
Biocontrol Potential of Endophytic Plant-Growth-Promoting Bacteria against Phytopathogenic Viruses: Molecular Interaction with the Host Plant and Comparison with Chitosan

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Effect of ePGPB Strains against Target Viruses
2.1.1. Plant Height: A Growth Promotion Parameter
2.1.2. Systemic Symptomatic Leaves and Symptom Severity: Biocontrol Parameters
2.1.3. Relative Quantification of Virus
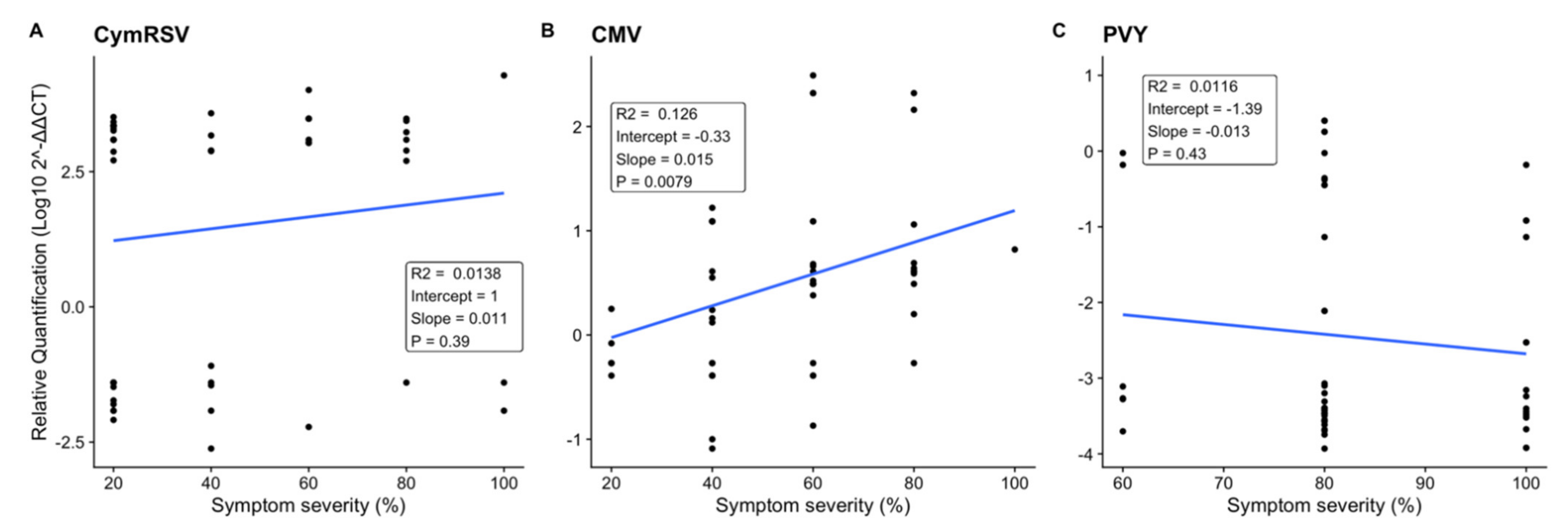
2.2. Effect of Symptom Severity on Virus Quantification
2.3. Effect of ePGPB Strains on Defense-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Microbial Strains
4.1.1. Plant Material
4.1.2. ePGPB Strains
4.1.3. Chitosan-Based Resistance Inductor
4.1.4. Phytopathogenic Viral Strains
4.2. Plant Growth Promotion, Biocontrol, and Defense-Related Gene Expression Bioassays
4.2.1. Experimental Setup
4.2.2. Inoculation of Plants with ePGPB Strains and Chitosan-Based Product
4.2.3. Virus Inoculation, Phenotypic Analysis, and Leaf Sampling
4.3. Relative Quantification of RNA
4.3.1. Relative Quantification of Virus
4.3.2. Relative Quantification of Defense-Related Genes
4.4. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hadidi, A.; Barba, M. Economic Impact of Pome and Stone Fruit Viruses and Viroids. In Virus and Virus Like Diseases of Pome and Stone Fruits; The American Phytopathological Society: St. Paul, MN, USA, 2011; Volume 1, pp. 1–7. [Google Scholar]
- Ziebell, H.; Carr, J.P. Cross-Protection: A Century of Mystery. Adv. Virus Res. 2010, 76, 211–264. [Google Scholar]
- Lewsey, M.; Palukaitis, P.; Carr, J.P. Plant-Virus Interactions: Defence and Counter-Defence. Mol. Asp. Plant Dis. Resist. 2009, 34, 134–176. [Google Scholar]
- Yu, A.V.; Yu, S.M. Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms. Plants 2019, 8, 575. [Google Scholar]
- King, A.M.Q.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011; Volume 9, ISBN 0123846854. [Google Scholar]
- Souiri, A.; Khataby, K.; Kasmi, Y.; Zemzami, M.; Amzazi, S.; Ennaji, M.M. Emerging and Reemerging Viral Diseases of Solanaceous Crops and Management Strategies for Detection and Eradication. In Emerging and Reemerging Viral Pathogens; Elsevier: Amsterdam, The Netherlands, 2020; pp. 847–877. [Google Scholar]
- Soosaar, J.L.M.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of Plant Resistance to Viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Koonin, E.V. Common Origins and Host-Dependent Diversity of Plant and Animal Viromes. Curr. Opin. Virol. 2011, 1, 322–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borah, B.K.; Sharma, S.; Kant, R.; Johnson, A.M.A.; Saigopal, D.V.R.; Dasgupta, I. Bacilliform DNA-containing Plant Viruses in the Tropics: Commonalities within a Genetically Diverse Group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Sastry, K.S.; Zitter, T.A. Management of Virus and Viroid Diseases of Crops in the Tropics. In Plant Virus and Viroid Diseases in the Tropics; Springer: Dodrecht, The Netherlands, 2014; pp. 149–480. [Google Scholar]
