
Plant Responses to Herbivory, Wounding, and Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Plant Defense Prior to Herbivore Attack
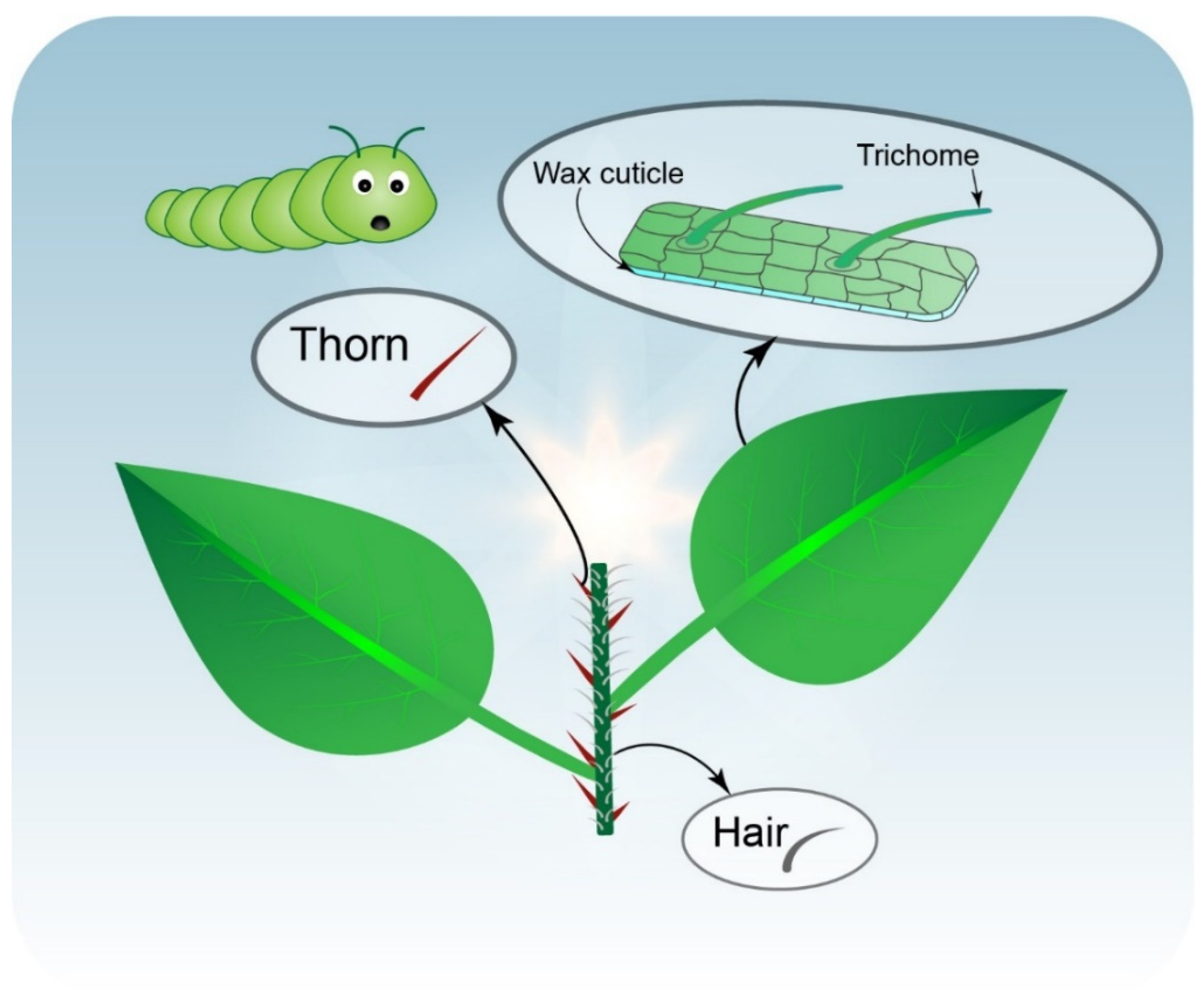
Plant Physical Barriers
3. Plant Responses during Herbivore Attack, Wounding, and Infection
3.1. Signal Transduction in Host Plants
3.2. Intracellular Signaling
3.3. Airborne Signaling
4. Plant Response to Biotic Attack through Epigenetic Regulation and SMs
4.1. Epigenetic Regulation
4.2. SMs
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Full Name |
| AOS | Allene oxide synthase |
| AW | Armyworm |
| BPH | Brown planthopper |
| BABA | β-aminobutyric acid |
| CaMs | Calmodulins |
| CMLs | Calmodulin-like proteins |
| CCaMKs | Ca2+/CaM-dependent protein kinases |
| CDPKs/CPKs | Calcium-dependent protein kinases |
| CBL | Calcineurin B-like protein |
| CIPK | CBL interacting protein kinase |
| OPDA | Cis-12-oxo-phytodienoic acid |
| [Ca2+] cyt | Cytosolic calcium |
| DAMPs | Damage-associated molecular patterns |
| ETI | Effector-triggered immunity |
| ELP2 | ELONGATOR PROTEIN2 |
| GLRs | Glutamate receptor-like channels |
| GABA | γ-aminobutyric acid |
| HAMPs | Herbivore-associated molecular patterns |
| HATs | Histone acetyltransferases |
| HDACs | Histone deacetylases |
| HUB1 | Histone monoubiquitination1 |
| H2O2 | Hydrogen peroxide |
| H2Bub | H2B monoubiquitination |
| JA | Jasmonic acid |
| JA-Ile | Jasmonoyl-L-isoleucine |
| JJW | JAV1–JAZ8–WRKY51 |
| LOX | Lipoxygenase |
| LncRNAs | Long non-coding RNAs |
| MeJA | Methyl jasmonate |
| miRNAs | MicroRNAs |
| MAMPs | Microbe-associated molecular patterns |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| OS | Oral secretions |
| ACAs | P-IIB autoinhibited Ca2+-ATPases |
| ECAs | P-IIA ER-type Ca2+-ATPases |
| PRRs | Pattern recognition receptors |
| PTI | Pattern-triggered immunity |
| PAE9 | Pectin acetylesterase 9 |
| Vm | Plasma membrane potential |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| SA | Salicylic acid |
| SMs | Secondary metabolites |
| SDG | Set domain group |
| siRNAs | Short interfering RNAs |
| SAR | Systemic acquired resistance |
| VOCs | Volatile organic compounds |
References
- Belete, T. Defense mechanisms of plants to insect pests: From morphological to biochemical approach. Trends Tech. Sci. Res. 2018, 2, 555584. [Google Scholar] [CrossRef] [Green Version]
- Kloth, K.J.; Abreu, I.N.; Delhomme, N.; Petrík, I.; Villard, C.; Ström, C.; Amini, F.; Novák, O.; Moritz, T.; Albrectsen, B.R. PECTIN AceTyLeSTerase9 affects the transcriptome and metabolome and delays aphid feeding. Plant Physiol. 2019, 181, 1704–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Zhang, Y. Short- and long-distance signaling in plant defense. Plant J. 2021, 105, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Zebelo, S.A.; Maffei, M.E. Role of early signalling events in plant-insect interactions. J. Exp. Bot. 2015, 66, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares-Silva, M.; Diniz, F.F.; Gomes, G.N.; Bahia, D. The mitogen-activated protein kinase (MAPK) pathway: Role in immune evasion by trypanosomatids. Front. Microbiol. 2016, 7, 183. [Google Scholar] [CrossRef]
- Turgut-Kara, N.; Arikan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148, 47–54. [Google Scholar] [CrossRef]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and mechanism of jasmonic acid in plant responses to abiotic and biotic stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef]
- Shah, J.M.; Joseph, J.T. Involvement of epigenetic regulation in plant defence during biotic stress. In Plant Defense Mechanisms; Kimatu, J.N., Ed.; IntechOpen: London, UK, 2022. [Google Scholar]
- Ashapkin, V.V.; Kutueva, L.I.; Aleksandrushkina, N.I.; Vanyushin, B.F. Epigenetic mechanisms of plant adaptation to biotic and abiotic stresses. Int. J. Mol. Sci. 2020, 21, 7457. [Google Scholar] [CrossRef]
- Joseph, J.T.; Shah, J.M. Biotic stress-induced epigenetic changes and transgenerational memory in plants. Biologia 2022, 1–15. [Google Scholar] [CrossRef]
- Lewandowska, M.; Keyl, A. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kong, L.; Zhi, P.; Chang, C. Update on cuticular wax biosynthesis and its roles in plant disease resistance. Int. J. Mol. Sci. 2020, 21, 5514. [Google Scholar] [CrossRef] [PubMed]
- Markstädter, C.; Federle, W.; Jetter, R.; Riederer, M.; Hölldobler, B. Chemical composition of the slippery epicuticular wax blooms on macaranga (Euphorbiaceae) ant-plants. Chemoecology 2000, 10, 33–40. [Google Scholar] [CrossRef]
