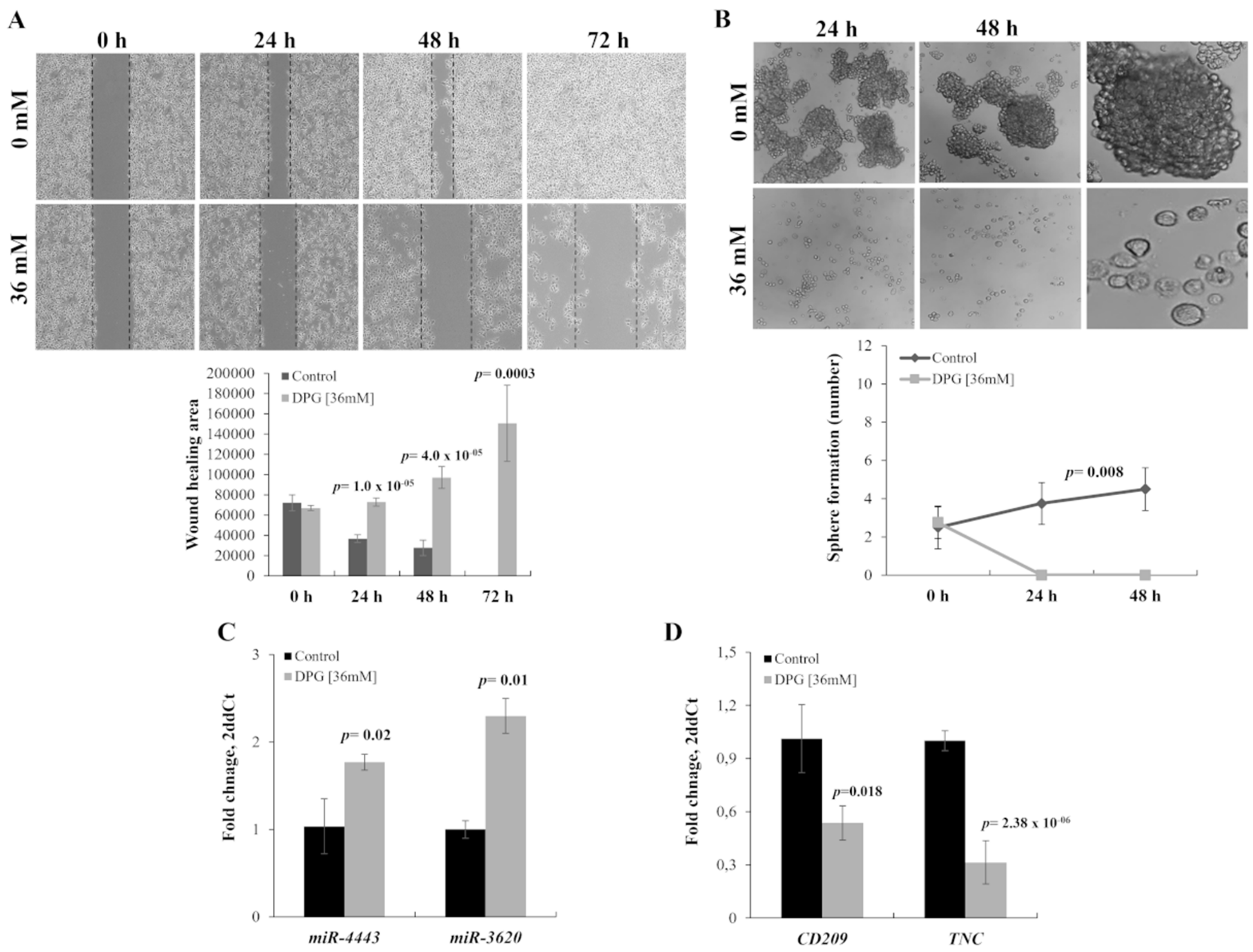
3.1. DPG Effect on Melanoma Cells Proliferation and Apoptosis
3.2. DPG Effect on Melanoma Stem-Like (SK-MEL-28) Cells
3.3. DPG Effect on miRs
3.4. TPA-Induction of MMP-9 Activation and Migration of SK-MEL-28 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AUs | arbitrary units |
| BAX | BCL2 Associated X |
| BCL-2 | BCL2 Apoptosis Regulator |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase |
| CAS | chemical abstracts service; |
| CD209 | cluster of differentiation 209; |
| DAPI | 4′,6-Diamidine-2′-phenylindole dihydrochloride |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FCS | fetal calf serum |
| DMSO | dimethyl sulfoxide |
| GA | Glycyrric acid |
| DPG | Dipotassium glycyrrhizinate |
| E2F-1 | E2F transcription factor 1 |
| ELK-1 | ETS transcription factor ELK1 |
| HaCat | keratinocyte cell line |
| IKK | kappa β kinase |
| IRAK2 | interleukin-1 receptor-associated kinase 2 |
| qPCR | quantitative polymerase chain reaction |
| MMP-9 | Matrix Metalloproteinase 9 |
| NFAT | NFAT nuclear factor |
| NF-kB | nuclear factor kappa B |
| miRs | microRNAs; MTT: (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| PARP-1 | Poly [ADP-ribose] Polymerase 1 |
| PBS | phosphate buffered saline |
| q-PCR | quantitative transcriptase polymerase chain reaction |
| RT-PCR | reverse-transcriptase polymerase chain reaction |
| SK-MEL-28 | human melanoma cell line |
| SD | standard deviation |
| TNC | Tenascin |
| TMZ | Temozolomide |
| TRAF6 | TNF receptor associated factor |
| TPA | 12-O-Tetradecanoylphorbol-13-acetate |
| TUNEL | terminal deoxynucleotidyl transferase (TdT) dUTP Nick-End labeling |
References
- Hibasami, H.; Iwase, H.; Yoshioka, K.; Takahashi, H. Glycyrrhizin induces apoptosis in human stomach cancer KATO III and human promyelocytic leukemia HL-60 cells. Int. J. Mol. Med. 2005, 16, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Hibasami, H.; Iwase, H.; Yoshioka, K.; Takahashi, H. Glycyrrhetic acid (a metabolic substance and aglycon of glycyrrhizin) induces apoptosis in human hepatoma, promyelocytic leukemia, and stomach cancer cells. Int. J. Mol. Med. 2006, 17, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivasakthivel, T.; Xu, L.; Ramaswamy, K.; Gnanasekar, M. Glycyrrhizin induces apoptosis in prostate cancer cell lines DU-145 and LNCaP. Oncol. Rep. 2008, 20, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Bonafé, G.A.; dos Santos, J.S.; Ziegler, J.V.; Umezawa, K.; Ribeiro, M.L.; Rocha, T.; Ortega, M.M. Growth inhibitory effects of dipotassium glycyrrhizinate in glioblastoma cell lines by targeting microRNAs through the NF-kB signaling pathway. Front. Cell. Neurosci. 2019, 13, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Nayak, L.; Lee, E.Q.; Wen, P.Y. Epidemiology of brain metastases. Curr. Oncol. Rep. 2012, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Egger, M.E.; McMasters, K.M.; Hao, H. Differential expression of ABCB5 in BRAF inhibitor-resistant melanoma cell lines. BMC Cancer 2018, 18, 675. [Google Scholar] [CrossRef]