- Castle, S.; Palumbo, J.; Merten, P.; Cowden, C.; Prabhaker, N. Effects of Foliar and Systemic Insecticides on Whitefly Transmission and Incidence of Cucurbit yellow stunting disorder virus. Pest Manag. Sci. 2017, 73, 1462–1472. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.A.; Nagle, C.A.; MacVean, C.M.; Vallad, G.E.; van Santen, E.; Hutton, S.F. Comparing Host Plant Resistance, Repellent Mulches, and at-Plant Insecticides for Management of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) and Tomato yellow leaf curl virus. J. Econ. Entomol. 2019, 112, 236–243. [Google Scholar] [CrossRef]
- Pretty, J.; Bharucha, Z.P. Sustainable Intensification in Agricultural Systems. Ann. Bot. 2014, 114, 1571–1596. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.; Chen, Y.; Hao, X.; Wu, Y. Advances and Prospects in Biogenic Substances against Plant Virus: A Review. Pestic. Biochem. Physiol. 2017, 135, 15–26. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, D.-D.; Dong, X.-W.; Zhao, P.-B.; Chen, L.-L.; Song, X.-Y.; Wang, X.-J.; Chen, X.-L.; Shi, M.; Zhang, Y.-Z. Antimicrobial Peptaibols Induce Defense Responses and Systemic Resistance in Tobacco against Tobacco mosaic virus. FEMS Microbiol. Lett. 2010, 313, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Laimer, M.; Barba, M. Elimination of Systemic Pathogens by Thermotherapy, Tissue Culture, or in Vitro Micrografting. Virus and Virus-Like Diseases of Pome and Stone Fruits; APS: St. Paul, MN, USA, 2011; pp. 389–393. [Google Scholar]
- Wang, B.; Ma, Y.; Zhang, Z.; Wu, Z.; Wu, Y.; Wang, Q.; Li, M. Potato Viruses in China. Crop Prot. 2011, 30, 1117–1123. [Google Scholar] [CrossRef]
- Bellich, B.; D’Agostino, I.; Semeraro, S.; Gamini, A.; Cesàro, A. “The Good, the Bad and the Ugly” of Chitosans. Mar. Drugs 2016, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Kanmani, P.; Aravind, J.; Kamaraj, M.; Sureshbabu, P.; Karthikeyan, S. Environmental Applications of Chitosan and Cellulosic Biopolymers: A Comprehensive Outlook. Bioresour. Technol. 2017, 242, 295–303. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Abscisic Acid Is Involved in Chitosan-Induced Resistance to Tobacco necrosis virus (TNV). Plant Physiol. Biochem. 2008, 46, 1106–1111. [Google Scholar] [CrossRef]
- Lu, H.; Zhao, X.; Wang, W.; Yin, H.; Xu, J.; Bai, X.; Du, Y. Inhibition Effect on Tobacco mosaic virus and Regulation Effect on Calreticulin of Oligochitosan in Tobacco by Induced Ca2+ Influx. Carbohydr. Polym. 2010, 82, 136–142. [Google Scholar] [CrossRef]
- Mishra, S.; Jagadeesh, K.S.; Krishnaraj, P.U.; Prem, S. Biocontrol of Tomato leaf curl virus (ToLCV) in Tomato with Chitosan Supplemented Formulations of Pseudomonas sp. under Field Conditions. Aust. J. Crop Sci. 2014, 8, 347–355. [Google Scholar]
- Firmansyah, D. Use of Chitosan and Plant Growth Promoting Rhizobacteria to Control Squash mosaic virus on Cucumber Plants. Undergraduate Thesis, Bogor Agricultural University, Bogor, Indonesia, 2017. [Google Scholar]
