
Short-Term, Voluntary Exercise Affects Morpho-Functional Maturation of Adult-Generated Neurons in Rat Hippocampus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
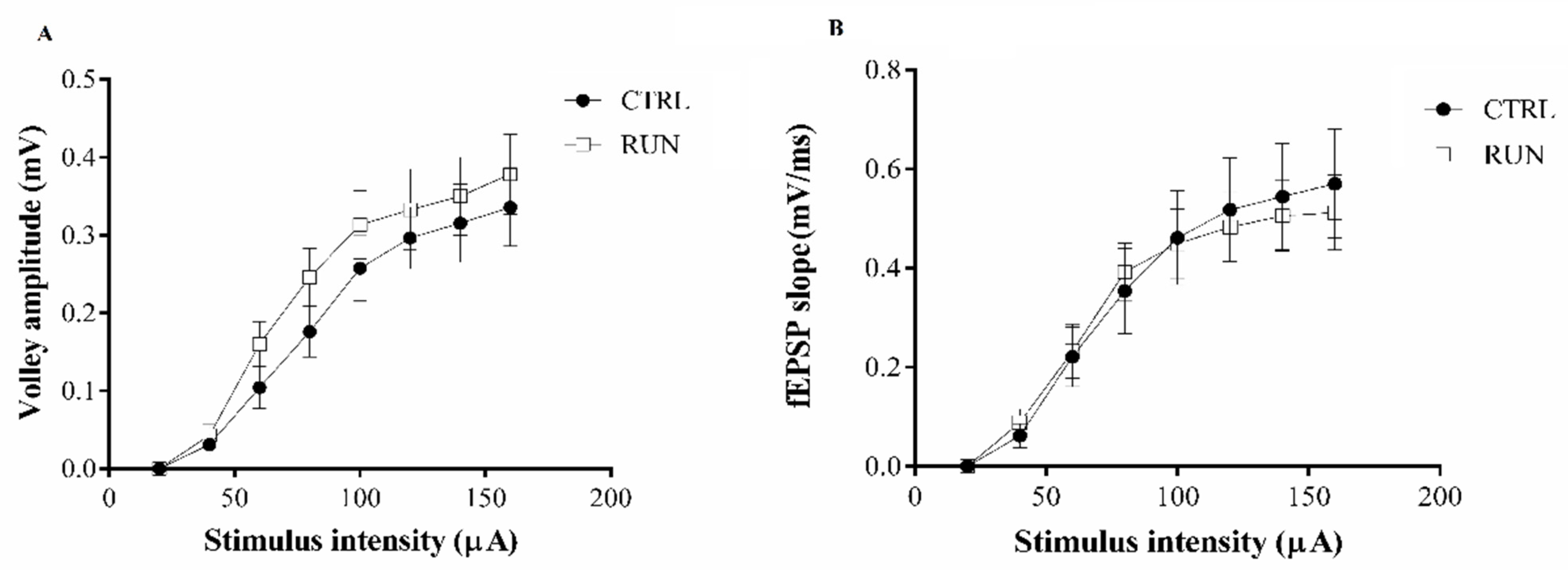
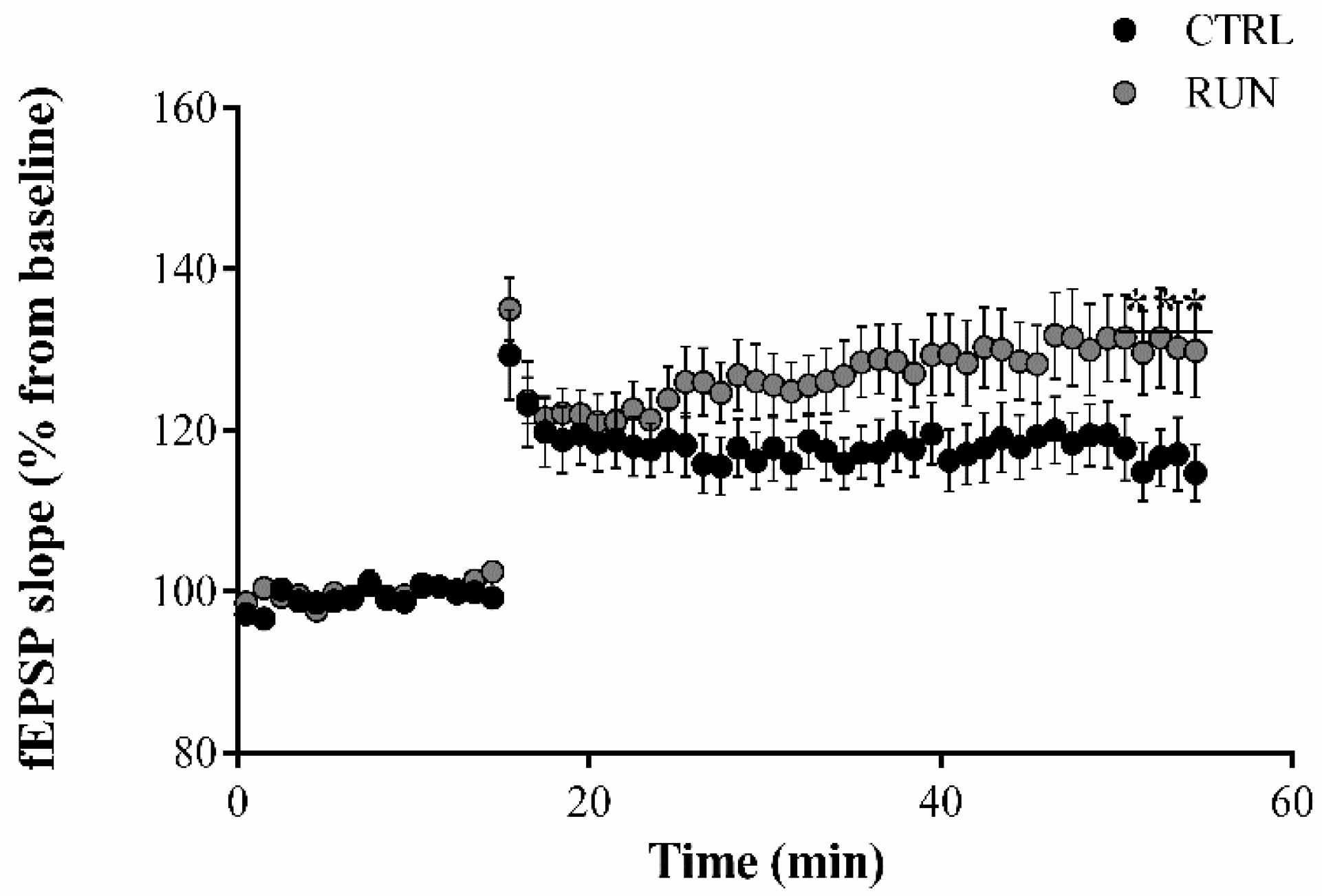
2.1. Electrophysiological Analysis
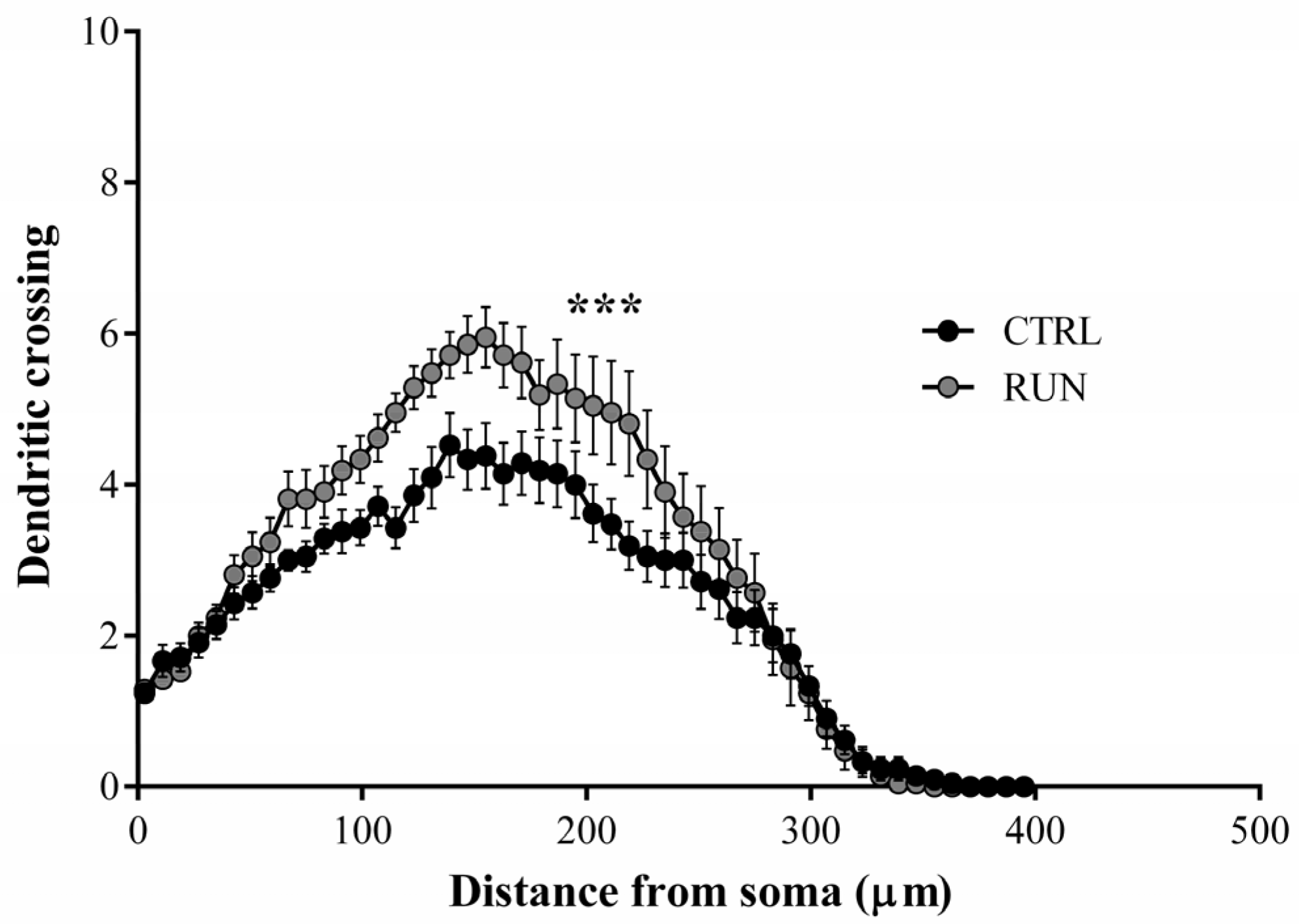
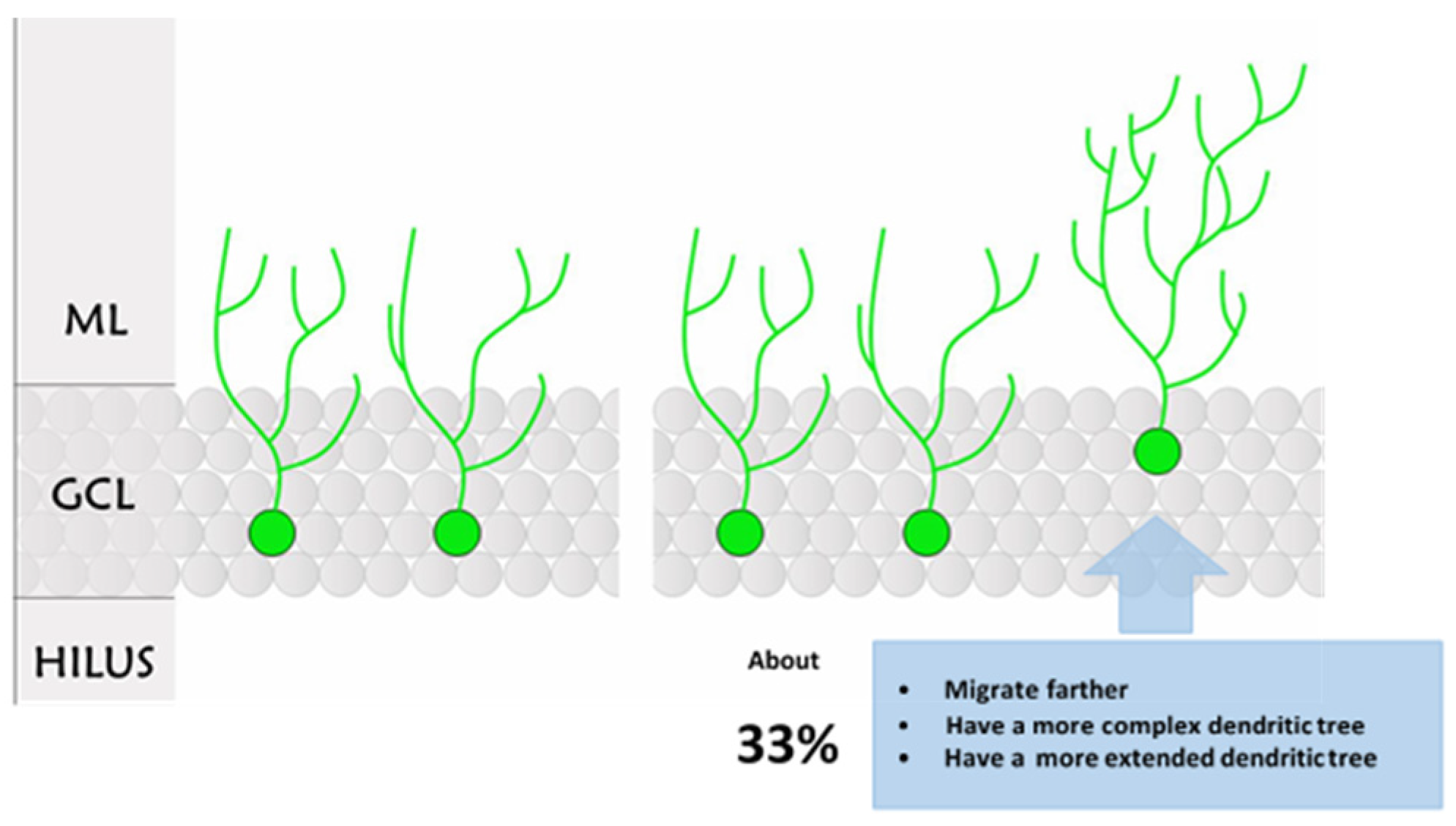
2.2. Morphological Analysis
3. Discussion
4. Materials and Methods
4.1. Animals and Running Protocol
4.2. Electrophysiological Experiments
4.2.1. Slices Preparation
4.2.2. Electrophysiological Field Recordings
4.3. Morphological Analysis
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ambrogini, P.; Betti, M.; Galati, C.; Di Palma, M.; Lattanzi, D.; Savelli, D.; Galli, F.; Cuppini, R.; Minelli, A. Alpha-Tocopherol and Hippocampal Neural Plasticity in Physiological and Pathological Conditions. Int. J. Mol. Sci. 2016, 17, 2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grindey, A.; Bradshaw, T. Do different adverse childhood experiences lead to specific symptoms of psychosis in adulthood? A systematic review of the current literature. Int. J. Ment. Health Nurs. 2022. [Google Scholar] [CrossRef] [PubMed]