- Ali Azam, M.; Khan, M.A.U.; Rao, A.Q.; Iqbal, A.; ud Din, S.; Shahid, A.A. Biochemical evidence of epicuticular wax compounds involved in cotton-whitefly interaction. PLoS ONE 2021, 16, e0250902. [Google Scholar] [CrossRef]
- Kariyat, R.R.; Hardison, S.B.; De Moraes, C.M.; Mescher, M.C. Plant spines deter herbivory by restricting caterpillar movement. Biol. Lett. 2017, 13, 20170176. [Google Scholar] [CrossRef]
- Singh, A.; Dilkes, B.; Sela, H.; Tzin, V. The effectiveness of physical and chemical defense responses of wild emmer wheat against aphids depends on leaf position and genotype. Front. Plant Sci. 2021, 12, 667820. [Google Scholar] [CrossRef]
- De Resende, J.T.V.; Filho, R.B.D.L.; Ribeiro, L.K.; Corrêa, J.V.W.; Maciel, C.D.D.G.; Youssef, K. Strawberry genotypes with resistance to Tetranychus urticae mediated by leaf trichomes. Ciênc. Agrotec. 2020, 44, e006920. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, F.E.; Peiffer, M.; Ray, S.; Tan, C.; Felton, G.W. Silicon-mediated enhancement of herbivore resistance in agricultural crops. Front. Plant Sci. 2021, 12, 631824. [Google Scholar] [CrossRef]
- Kaur, J.; Kariyat, R. Role of trichomes in plant stress biology. In Evolutionary Ecology of Plant-Herbivore Interaction; Núñez-Farfán, J., Valverde, P., Eds.; Springer: Cham, Switzerland, 2020; pp. 15–35. [Google Scholar]
- De Bobadilla, M.F.; Van Wiechen, R.; Gort, G.; Poelman, E.H. Plasticity in induced resistance to sequential attack by multiple herbivores in Brassica nigra. Oecologia 2022, 198, 11–20. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Bao, Q.; Wang, H.; Hou, S. Plant elicitor peptide 1 fortifies root cell walls and triggers a systemic root-to-shoot immune signaling in Arabidopsis. Plant Signal. Behav. 2022, 17, e2034270. [Google Scholar] [CrossRef]
- Gandhi, A.; Kariyat, R.R.; Chappa, C.; Tayal, M.; Sahoo, N. Tobacco hornworm (Manduca sexta) oral secretion elicits reactive oxygen species in isolated tomato protoplasts. Int. J. Mol. Sci. 2020, 21, 8297. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P. Receptor kinases in plant responses to herbivory. Curr. Opin. Biotechnol. 2021, 70, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Liu, H.; Sun, Y.; Xu, Z.; Liang, S.; Li, B.; Ding, X.; Li, J.; Wang, Q.; Sun, L.; et al. Transcriptome and metabolome analysis reveal that oral secretions from Helicoverpa armigera and Spodoptera litura influence wound-induced host response in cotton. Crop J. 2020, 8, 929–942. [Google Scholar] [CrossRef]
- Gandhi, A.; Kariyat, R.; Harikishore, A.; Ayati, M.; Bhunia, A.; Sahoo, N. Deciphering the role of ion channels in early defense signaling against herbivorous insects. Cells 2021, 10, 2219. [Google Scholar] [CrossRef]
- Camoni, L.; Barbero, F.; Aducci, P.; Maffei, M.E. Spodoptera littoralis oral secretions inhibit the activity of Phaseolus lunatus plasma membrane H+-ATPase. PLoS ONE 2018, 13, e0202142. [Google Scholar] [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef]
- Ghosh, S.; Bheri, M.; Bisht, D.; Pandey, G.K. Calcium signaling and transport machinery: Potential for development of stress tolerance in plants. Curr. Plant Biol. 2022, 29, 100235. [Google Scholar] [CrossRef]
- Noman, M.; Aysha, J.; Ketehouli, T.; Yang, J.; Du, L.; Wang, F.; Li, H. Calmodulin binding transcription activators: An interplay between calcium signalling and plant stress tolerance. J. Plant Physiol. 2021, 256, 153327. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 655. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tsuda, K.; Sato, M.; Cohen, J.D.; Katagiri, F.; Glazebrook, J. Arabidopsis CaM binding protein CBP60g contributes to MAMP-induced SA accumulation and is involved in disease resistance against Pseudomonas syringae. PLoS Pathog. 2009, 5, e1000301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Dunand, C.; Snedden, W.; Galaud, J.P. CaM and CML emergence in the green lineage. Trends Plant Sci. 2015, 20, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Heo, W.D.; Lee, S.H.; Kim, M.C.; Kim, J.C.; Chung, W.S.; Chun, H.J.; Lee, K.J.; Park, C.Y.; Park, H.C.; Choi, J.Y.; et al. Involvement of specific calmodulin isoforms in salicylic acid-independent activation of plant disease resistance responses. Proc. Natl. Acad. Sci. USA 1999, 96, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Kanchiswamy, C.N.; Maffei, M.E. Calcium signaling preceding the emission of plant volatiles in plant–insect interactions. J. Indian Inst. Sci. 2015, 95, 15–24. [Google Scholar]