- Iriti, M.; Varoni, E.M. Chitosan-Induced Antiviral Activity and Innate Immunity in Plants. Environ. Sci. Pollut. Res. 2015, 22, 2935–2944. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Bhore, S.J.; Ravichantar, N.; Loh, C.Y. Screening of Endophytic Bacteria Isolated from Leaves of Sambung Nyawa [Gynura procumbens (Lour.) Merr.] for Cytokinin-like Compounds. Bioinformation 2010, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Cipollini, D.; Rigsby, C.M.; Barto, E.K. Microbes as Targets and Mediators of Allelopathy in Plants. J. Chem. Ecol. 2012, 38, 714–727. [Google Scholar] [CrossRef]
- Phetcharat, P.; Duangpaeng, A. Screening of Endophytic Bacteria from Organic Rice Tissue for Indole Acetic Acid Production. Procedia Eng. 2012, 32, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Rashid, S.; Charles, T.C.; Glick, B.R. Isolation and Characterization of New Plant Growth-Promoting Bacterial Endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Ullah, A.; Mushtaq, H.; Fahad, S.; Shah, A.; Chaudhary, H.J. Plant Growth Promoting Potential of Bacterial Endophytes in Novel Association with Olea ferruginea and Withania coagulans. Microbiology 2017, 86, 119–127. [Google Scholar] [CrossRef]
- Raupach, G.S.; Liu, L.; Murphy, J.F.; Tuzun, S.; Kloepper, J.W. Induced Systemic Resistance in Cucumber and Tomato against Cucumber mosaic cucumovirus Using Plant Growth-Promoting Rhizobacteria (PGPR). Plant Dis. 1996, 80, 891–894. [Google Scholar] [CrossRef]
- Zehnder, G.W.; Yao, C.; Murphy, J.F.; Sikora, E.R.; Kloepper, J.W. Induction of Resistance in Tomato against Cucumber mosaic cucumovirus by Plant Growth-Promoting Rhizobacteria. Biocontrol 2000, 45, 127–137. [Google Scholar] [CrossRef]
- Murphy, J.F.; Reddy, M.S.; Ryu, C.-M.; Kloepper, J.W.; Li, R. Rhizobacteria-Mediated Growth Promotion of Tomato Leads to Protection against Cucumber mosaic virus. Phytopathology 2003, 93, 1301–1307. [Google Scholar] [CrossRef] [Green Version]
- Kandan, A.; Ramiah, M.; Vasanthi, V.J.; Radjacommare, R.; Nandakumar, R.; Ramanathan, A.; Samiyappan, R. Use of Pseudomonas fluorescens-Based Formulations for Management of Tomato spotted wilt virus (TSWV) and Enhanced Yield in Tomato. Biocontrol. Sci. Technol. 2005, 15, 553–569. [Google Scholar] [CrossRef]
- Kavino, M.; Harish, S.; Kumar, N.; Saravanakumar, D.; Damodaran, T.; Soorianathasundaram, K.; Samiyappan, R. Rhizosphere and Endophytic Bacteria for Induction of Systemic Resistance of Banana Plantlets against Bunchy Top Virus. Soil Biol. Biochem. 2007, 39, 1087–1098. [Google Scholar] [CrossRef]
- Wang, S.; Wu, H.; Qiao, J.; Ma, L.; Liu, J.; Xia, Y.; Gao, X. Molecular Mechanism of Plant Growth Promotion and Induced Systemic Resistance to Tobacco mosaic virus by Bacillus spp. J. Microbiol. Biotechnol. 2009, 19, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, C.; Zhai, X.; Shen, L.; Qian, Y.; Wang, F. Inactivation of Tobacco mosaic virus in Soil by Pseudomonas putida A3-m Strain to Prevent Virus Mosaic Disease. Afr. J. Microbiol. Res. 2012, 6, 6300–6307. [Google Scholar] [CrossRef]