- Hakamata, Y.; Suzuki, Y.; Kobashikawa, H.; Hori, H. Neurobiology of early life adversity: A systematic review of meta-analyses towards an integrative account of its neurobiological trajectories to mental disorders. Front. Neuroendocrinol. 2022, 65, 100994. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.; Beck, M.M.; Bilenberg, N.; Wienecke, J.; Astrup, A.; Lundbye-Jensen, J. Effects of Exercise on Cognitive Performance in Children and Adolescents with ADHD: Potential Mechanisms and Evidence-based Recommendations. J. Clin. Med. 2019, 8, 841. [Google Scholar] [CrossRef] [Green Version]
- Hillman, C.H.; McDonald, K.M.; Logan, N.E. A Review of the Effects of Physical Activity on Cognition and Brain Health across Children and Adolescence. Build. Future Health Well-Being Thriving Toddlers Young Child. 2020, 95, 116–126. [Google Scholar]
- Ramirez-Amaya, V.; Marrone, D.F.; Gage, F.H.; Worley, P.F.; Barnes, C.A. Integration of new neurons into functional neural networks. J. Neurosci. 2006, 26, 12237–12241. [Google Scholar] [CrossRef]
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and functional implications of adult neurogenesis. Cell 2008, 132, 645–660. [Google Scholar] [CrossRef] [Green Version]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [Green Version]