- Aizer, A.A.; Lee, E.Q. Brain metastases. Neurol. Clin. 2018, 36, 557–577. [Google Scholar] [CrossRef]
- Achrol, A.S.; Rennert, R.C.; Anders, C.; Soffietti, R.; Ahluwalia, M.S.; Nayak, L.; Peters, S.; Arvold, N.D.; Harsh, G.R.; Steeg, P.S.; et al. Brain metastases. Nat. Rev. Dis. Primers 2010, 5, 5. [Google Scholar] [CrossRef]
- Vosoughi, E.; Lee, J.M.; Miller, J.R.; Nosrati, M.; Minor, D.R.; Abendroth, R.; Lee, J.W.; Andrews, B.T.; Leng, L.Z.; Wu, M.; et al. Survival and clinical outcomes of patients with melanoma brain metastasis in the era of checkpoint inhibitors and targeted therapies. BMC Cancer 2018, 18, 490. [Google Scholar] [CrossRef]
- Davies, M.A.; Liu, P.; McIntyre, S.; Kim, K.B.; Papadopoulos, N.; Hwu, W.J.; Hwu, P.; Bedikian, A. Prognostic factors for survival in melanoma patients with brain metastases. Cancer 2011, 117, 1687–1696. [Google Scholar] [CrossRef]
- Kashani-Sabet, M.; Shaikh, L.; Miller, J.R., III; Nosrati, M.; Ferreira, C.M.; Debs, R.J.; Sagebiel, R.W. NF-kappa B in the vascular progression of melanoma. J. Clin. Oncol. 2004, 22, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Kashani-Sabet, M.; Liu, Y.; Fong, S.; Desprez, P.Y.; Liu, S.; Tu, G.; Nosrati, M.; Handumrongkul, C.; Liggitt, D.; Thor, A.D.; et al. Identification of gene function and functional pathways by systemic plasmid-based ribozyme targeting in adult mice. Proc. Natl. Acad. Sci. USA 2002, 99, 3878–3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNulty, S.E.; del Rosario, R.; Cen, D.; Meyskens, F.L., Jr.; Yang, S. Comparative expression of NFkappaB proteins in melanocytes of normal skin vs. benign intradermal naevus and human metastatic melanoma biopsies. Pigment Cell Res. 2004, 17, 173–180. [Google Scholar] [CrossRef]
- Liu, J.; Suresh Kumar, K.; Yu, D.; Molton, S.A.; McMahon, M.; Herlyn, M.; Thomas-Tikhonenko, A.; Fuchs, S.Y. Oncogenic BRAF regulates β-Trcp expression and NF-kB activity in human melanoma cells. Oncogene 2007, 26, 1954–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhead, J.L.; Fallon, R.; Figueredo, H.; Langdale, J.; Malcom, A.D.B. Alternative methodology of gene diagnosis. In Human Genetic Diseases: A Practical Approach; Davies, K.E., Ed.; IRL Press Limited: Oxford, UK, 1986; pp. 51–64. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Barciszewska, A.M.; Gurda, D.; Głodowicz, P.; Nowak, S.; Naskręt-Barciszewska, M.Z. A new epigenetic mechanism of temozolomide action in glioma cells. PLoS ONE 2015, 10, e0136669. [Google Scholar] [CrossRef] [Green Version]
- Li, R.H.; Hou, X.Y.; Yang, C.S.; Liu, W.L.; Tang, J.Q.; Liu, Y.Q.; Jiang, G. Temozolomide for treating malignant melanoma. J. Coll. Physicians Surg. Pak. 2015, 25, 680–688. [Google Scholar]
- Sztiller-Sikorska, M.; Koprowska, K.; Jakubowska, J.; Zalesna, I.; Stasiak, M.; Duechler, M.; Czyz, M.E. Sphere formation and self-renewal capacity of melanoma cells is affected by the microenvironment. Melanoma Res. 2012, 22, 215–224. [Google Scholar] [CrossRef]