- Kumar, S.; Chauhan, P.S.; Agrawal, L.; Raj, R.; Srivastava, A.; Gupta, S.; Mishra, S.K.; Yadav, S.; Singh, P.C.; Raj, S.K. Paenibacillus lentimorbus Inoculation Enhances Tobacco Growth and Extenuates the Virulence of Cucumber mosaic virus. PLoS ONE 2016, 11, e0149980. [Google Scholar] [CrossRef] [PubMed]
- Beris, D.; Theologidis, I.; Skandalis, N.; Vassilakos, N. Bacillus amyloliquefaciens Strain MBI600 Induces Salicylic Acid Dependent Resistance in Tomato Plants against Tomato spotted wilt virus and Potato virus Y. Sci. Rep. 2018, 8, 10320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Verma, J.; Srivastava, A.; Srivastava, S.; Prasad, V. Induced Antiviral resistance induction and growth promotion by the rhizobacterium Pantoea agglomerans. J. Glob. Biosci. 2019, 8, 6219–6235. [Google Scholar]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Alazem, M.; Lin, N. Roles of Plant Hormones in the Regulation of Host–Virus Interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Li, H.; Ding, X.; Wang, C.; Ke, H.; Wu, Z.; WANG, Y.; Liu, H.; Guo, J. Control of Tomato Yellow Leaf Curl Virus Disease by Enterobacter asburiae BQ9 as a Result of Priming Plant Resistance in Tomatoes. Turk. J. Biol. 2016, 40, 150–159. [Google Scholar] [CrossRef]
- Passera, A.; Venturini, G.; Battelli, G.; Casati, P.; Penaca, F.; Quaglino, F.; Bianco, P.A. Competition Assays Revealed Paenibacillus pasadenensis Strain R16 as a Novel Antifungal Agent. Microbiol. Res. 2017, 198, 16–26. [Google Scholar] [CrossRef]
- Passera, A.; Vacchini, V.; Cocetta, G.; Arpanahi, A.A.; Casati, P.; Ferrante, A.; Piazza, L. Towards Nutrition-Sensitive Agriculture: An Evaluation of Biocontrol Effects, Nutritional Value, and Ecological Impact of Bacterial Inoculants. Sci. Total Environ. 2020, 724, 138127. [Google Scholar] [CrossRef]
- Passera, A.; Compant, S.; Casati, P.; Maturo, M.G.; Battelli, G.; Quaglino, F.; Antonielli, L.; Salerno, D.; Brasca, M.; Toffolatti, S.L. Not Just a Pathogen? Description of a Plant-Beneficial Pseudomonas syringae Strain. Front. Microbiol. 2019, 10, 1409. [Google Scholar] [CrossRef] [Green Version]
- Hollings, M.; STONE, O.M.; Barton, R.J. Pathology, Soil Transmission and Characterization of Cymbidium Ringspot, a Virus from Cymbidium Orchids and White Clover (Trifolium repens). Ann. Appl. Biol. 1977, 85, 233–248. [Google Scholar] [CrossRef]
- Douine, L.; Quiot, J.B.; Marchoux, G.; Archange, P. Index of Plants Susceptible to Cucumber mosaic virus (CMV). Bibliographical Study. Ann. Phytopathol. 1979, 11, 439–475. [Google Scholar]
- Brunt, A.A.; Crabtree, K.; Dallwitz, A.J. Analítico: Viruses of Plants; Descriptions and Lists from the VIDE Database; CAB INTERNATIONAL: Wallingrod, UK, 1996. [Google Scholar]