- Vadassery, J.; Reichelt, M.; Hause, B.; Gershenzon, J.; Boland, W.; Mithöfer, A. CML42-mediated calcium signaling coordinates responses to spodoptera herbivory and abiotic stresses in Arabidopsis. Plant Physiol. 2012, 159, 1159–1175. [Google Scholar] [CrossRef] [Green Version]
- Scholz, S.S.; Vadassery, J.; Heyer, M.; Reichelt, M.; Bender, K.W.; Snedden, W.A.; Boland, W.; Mithöfer, A. Mutation of the Arabidopsis calmodulin-like protein CML37 deregulates the jasmonate pathway and enhances susceptibility to herbivory. Mol. Plant 2014, 7, 1712–1726. [Google Scholar] [CrossRef] [Green Version]
- Vadassery, J.; Scholz, S.S.; Mithöfer, A. Multiple calmodulin-like proteins in Arabidopsis are induced by insect-derived (Spodoptera littoralis) oral secretion. Plant Signal. Behav. 2012, 7, 1277–1280. [Google Scholar] [CrossRef] [Green Version]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2 + sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Ramachandiran, S.; Takezawa, D.; Wang, W.; Poovaiah, B.W.; Aeronautics, N. Functional domains of plant chimeric calcium/ calmodulin-dependent protein kinase: Regulation by autoinhibitory and visinin-like. J. Biochem. 1997, 121, 984–990. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; He, Q.; Chai, M.; Huang, Y.; Chen, F.; Wang, X.; Liu, Y.; Cai, H.; Qin, Y. Genome-wide investigation of calcium-dependent protein kinase gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2020, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Atif, R.M.; Shahid, L.; Waqas, M.; Ali, B.; Rashid, M.A.R.; Azeem, F.; Nawaz, M.A.; Wani, S.H.; Chung, G. Insights on calcium-dependent protein kinases (CPKs) signaling for abiotic stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 5289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardar, A.; Nandi, A.K.; Chattopadhyay, D. CBL-interacting protein kinase 6 negatively regulates immune response to Pseudomonas syringae in Arabidopsis. J. Exp. Bot. 2017, 68, 3573–3584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; He, X.; Hu, W.; Liu, G.; Wang, P.; He, C.; Shi, H.; Calmodulin, C. Functional analysis of MeCIPK23 and MeCBL1/ 9 in cassava defense response against Xanthomonas axonopodis Pv. manihotis. Plant Cell Rep. 2018, 37, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, Q.; Yu, Y.; Qiao, Y.; Haq, S.; Gong, Z. The CBL–CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Wang, X.; Hao, L.; Zhu, B.; Jiang, Z. Plant calcium signaling in response to potassium deficiency. Int. J. Mol. Sci. 2018, 19, 3456. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Zhang, T.; Koo, A.J.; Stacey, G.; Tanaka, K. Extracellular ATP acts on jasmonate signaling to reinforce plant defense. Plant Physiol. 2018, 176, 511–523. [Google Scholar] [CrossRef] [Green Version]
- García Bossi, J.; Kumar, K.; Barberini, M.L.; Domínguez, G.D.; Rondón Guerrero, Y.D.C.; Marino-Buslje, C.; Obertello, M.; Muschietti, J.P.; Estevez, J.M. The role of P-Type IIA and P-Type IIB Ca2+-ATPases in plant development and growth. J. Exp. Bot. 2020, 71, 1239–1248. [Google Scholar] [CrossRef]
- Poór, P. Effects of salicylic acid on the metabolism of mitochondrial reactive oxygen species in plants. Biomolecules 2020, 10, 341. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, N.; Shan, J.; Peng, Y.; Guo, J.; Zhou, C.; Shi, S.; Zheng, X.; Wu, D.; Guan, W.; et al. Salivary protein 1 of brown planthopper is required for survival and induces immunity response in plants. Front. Plant Sci. 2020, 11, 571280. [Google Scholar] [CrossRef]
- Herrera-Vásquez, A.; Salinas, P.; Holuigue, L. Salicylic acid and reactive oxygen species interplay in the transcriptional control of defense genes expression. Front. Plant Sci. 2015, 6, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, A.; Christensen, S.A.; Hunter, C.T.; Alborn, H.T. Herbivore-derived fatty-acid amides elicit reactive oxygen species burst in plants. J. Exp. Bot. 2018, 69, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Turkan, I. ROS and RNS: Key signalling molecules in plants. J. Exp. Bot. 2018, 69, 3313–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, V.C.; Drzewiecka, K.; Jeleń, H.; Narozna, D.; Rucińska-Sobkowiak, R.; Kesy, J.; Floryszak-Wieczorek, J.; Gabryś, B.; Morkunas, I. Differential induction of Pisum sativum defense signaling molecules in response to pea aphid infestation. Plant Sci. 2014, 221–222, 1–12. [Google Scholar] [CrossRef]
- Woźniak, A.; Formela, M.; Bilman, P.; Grześkiewicz, K.; Bednarski, W.; Marczak, Ł.; Narożna, D.; Dancewicz, K.; Mai, V.C.; Borowiak-Sobkowiak, B.; et al. The dynamics of the defense strategy of pea induced by exogenous nitric oxide in response to aphid infestation. Int. J. Mol. Sci. 2017, 18, 329. [Google Scholar] [CrossRef] [Green Version]