- Steiner, B.; Zurborg, S.; Hörster, H.; Fabel, K.; Kempermann, G. Differential 24 h responsiveness of Prox1-expressing precursor cells in adult hippocampal neurogenesis to physical activity, environmental enrichment, and kainic acid-induced seizures. Neuroscience 2008, 154, 521–529. [Google Scholar] [CrossRef]
- Umegaki, H.; Sakurai, T.; Arai, H. Active Life for Brain Health: A Narrative Review of the Mechanism Underlying the Protective Effects of Physical Activity on the Brain. Front. Aging Neurosci. 2021, 13, 761674. [Google Scholar] [CrossRef]
- Johnson, R.A.; Rhodes, J.S.; Jeffrey, S.L.; Garland, T., Jr.; Mitchell, G.S. Hippocampal brain-derived neurotrophic factor but not neurotrophin-3 increases more in mice selected for increased voluntary wheel running. Neuroscience 2003, 121, 1–7. [Google Scholar] [CrossRef]
- Moon, H.Y.; Kim, S.H.; Yang, Y.R.; Song, P.; Yu, H.S.; Park, H.G.; Hwang, O.; Lee-Kwon, W.; Seo, J.K.; Hwang, D.; et al. Macrophage migration inhibitory factor mediates the antidepressant actions of voluntary exercise. Proc. Natl. Acad. Sci. USA 2012, 109, 13094–13099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeusen, R.; Thorré, K.; Chaouloff, F.; Sarre, S.; De Meirleir, K.; Ebinger, G.; Michotte, Y. Effects of tryptophan and/or acute running on extracellular 5-HT and 5-HIAA levels in the hippocampus of food-deprived rats. Brain Res. 1996, 740, 245–252. [Google Scholar] [CrossRef]