- Yardimci, N.; Eryigit, H. Identification of Cucumber mosaic virus in Tomato (Lycopersicon esculentum) Growing Areas in the North-west Mediterranean Region of Turkey. N. Z. J. Crop Hortic. Sci. 2006, 34, 173–175. [Google Scholar] [CrossRef]
- Edwardson, J.R.; Christie, R.G. Cucumoviruses. In CRC Handbook of Viruses Infecting Legumes; CRC Press: Boca Raton, FL, USA, 1991; pp. 293–319. [Google Scholar]
- CABI Knowledge Bank. Potato virus X. Available online: https://www.plantwise.org/knowledgebank/datasheet/43759 (accessed on 28 October 2020).
- Miozzi, L.; Vaira, A.M.; Brilli, F.; Casarin, V.; Berti, M.; Ferrandino, A.; Nerva, L.; Accotto, G.P.; Lanfranco, L. Arbuscular Mycorrhizal Symbiosis Primes Tolerance to Cucumber mosaic virus in Tomato. Viruses 2020, 12, 675. [Google Scholar] [CrossRef]
- Palukaitis, P.; García-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar]
- Ephytia Inrea Potato—Potato virus X (PVX). Available online: http://ephytia.inra.fr/en/C/21025/Potato-Potato-virus-X-PVX (accessed on 28 October 2020).
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of Endophytes for Sustainable Agricultural Intensification. Mol. Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Chirkov, S.N.; Il’ina, A.V.; Surgucheva, N.A.; Letunova, E.V.; Varitsev, Y.A.; Tatarinova, N.Y.; Varlamov, V.P. Effect of Chitosan on Systemic Viral Infection and Some Defense Responses in Potato Plants. Russ. J. Plant Physiol. 2001, 48, 774–779. [Google Scholar] [CrossRef]
- Rendina, N.; Nuzzaci, M.; Scopa, A.; Cuypers, A.; Sofo, A. Chitosan-Elicited Defense Responses in Cucumber mosaic virus (CMV)-Infected Tomato Plants. J. Plant Physiol. 2019, 234, 9–17. [Google Scholar] [CrossRef]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of Reference Genes for Gene Expression Studies in Virus-Infected Nicotiana benthamiana Using Quantitative Real-Time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef]
- Nakasugi, K.; Crowhurst, R.N.; Bally, J.; Wood, C.C.; Hellens, R.P.; Waterhouse, P.M. De Novo Transcriptome Sequence Assembly and Analysis of RNA Silencing Genes of Nicotiana benthamiana. PLoS ONE 2013, 8, e59534. [Google Scholar] [CrossRef] [Green Version]
- Bally, J.; Nakasugi, K.; Jia, F.; Jung, H.; Ho, S.Y.W.; Wong, M.; Paul, C.M.; Naim, F.; Wood, C.C.; Crowhurst, R.N. The Extremophile Nicotiana benthamiana Has Traded Viral Defence for Early Vigour. Nat. Plants 2015, 1, 15165. [Google Scholar] [CrossRef]
- Guo, Q.; Li, Y.; Lou, Y.; Shi, M.; Jiang, Y.; Zhou, J.; Sun, Y.; Xue, Q.; Lai, H. Bacillus Amyloliquefaciens Ba13 Induces Plant Systemic Resistance and Improves Rhizosphere Microecology against Tomato Yellow Leaf Curl Virus Disease. Appl. Soil Ecol. 2019, 137, 154–166. [Google Scholar] [CrossRef]
- Thakur, A. Evaluation of Biological Control Strategies against a Range of Plant Pathogens. Master’s Thesis, Agricultural University of Athens, Athens, Greece, 2017. [Google Scholar]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, M.; Wang, W.E.I.; Li, D.; Huang, Q.; Wang, Y.; Zheng, X.; Zhang, Z. Silencing of G Proteins Uncovers Diversified Plant Responses When Challenged by Three Elicitors in Nicotiana benthamiana. Plant Cell Environ. 2012, 35, 72–85. [Google Scholar] [CrossRef]