- Molassiotis, A.; Job, D.; Ziogas, V.; Tanou, G. Citrus plants: A model system for unlocking the secrets of NO and ROS-inspired priming against salinity and drought. Front. Plant Sci. 2016, 7, 229. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Fan, M.; Yang, M.; Zhao, J.; Zhang, W.; Su, Y.; Xiao, L.; Deng, H.; Xie, D. Injury activates Ca2+/calmodulin-dependent phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis. Mol. Cell 2018, 70, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Grenzi, M.; Bonza, M.C.; Alfieri, A.; Costa, A. Structural insights into long-distance signal transduction pathways mediated by plant glutamate receptor-like channels. New Phytol. 2021, 229, 1261–1267. [Google Scholar] [CrossRef]
- Thorpe, M.R.; Ferrieri, A.P.; Herth, M.M.; Ferrieri, R.A. 11C-Imaging: Methyl jasmonate moves in both phloem and xylem, promotes transport of jasmonate, and of photoassimilate even after proton transport is decoupled. Planta 2007, 226, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Farmer, E.E.; Gasperini, D.; Acosta, I.F. The squeeze cell hypothesis for the activation of jasmonate synthesis in response to wounding. J. Physiol. 2014, 204, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, H.; Lin, J. Systemin-mediated long-distance systemic defense responses. New Phytol. 2020, 226, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, I.; Hause, B.; Maucher, H.; Pitzschke, A.; Miersch, O.; Ziegler, J.; Ryan, C.A.; Wasternack, C. Allene oxide cyclase dependence of the wound response and vascular bundle-specific generation of jasmonates in tomato—Amplification in wound signalling. Plant J. 2003, 33, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, A.; Caldelari, D.; Wolfender, J.L.; Farmer, E.E. Four 13-lipoxygenases contribute to rapid jasmonate synthesis in wounded Arabidopsis thaliana leaves: A role for lipoxygenase 6 in responses to long-distance wound signals. New Phytol. 2013, 197, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 Review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Schulze, A.; Zimmer, M.; Mielke, S.; Stellmach, H.; Melnyk, C.W.; Hause, B.; Gasperini, D. Wound-induced shoot-to-root relocation of JA-Ile precursors coordinates Arabidopsis growth. Mol. Plant 2019, 12, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Sato, C.; Aikawa, K.; Sugiyama, S.; Nabeta, K.; Masuta, C.; Matsuura, H. Distal transport of exogenously applied jasmonoyl-isoleucine with wounding stress. Plant Cell Physiol. 2011, 52, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.; Takeishi, S.; Kiatoka, N.; Sato, C.; Sueda, K. Phytochemistry transportation of de novo synthesized jasmonoyl isoleucine in tomato. Phytochemistry 2012, 83, 25–33. [Google Scholar] [CrossRef]
- Glauser, G.; Grata, E.; Dubugnon, L.; Rudaz, S.; Farmer, E.E.; Wolfender, J.L. Spatial and temporal dynamics of jasmonate synthesis and accumulation in Arabidopsis in response to wounding. J. Biol. Chem. 2008, 283, 16400–16407. [Google Scholar] [CrossRef] [Green Version]
- Vos, I.A.; Verhage, A.; Schuurink, R.C.; Watt, L.G.; Pieterse, C.M.J.; Van Wees, S.C.M. Onset of herbivore-induced resistance in systemic tissue primed for jasmonate-dependent defenses is activated by abscisic acid. Front. Plant Sci. 2013, 4, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyer, M.; Reichelt, M.; Mithöfer, A. A holistic approach to analyze systemic jasmonate accumulation in individual leaves of Arabidopsis rosettes upon wounding. Front. Plant Sci. 2018, 9, 1569. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral scents and fruit aromas: Functions, compositions, biosynthesis, and regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef] [PubMed]
- Duran-flores, D.; Heil, M. Sources of specificity in plant damaged-self recognition. Curr. Opin. Plant Biol. 2016, 32, 77–87. [Google Scholar] [CrossRef]
- Meents, A.K.; Mithöfer, A. Plant–plant communication: Is there a role for volatile damage-associated molecular patterns? Front. Plant Sci. 2020, 11, 583275. [Google Scholar] [CrossRef]
- Chen, Y.C.; Siems, W.F.; Pearce, G.; Ryan, C.A. Six peptide wound signals derived from a single precursor protein in Ipomoea batatas leaves activate the expression of the defense gene sporamin. J. Biol. Chem. 2008, 283, 11469–11476. [Google Scholar] [CrossRef] [Green Version]