- Cohen-Armon, M.; Visochek, L.; Rozensal, D.; Kalal, A.; Geistrikh, I.; Klein, R.; Bendetz-Nezer, S.; Yao, Z.; Seger, R. DNA-independent PARP-1 activation by phosphorylated ERK2 increases Elk1 activity: A link to histone acetylation. Mol. Cell 2006, 25, 297–308. [Google Scholar] [CrossRef]
- Cohen-Armon, M. ARP-1 activation in the ERK signaling pathway. Trends Pharmacol. Sci. 2007, 28, 556–560. [Google Scholar] [CrossRef]
- Hassa, P.O.; Hottiger, M.O. A role of poly (ADP-ribose) polymerase in NF-kappaB transcriptional activation. Biol. Chem. 1999, 380, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Hassa, P.O.; Hottiger, M.O. The functional role of poly(ADP-ribose)polymerase 1 as novel coactivator of NF-kappa B in inflammatory disorders. Cell. Mol. Life Sci. 2002, 59, 1534–1553. [Google Scholar] [CrossRef] [PubMed]
- Olabisi, O.A.; Soto-Nieves, N.; Nieves, E.; Yang, T.T.; Yang, X.; Yu, R.Y.; Suk, H.Y.; Macian, F.; Chow, C.W. Regulation of transcription factor NFAT by ADP-ribosylation. Mol. Cell. Biol. 2008, 28, 2860–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simbulan-Rosenthal, C.M.; Rosenthal, D.S.; Luo, R.; Samara, R.; Espinoza, L.A.; Hassa, P.O.; Hottiger, M.O.; Smulson, M.E. PARP-1 binds E2F-1 independently of its DNA binding and catalytic domains, and acts as a novel coactivator of E2F-1-mediated transcription during re-entry of quiescent cells into S phase. Oncogene 2003, 22, 8460–8471. [Google Scholar] [CrossRef] [Green Version]
- Zingarelli, B.; Hake, P.W.; O’Connor, M.; Denenberg, A.; Kong, S.; Aronow, B.J. Absence of poly(ADP-ribose)polymerase-1 alters nuclear factor-kappa B activation and gene expression of apoptosis regulators after reperfusion injury. Mol. Med. 2003, 9, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Pavlick, K.P.; Laroux, F.S.; Verma, S.K.; Jordan, P.; Grisham, M.B.; Williams, L.; Alexander, J.S. Regulation and distribution of MAdCAM-1 in endothelial cells in vitro. Am. J. Physiol. Cell Physiol. 2021, 281, C1096–C1105. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.; Warren, A.; Oshima, T.; Williams, L.; Li, J.H.; Alexander, J.S. Poly ADP ribose-polymerase inhibitors prevent the upregulation of ICAM-1 and E-selectin in response to Th1 cytokine stimulation. Inflammation 2001, 25, 157–163. [Google Scholar] [CrossRef]
- Von Lukowicz, T.; Hassa, P.O.; Lohmann, C.; Boren, J.; Braunersreuther, V.; Mach, F.; Odermatt, B.; Gersbach, M.; Camici, G.G.; Stähli, B.E.; et al. PARP1 is required for adhesion molecule expression in atherogenesis. Cardiovasc. Res. 2008, 78, 158–166. [Google Scholar] [CrossRef]
- Nicolescu, A.C.; Holt, A.; Kandasamy, A.D.; Pacher, P.; Schulz, R. Inhibition of matrix metalloproteinase-2 by PARP inhibitors. Biochem. Biophys. Res. Commun. 2009, 387, 646–650. [Google Scholar] [CrossRef] [Green Version]