- Wiermer, M.; Feys, B.J.; Parker, J.E. Plant Immunity: The EDS1 Regulatory Node. Curr. Opin. Plant Biol. 2005, 8, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Spoel, S.H.; Koornneef, A.; Claessens, S.M.C.; Korzelius, J.P.; van Pelt, J.A.; Mueller, M.J.; Buchala, A.J.; Métraux, J.-P.; Brown, R.; Kazan, K. NPR1 Modulates Cross-Talk between Salicylate-and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol. Plant Cell 2003, 15, 760–770. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a Multifaceted Hormone to Combat Disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [Green Version]
- Chirkov, S.N. The Antiviral Activity of Chitosan. Appl. Biochem. Microbiol. 2002, 38, 1–8. [Google Scholar] [CrossRef]
- Beatrice, C.; Linthorst, J.M.H.; Cinzia, F.; Luca, R. Enhancement of PR1 and PR5 Gene Expressions by Chitosan Treatment in Kiwifruit Plants Inoculated with Pseudomonas syringae pv. Actinidiae. Eur. J. Epidemiol. 2017, 148, 163–179. [Google Scholar] [CrossRef]
- Bulgari, D.; Casati, P.; Crepaldi, P.; Daffonchio, D.; Quaglino, F.; Brusetti, L.; Bianco, P.A. Restructuring of Endophytic Bacterial Communities in Grapevine Yellows-Diseased and Recovered Vitis vinifera L. Plants. Appl. Environ. Microbiol. 2011, 77, 5018–5022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgari, D.; Bozkurt, A.I.; Casati, P.; Çağlayan, K.; Quaglino, F.; Bianco, P.A. Endophytic Bacterial Community Living in Roots of Healthy and ‘Candidatus Phytoplasma mali’-Infected Apple (Malus domestica, Borkh.) Trees. Antonie Van Leeuwenhoek 2012, 102, 677–687. [Google Scholar] [CrossRef]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma Harzianum T-22 Induces Systemic Resistance in Tomato Infected by Cucumber mosaic virus. Front. Plant Sci. 2016, 7, 1520. [Google Scholar] [CrossRef] [Green Version]
- Townsend, G.R. Methods for Estimating Losses Caused by Diseases in Fungicide Experiments. Plant Dis. Report. 1943, 27, 340–343. [Google Scholar]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and Effective Method for RNA Extraction from Different Tissues of Grapevine and Other Woody Plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2− ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Passera, A.; Rossato, M.; Oliver, J.S.; Battelli, G.; Shahzad, G.-I.-R.; Cosentino, E.; Sage, J.M.; Toffolatti, S.L.; Lopatriello, G.; Davis, J.R. Characterization of Lysinibacillus fusiformis Strain S4C11: In Vitro, in Planta, and in Silico Analyses Reveal a Plant-Beneficial Microbe. Microbiol. Res. 2021, 244, 126665. [Google Scholar] [CrossRef]