- Arce, C.M.; Besomi, G.; Glauser, G.; Turlings, T.C.J. Caterpillar-induced volatile emissions in cotton: The relative importance of damage and insect-derived factors. Front. Plant Sci. 2021, 12, 709858. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Schultz, J.C. Rapid changes in tree leaf chemistry induced by damage: Evidence for communication between plants. Science 1983, 221, 277–279. [Google Scholar] [CrossRef]
- Choi, D.; Bostock, R.M.; Avdiushko, S.; Hildebrand, D.F. Lipid-derived signals that discriminate wound- and pathogen-responsive isoprenoid pathways in plants: Methyl jasmonate and the fungal elicitor arachidonic acid induce different 3-hydroxy-3-methylglutaryl-coenzyme a reductase genes and antimicrobial isopre. Proc. Natl. Acad. Sci. USA 1994, 91, 2329–2333. [Google Scholar] [CrossRef] [Green Version]
- Zeringue, H.J. Changes in cotton leaf chemistry induced by volatile elicitors. Phytochemistry 1987, 26, 1357–1360. [Google Scholar] [CrossRef]
- Karban, R.; Maron, J.; Felton, G.W.; Ervin, G.; Eichenseer, H. Herbivore damage to sagebrush induces resistance in wild tobacco: Evidence for eavesdropping between plants. Oikos 2003, 100, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Farmer, E.E.; Ryan, C.A. Interplant communication: Airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713–7716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, M.S. Ethylene in plant growth, development, and senescence. In Plant Hormones; Davies, P.J., Ed.; Springer: Berlin, Germany, 1995; pp. 486–508. [Google Scholar]
- Piesik, D.; Wenda-Piesik, A. Sitophilus granarius responses to blends of five groups of cereal kernels and one group of plant volatiles. J. Stored Prod. Res. 2015, 63, 63–66. [Google Scholar] [CrossRef]
- Wenda-Piesik, A.; Piesik, D.; Nowak, A.; Wawrzyniak, M. Tribolium confusum responses to blends of cereal kernels and plant volatiles. J. Appl. Entomol. 2016, 140, 558–563. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Wang, X.; Chang, C. Insight into the role of epigenetic processes in abiotic and biotic stress response in wheat and barley. Int. J. Mol. Sci. 2020, 21, 1480. [Google Scholar] [CrossRef] [Green Version]
- Kellenberger, R.T.; Schlüter, P.M.; Schiestl, F.P. Herbivore-induced DNA demethylation changes floral signalling and attractiveness to pollinators in Brassica rapa. PLoS ONE 2016, 11, e0166646. [Google Scholar] [CrossRef] [Green Version]
- Rambani, A.; Pantalone, V.; Yang, S.; Rice, J.H.; Song, Q.; Mazarei, M.; Arelli, P.R.; Meksem, K.; Stewart, C.N.; Hewezi, T. Identification of introduced and stably inherited DNA methylation variants in soybean associated with soybean cyst nematode parasitism. New Phytol. 2020, 227, 168–184. [Google Scholar] [CrossRef]
- Geng, S.; Kong, X.; Song, G.; Jia, M.; Guan, J.; Wang, F.; Qin, Z.; Wu, L.; Lan, X.; Li, A.; et al. DNA Methylation dynamics during the interaction of wheat progenitor Aegilops tauschii with the obligate biotrophic fungus Blumeria graminis F. Sp. tritici. New Phytol. 2019, 221, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Jeyaraj, A.; Elango, T.; Li, X.; Guo, G. Utilization of microRNAs and their regulatory functions for improving biotic stress tolerance in tea plant [Camellia sinensis (L.) O. Kuntze]. RNA Biol. 2020, 17, 1365–1382. [Google Scholar] [CrossRef]
- Prabu, G.R.; Mandal, A.K.A. Computational identification of miRNAs and their target genes from expressed sequence tags of tea (Camellia sinensis). Genom. Proteom. Bioinform. 2010, 8, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Brant, E.J.; Budak, H. Plant small non-coding RNAs and their roles in biotic stresses. Front. Plant Sci. 2018, 9, 1038. [Google Scholar] [CrossRef] [Green Version]
- Jeyaraj, A.; Liu, S.; Zhang, X.; Zhang, R.; Shangguan, M.; Wei, C. Genome-wide identification of microRNAs responsive to Ectropis oblique feeding in tea plant (Camellia sinensis L.). Sci. Rep. 2017, 7, 13634. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Lv, W.; Hu, L.; Rao, W.; Zeng, Y.; Zhu, L.; He, Y. Identification and analysis of brown planthopper-responsive microRNAs in resistant and susceptible rice plants. Sci. Rep. 2017, 7, 8712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.; Lin, J.; Li, Y.; Jhu, M.; King, Y.; Jeng, S. MicroR408 regulates defense response upon wounding in sweet potato. J. Exp. Bot. 2019, 70, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zheng, H.; Sui, N. Regulation mechanism of long non-coding rna in plant response to stress. Biochem. Biophys. Res. Commun. 2018, 503, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jin, J.; Xu, J.; Wang, L.; Li, J.; Lou, Y.; Baldwin, I.T. Long non-coding RNAs associate with jasmonate-mediated plant defence against herbivores. Plant Cell Environ. 2021, 44, 982–994. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, S.; Jin, J.; Li, R. Identification of herbivore-elicited long non-coding RNAs in rice. Plant Signal. Behav. 2021, 16, 1916702. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Jin, J.J.; Li, L.H.; Qu, S.H. Long non-coding RNAs responsive to blast fungus infection in rice. Rice 2020, 13, 77. [Google Scholar] [CrossRef]