- Klempin, F.; Babu, H.; De Pietri Tonelli, D.; Alarcon, E.; Fabel, K.; Kempermann, G. Oppositional effects of serotonin receptors 5-HT1a, 2, and 2c in the regulation of adult hippocampal neurogenesis. Front. Mol. Neurosci. 2010, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Wang, H.; Zeng, Y.; Qu, Y.; Liu, Q.; Zhao, F.; Duan, J.; Jiang, Y.; Li, S.; Ying, J.; et al. Physical exercise promotes brain remodeling by regulating epigenetics, neuroplasticity and neurotrophins. Rev. Neurosci. 2021, 32, 615–629. [Google Scholar] [CrossRef]
- Jin, K.; Zhu, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Greenberg, D.A. Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11946–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabel, K.; Fabel, K.; Tam, B.; Kaufer, D.; Baiker, A.; Simmons, N.; Kuo, C.J.; Palmer, T.D. VEGF is necessary for exercise-induced adult hippocampal neurogenesis. Eur. J. Neurosci. 2003, 18, 2803–2812. [Google Scholar] [CrossRef] [PubMed]
- Carro, E.; Nuñez, A.; Busiguina, S.; Torres-Aleman, I. Circulating insulin-like growth factor I mediates effects of exercise on the brain. J. Neurosci. 2000, 20, 2926–2933. [Google Scholar] [CrossRef]
- Trejo, J.L.; Carro, E.; Torres-Aleman, I. Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J. Neurosci. 2001, 21, 1628–1634. [Google Scholar] [CrossRef] [Green Version]