- Feng, J.-L.; Chen, S.-N.; Tang, X.-S.; Ding, X.-F.; Du, Z.-Y.; Chen, J.-S. Quantitative Determination of Cucumber mosaic virus Genome RNAs in Virions by Real-Time Reverse Transcription-Polymerase Chain Reaction. Acta Biochim. Biophys. Sin. 2006, 38, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-S.; Paul, D.; Ryu, K.-R.; Kim, E.-Y.; Kim, Y.-K. Bacillus Vallismortis Strain EXTN-1 Mediated Systemic Resistance against Potato virus Y and X in the Field. Plant Pathol. J. 2006, 22, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Nie, B.; Liu, J.; Song, B. A Reexamination of the Effectiveness of Ribavirin on Eradication of Viruses in Potato Plantlets in Vitro Using ELISA and Quantitative RT-PCR. Am. J. Potato Res. 2014, 91, 304–311. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Biocontrol Assay (n#) | Defense-Related Gene Expression Assay (n#) |
|---|---|---|
| NT-C | 10 | 5 |
| CymRSV, CMV, PVX, PVY | 10 | - |
| R8, R16, 255-7, 260-02 | 10 | 5 |
| CHI-C | 5 | 5 |
| CHI | 5 | - |
| Primer Pair | Sequence 5′-3′ | Fragment (Target Gene) | Bibliography |
|---|---|---|---|
| PP2A—forward | GACCCTGATGTTGATGTTCGCT | 123bp | [59] |
| PP2A—reverse | GAGGGATTTGAAGAGAGATTTC | ||
| CymRSV—forward | GTA CAT GCG TCA CTT GGG GA | 195bp (RNA polymerase) | [79] |
| CymRSV—reverse | TCT CAG CAT CTT CCA ACC GC | ||
| CMV—forward | CTG GCG ACA ATC TGG AGG GA | 153bp (Movement protein) | [80] |
| CMV—reverse | CGA TAA CGA CAG CAA AAC AC | ||
| PVX—forward | GCC CAA TTG TTA CAC ACC | 101bp (Coat protein) | [81] |
| PVX—reverse | CTA GCC TCA TCT TAA TG | ||
| PVY—forward | GGT AGC ACA ACT ATA CGG TGC | 100bp (Coat protein) | [82] |
| PVY—reverse | GAT GTT TGG GGT CGA TCC A |
| Primer Pair | Sequence 5′-3′ | Fragment | Bibliography |
|---|---|---|---|
| PP2A—forward | GACCCTGATGTTGATGTTCGCT | 123 bp | [59] |
| PP2A—reverse | GAGGGATTTGAAGAGAGATTTC | ||
| NPR1—forward | GGC CTT GCC TCA TGA TAT TG | 187 bp | [66] |
| NPR1—reverse | GCT ACA GCA TAA TGG AGA GC | ||
| PR2B—forward | CTAAAGAGGGTAGCCCAAGA | 147 bp | [66] |
| PR2B—reverse | GTCCCAAACTCCACCAGAGA | ||
| EDS1—forward | GGACAATGGGAGAAGCAGAA | 118 bp | [66] |
| EDS1—reverse | GAACGCATCATAATACCCGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahzad, G.-i.-R.; Passera, A.; Maldera, G.; Casati, P.; Marcello, I.; Bianco, P.A. Biocontrol Potential of Endophytic Plant-Growth-Promoting Bacteria against Phytopathogenic Viruses: Molecular Interaction with the Host Plant and Comparison with Chitosan. Int. J. Mol. Sci. 2022, 23, 6990. https://doi.org/10.3390/ijms23136990
Shahzad G-i-R, Passera A, Maldera G, Casati P, Marcello I, Bianco PA. Biocontrol Potential of Endophytic Plant-Growth-Promoting Bacteria against Phytopathogenic Viruses: Molecular Interaction with the Host Plant and Comparison with Chitosan. International Journal of Molecular Sciences. 2022; 23(13):6990. https://doi.org/10.3390/ijms23136990
Chicago/Turabian StyleShahzad, Gul-i-Rayna, Alessandro Passera, Giusva Maldera, Paola Casati, Iriti Marcello, and Piero Attilio Bianco. 2022. "Biocontrol Potential of Endophytic Plant-Growth-Promoting Bacteria against Phytopathogenic Viruses: Molecular Interaction with the Host Plant and Comparison with Chitosan" International Journal of Molecular Sciences 23, no. 13: 6990. https://doi.org/10.3390/ijms23136990