- Wang, G.; Wang, X.; Zhang, Y.; Yang, J.; Li, Z.; Wu, L.; Wu, J.; Wu, N.; Liu, L.; Liu, Z.; et al. Dynamic characteristics and functional analysis provide new insights into long non-coding RNA responsive to Verticillium dahliae infection in Gossypium hirsutum. BMC Plant Biol. 2021, 21, 68. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Z.; Feng, P.; Zhong, X.; Ma, Q.; Su, Q.; Wang, X.; Li, C.; Yang, Y. Identification and the potential roles of long non-coding RNAs in cotton leaves damaged by Aphis gossypii. Plant Growth Regul. 2019, 88, 215–225. [Google Scholar] [CrossRef]
- Zhi, P.; Chang, C. Exploiting epigenetic variations for crop disease resistance improvement. Front. Plant Sci. 2021, 12, 692328. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Luo, M.; Zhang, W.; Zhao, J.; Zhang, J.; Wu, K.; Tian, L.; Duan, J. Histone acetyltransferases in rice (Oryza sativa L.): Phylogenetic analysis, subcellular localization and expression. BMC Plant Biol. 2012, 12, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Prado, J.S.; Piquerez, S.J.M.; Bendahmane, A.; Hirt, H.; Raynaud, C.; Benhamed, M. Modify the histone to win the battle: Chromatin dynamics in plant–pathogen interactions. Front. Plant Sci. 2018, 9, 355. [Google Scholar] [CrossRef]
- Ramos-Cruz, D.; Troyee, A.N.; Becker, C. Epigenetics in plant organismic interactions. Curr. Opin. Plant Biol. 2021, 61, 102060. [Google Scholar] [CrossRef]
- Latrasse, D.; Jégu, T.; Li, H.; de Zelicourt, A.; Raynaud, C.; Legras, S.; Gust, A.; Samajova, O.; Veluchamy, A.; Rayapuram, N.; et al. MAPK-triggered chromatin reprogramming by histone deacetylase in plant innate immunity. Genome Biol. 2017, 18, 131. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Yang, L.; Chen, X.; Wang, Z.; Sun, Q.; Hong, A.; Zhang, A.; Zhong, X.; Hua, J. HOS15 and HDA9 negatively regulate immunity through histone deacetylation of intracellular immune receptor NLR genes in Arabidopsis. New Phytol. 2020, 226, 507–522. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Q.; Wu, Z.; Wang, H.; Han, S.; Jin, Y.; Zhou, J.; Zhang, Z.; Jiang, J.; Shen, Y.; et al. HISTONE DEACETYLASE 6 represses pathogen defence responses in Arabidopsis thaliana. Plant Cell Environ. 2017, 40, 2972–2986. [Google Scholar] [CrossRef]
- Kong, L.; Zhi, P.; Liu, J.; Li, H.; Zhang, X.; Xu, J.; Zhou, J.; Wang, X.; Chang, C. Epigenetic activation of enoyl-coa reductase by an acetyltransferase complex triggers wheat wax biosynthesis. Plant Physiol. 2020, 183, 1250–1267. [Google Scholar] [CrossRef]
- Li, W.; Xiong, Y.; Lai, L.B.; Zhang, K.; Li, Z.; Kang, H.; Dai, L.; Gopalan, V.; Wang, G.L.; Liu, W. The rice RNase P protein subunit Rpp30 confers broad-spectrum resistance to fungal and bacterial pathogens. Plant Biotechnol. J. 2021, 19, 1988–1999. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Su, H.; Li, Z.; Mai, P.; Sun, B.; Li, Z.; Heng, Z.; Xu, X.; Yang, S.; Li, T. Involvement of histone acetylation in tomato resistance to Ralstonia solanacearum. Sci. Hortic. 2021, 285, 110163. [Google Scholar] [CrossRef]
- Wang, C.; Wang, C.; Zou, J.; Yang, Y.; Li, Z.; Zhu, S. Epigenetics in the plant–virus interaction. Plant Cell Rep. 2019, 38, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Zimmerli, L. The histone demethylase IBM1 positively regulates Arabidopsis immunity by control of defense gene expression. Front. Plant Sci. 2019, 10, 1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ménard, R.; Li, Y.; Coruzzi, G.M.; Heitz, T.; Shen, W.H.; Berr, A. Arabidopsis SDG8 potentiates the sustainable transcriptional induction of the pathogenesis-related genes PR1 and PR2 during plant defense response. Front. Plant Sci. 2020, 11, 277. [Google Scholar] [CrossRef] [Green Version]