- Geijtenbeek, T.B.H.; Engering, A.; Van Kooyk, Y. DC-SIGN, a C-type lectin on dendritic cells that unveils many aspects of dendritic cell biology. J. Leuk. Biol. 2002, 71, 921–931. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.H.; Torensma, R.; Van Vliet, S.J.; van Duijnhoven, G.C.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Kirkwood, J.M.; Wells, A. Tenascin-C Signaling in melanoma. Cell Adhes. Migr. 2015, 9, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Chao, C.Y.; Lin, L.L.; Lu, C.Y.; Liu, K.L.; Lii, C.K.; Li, C.C. Inhibition of matrix metalloproteinase-9 expression by docosahexaenoic acid mediated by heme oxygenase 1 in 12-O-tetradecanoylphorbol-13-acetate-induced MCF-7 human breast cancer cells. Arch. Toxicol. 2013, 87, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.T.; Lee, T.H.; Cheng, C.H.; Chen, K.C.; Chen, Y.C.; Lin, C.W. Antroquinonol from Antrodia Camphorata suppresses breast tumor migration/invasion through inhibiting ERK-AP-1- and AKT-NF-kB-dependent MMP-9 and epithelial-mesenchymal transition expressions. Food Chem. Toxicol. 2015, 78, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lii, C.K.; Chang, J.W.; Chen, J.J.; Chen, H.W.; Liu, K.L.; Yeh, S.L.; Wang, T.S.; Liu, S.H.; Tsai, C.H.; Li, C.C. Docosahexaenoic acid inhibits 12-O-tetradecanoylphorbol-13- acetate-induced fascin-1-dependent breast cancer cell migration by suppressing the PKCδ- and Wnt-1/β-catenin-mediated pathways. Oncotarget 2016, 7, 25162–25179. [Google Scholar] [CrossRef] [Green Version]
- Cheng-Wei, L.; Shing-Chuan, S.; Chih-Chiang, C.; Liang-Yo, Y.; Lin-Ting, S.; Yen-Chou, C. 12-O-tetradecanoylphorbol-13-acetate-induced invasion/migration of glioblastoma cells through activating PKCalpha/ERK/NF-kappaB-dependent MMP-9 expression. J. Cell Physiol. 2010, 225, 472–481. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonafé, G.A.; dos Santos, J.S.; Ziegler, J.V.; Marson, F.A.L.; Rocha, T.; Ortega, M.M. Dipotassium Glycyrrhizinate on Melanoma Cell Line: Inhibition of Cerebral Metastases Formation by Targeting NF-kB Genes-Mediating MicroRNA-4443 and MicroRNA-3620—Dipotassium Glycyrrhizinate Effect on Melanoma. Int. J. Mol. Sci. 2022, 23, 7251. https://doi.org/10.3390/ijms23137251
Bonafé GA, dos Santos JS, Ziegler JV, Marson FAL, Rocha T, Ortega MM. Dipotassium Glycyrrhizinate on Melanoma Cell Line: Inhibition of Cerebral Metastases Formation by Targeting NF-kB Genes-Mediating MicroRNA-4443 and MicroRNA-3620—Dipotassium Glycyrrhizinate Effect on Melanoma. International Journal of Molecular Sciences. 2022; 23(13):7251. https://doi.org/10.3390/ijms23137251
Chicago/Turabian StyleBonafé, Gabriel Alves, Jéssica Silva dos Santos, Jussara Vaz Ziegler, Fernando Augusto Lima Marson, Thalita Rocha, and Manoela Marques Ortega. 2022. "Dipotassium Glycyrrhizinate on Melanoma Cell Line: Inhibition of Cerebral Metastases Formation by Targeting NF-kB Genes-Mediating MicroRNA-4443 and MicroRNA-3620—Dipotassium Glycyrrhizinate Effect on Melanoma" International Journal of Molecular Sciences 23, no. 13: 7251. https://doi.org/10.3390/ijms23137251