- Dóczi, R.; Brader, G.; Pettkó-Szandtner, A.; Rajh, I.; Djamei, A.; Pitzschke, A.; Teige, M.; Hirt, H. The Arabidopsis mitogen-activated protein kinase kinase MKK3 is upstream of group c mitogen-activated protein kinases and participates in pathogen signaling. Plant Cell 2007, 19, 3266–3279. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.W.; Roux, M.; Petersen, M.; Mundy, J. MAP kinase cascades in Arabidopsis innate immunity. Front. Plant Sci. 2012, 3, 169. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Chen, X.; Zhong, X.; Zhao, Y.; Liu, X.; Zhou, S.; Cheng, S.; Zhou, D.X. Jumonji C domain protein JMJ705-mediated removal of histone H3 Lysine 27 trimethylation is involved in defense-related gene activation in rice. Plant Cell 2013, 25, 4725–4736. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Pei, B.L.; Zhang, L.F.; Li, Y.Z. Histone H2B monoubiquitination is involved in regulating the dynamics of microtubules during the defense response to Verticillium dahliae toxins in Arabidopsis. Plant Physiol. 2014, 164, 1857–1865. [Google Scholar] [CrossRef] [Green Version]
- Schnell, J.D.; Hicke, L. Non-traditional functions of ubiquitin and ubiquitin-binding proteins. J. Biol. Chem. 2003, 278, 35857–35860. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, D.; Zhang, H.; Hong, Y.; Huang, L.; Liu, S.; Li, X.; Ouyang, Z.; Song, F. Tomato histone H2B monoubiquitination enzymes SlHUB1 and SlHUB2 contribute to disease resistance against Botrytis cinerea through modulating the balance between SA- and JA/ET-mediated signaling pathways. BMC Plant Biol. 2015, 15, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhawan, R.; Luo, H.; Foerster, A.M.; Abuqamar, S.; Du, H.N.; Briggs, S.D.; Scheid, O.M.; Mengiste, T. HISTONE MONOUBIQUITINATION1 interacts with a subunit of the mediator complex and regulates defense against necrotrophic fungal pathogens in Arabidopsis. Plant Cell 2009, 21, 1000–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, Q.; Zhao, L.; Fu, S.; Qin, L.; Wei, Y.; Fu, Y.B.; Wang, H. Arabidopsis UBC22, an E2 able to catalyze lysine-11 specific ubiquitin linkage formation, has multiple functions in plant growth and immunity. Plant Sci. 2020, 297, 110520. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Cao, L.; Miu, W.; Cao, R.; Peng, M.; Wan, W.; Huang, L.-J. Molecular rewiring of the jasmonate signaling pathway to control auxin-responsive gene expression. Cells 2020, 9, 641. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.M.; Sun, X.L.; Dong, W.X.; Wang, G.C.; Chen, Z.M. Herbivore species, infestation time, and herbivore density affect induced volatiles in tea plants. Chemoecology 2014, 24, 1–14. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Khare, S.; Singh, N.B.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Bano, C.; Yadav, R.K.; Amist, N. Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. J. Plant Biol. 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Scott, E.R.; Li, X.; Wei, J.P.; Kfoury, N.; Morimoto, J.; Guo, M.M.; Agyei, A.; Robbat, A.; Ahmed, S.; Cash, S.B.; et al. Changes in tea plant secondary metabolite profiles as a function of leafhopper density and damage. Front. Plant Sci. 2020, 11, 636. [Google Scholar] [CrossRef]
- Gantner, M.; Najda, A.; Piesik, D. Effect of phenolic acid content on acceptance of hazel cultivars by filbert aphid. Plant Prot. Sci. 2019, 55, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Wallis, C.M.; Galarneau, E.R.A. Phenolic compound induction in plant-microbe and plant-insect interactions: A meta-analysis. Front. Plant Sci. 2020, 11, 580753. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Stringlis, I.A.; De Jonge, R.; Pieterse, C.M.J. The age of coumarins in plant-microbe interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrero, A.F.; Del Pino, M.M.H.; Portero, A.G.; Burón, P.A.; Arteaga, J.F.; Alquézar, J.B.; Díaz, C.E.; Coloma, A.G. Terpenes and polyacetylenes from cultivated Artemisia granatensis boiss (Royal chamomile) and their defensive properties. Phytochemistry 2013, 94, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.A.; Ode, P.J.; Gols, R.; Ali, J. Population- and species-based variation of webworm-parasitoid interactions in hogweeds (Heracelum Spp.) in the Netherlands. Environ. Entomol. 2020, 49, 924–930. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, S.; Wang, S.; Shan, W.; Wang, X.; Lin, Y.; Su, F.; Yang, Z.; Yu, X. Defensive responses of tea plants (Camellia sinensis) against tea green leafhopper attack: A multi-omics study. Front. Plant Sci. 2020, 10, 1705. [Google Scholar] [CrossRef]
- Perkovich, C.; Ward, D. Differentiated plant defense strategies: Herbivore community dynamics affect plant–herbivore interactions. Ecosphere 2022, 13, e3935. [Google Scholar] [CrossRef]
- Kariñho-Betancourt, E. Plant-herbivore interactions and secondary metabolites of plants: Ecological and evolutionary perspectives. Bot. Sci. 2018, 96, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; De Schutter, K.; Van Damme, E.J.M.; Smagghe, G. Can plant lectins help to elucidate insect lectin-mediated immune response? Insects 2021, 12, 497. [Google Scholar] [CrossRef]
- Künstler, A.; Gullner, G.; Ádám, A.L.; Nagy, J.K.; Király, L. The versatile roles of sulfur-containing biomolecules in plant defense—A road to disease resistance. Plants 2020, 9, 1705. [Google Scholar] [CrossRef]
- Chen, X.; Li, S.; Zhao, X.; Zhu, X.; Wang, Y.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y. Modulation of (Homo)glutathione metabolism and H2O2 accumulation during soybean cyst nematode infections in susceptible and resistant soybean cultivars. Int. J. Mol. Sci. 2020, 21, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sontowski, R.; Gorringe, N.J.; Pencs, S.; Schedl, A.; Touw, A.J.; van Dam, N.M. Same difference? Low and high glucosinolate Brassica rapa varieties show similar responses upon feeding by two specialist root herbivores. Front. Plant Sci. 2019, 10, 1451. [Google Scholar] [CrossRef] [PubMed]
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant defense chemicals against insect pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Huffaker, A.; Kaplan, F.; Vaughan, M.M.; Dafoe, N.J.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E.A.; Schmelz, E.A. Novel acidic sesquiterpenoids constitute a dominant class of pathogen-induced phytoalexins in maize. Plant Physiol. 2011, 156, 2082–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaleva, V.; Bukhteeva, I.; Kit, O.Y.; Nesmelova, I.V. Plant defensins from a structural perspective. Int. J. Mol. Sci. 2020, 21, 5307. [Google Scholar] [CrossRef]
- Gajger, I.T.; Dar, S.A. Plant allelochemicals as sources of insecticides. Insects 2021, 12, 189. [Google Scholar] [CrossRef]
- Liu, X.; Vrieling, K.; Klinkhamer, P.G.L. Interactions between plant metabolites affect herbivores: A study with pyrrolizidine alkaloids and chlorogenic acid. Front. Plant Sci. 2017, 8, 903. [Google Scholar] [CrossRef] [Green Version]
- Easson, M.L.A.E.; Malka, O.; Paetz, C.; Hojná, A.; Reichelt, M.; Stein, B.; van Brunschot, S.; Feldmesser, E.; Campbell, L.; Colvin, J.; et al. Activation and detoxification of cassava cyanogenic glucosides by the whitefly Bemisia tabaci. Sci. Rep. 2021, 11, 13244. [Google Scholar] [CrossRef]
- Huang, T.; Jander, G.; De Vos, M. Non-protein amino acids in plant defense against insect herbivores: Representative cases and opportunities for further functional analysis. Phytochemistry 2011, 72, 1531–1537. [Google Scholar] [CrossRef]
- Hodge, S.; Ward, J.L.; Galster, A.M.; Beale, M.H.; Powell, G. The effects of a plant defence priming compound, β-aminobutyric acid, on multitrophic interactions with an insect herbivore and a hymenopterous parasitoid. BioControl 2011, 56, 699–711. [Google Scholar] [CrossRef]
- Scholz, S.S.; Reichelt, M.; Mekonnen, D.W.; Ludewig, F.; Mithöfer, A. Insect herbivory-elicited gaba accumulation in plants is a wound-induced, direct, systemic, and jasmonate-independent defense response. Front. Plant Sci. 2015, 6, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, e1862565-2. [Google Scholar] [CrossRef] [PubMed]
- Staszek, P.; Krasuska, U.; Bederska-Błaszczyk, M.; Gniazdowska, A. Canavanine increases the content of phenolic compounds in tomato (Solanum lycopersicum L.) Roots. Plants 2020, 9, 1595. [Google Scholar] [CrossRef] [PubMed]
- Mitri, C.; Soustelle, L.; Framery, B.; Bockaert, J.; Parmentier, M.L.; Grau, Y. Plant insecticide l-canavanine repels drosophila via the insect orphan GPCR DmX. PLoS Biol. 2009, 7, e1000147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.T.; Murthy, H.N.; Park, S.Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [Green Version]
- Ramabulana, A.T.; Steenkamp, P.A.; Madala, N.E.; Dubery, I.A. Profiling of altered metabolomic states in Bidens pilosa leaves in response to treatment by methyl jasmonate and methyl salicylate. Plants 2020, 9, 1275. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant Responses to Herbivory, Wounding, and Infection. Int. J. Mol. Sci. 2022, 23, 7031. https://doi.org/10.3390/ijms23137031
Mostafa S, Wang Y, Zeng W, Jin B. Plant Responses to Herbivory, Wounding, and Infection. International Journal of Molecular Sciences. 2022; 23(13):7031. https://doi.org/10.3390/ijms23137031
Chicago/Turabian StyleMostafa, Salma, Yun Wang, Wen Zeng, and Biao Jin. 2022. "Plant Responses to Herbivory, Wounding, and Infection" International Journal of Molecular Sciences 23, no. 13: 7031. https://doi.org/10.3390/ijms23137031