- El-Sayes, J.; Harasym, D.; Turco, C.V.; Locke, M.B.; Nelson, A.J. Exercise-Induced Neuroplasticity: A Mechanistic Model and Prospects for Promoting Plasticity. Neuroscientist 2019, 25, 65–85. [Google Scholar] [CrossRef]
- Ambrogini, P.; Lattanzi, D.; Ciuffoli, S.; Betti, M.; Fanelli, M.; Cuppini, R. Physical exercise and environment exploration affect synaptogenesis in adult-generated neurons in the rat dentate gyrus: Possible role of BDNF. Brain Res. 2013, 1534, 1–12. [Google Scholar] [CrossRef]
- Ambrogini, P.; Cuppini, R.; Lattanzi, D.; Ciuffoli, S.; Frontini, A.; Fanelli, M. Synaptogenesis in adult-generated hippocampal granule cells is affected by behavioral experiences. Hippocampus 2010, 20, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.C.; Inoue, K.; Okamoto, M.; Liu, Y.F.; Matsui, T.; Yook, J.S.; Soya, H. Voluntary resistance running induces increased hippocampal neurogenesis in rats comparable to load-free running. Neurosci. Lett. 2013, 537, 6–10. [Google Scholar] [CrossRef]
- Jagasia, R.; Steib, K.; Englberger, E.; Herold, S.; Faus-Kessler, T.; Saxe, M.; Gage, F.H.; Song, H.; Lie, D.C. GABA-cAMP response element-binding protein signaling regulates maturation and survival of newly generated neurons in the adult hippocampus. J. Neurosci. 2009, 29, 7966–7977. [Google Scholar] [CrossRef] [Green Version]
- Lattanzi, D.; Di Palma, M.; Cuppini, R.; Ambrogini, P. GABAergic Input Affects Intracellular Calcium Levels in Developing Granule Cells of Adult Rat Hippocampus. Int. J. Mol. Sci. 2020, 21, 1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuczewski, N.; Porcher, C.; Lessmann, V.; Medina, I.; Gaiarsa, J.L. Activity-dependent dendritic release of BDNF and biological consequences. Mol. Neurobiol. 2009, 39, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B. BDNF and activity-dependent synaptic modulation. Learn. Mem. 2003, 10, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Scharfman, H.; Goodman, J.; Macleod, A.; Phani, S.; Antonelli, C.; Croll, S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp. Neurol. 2005, 192, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.H.; Yamaguchi, M.; Eisch, A.J. Dynamic expression of TrkB receptor protein on proliferating and maturing cells in the adult mouse dentate gyrus. Hippocampus 2008, 18, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Luikart, B.W.; Birnbaum, S.; Chen, J.; Kwon, C.H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron 2008, 59, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, J.S.; Kee, N.; Wojtowicz, J.M. Effects of adult neurogenesis on synaptic plasticity in the rat dentate gyrus. J. Neurophysiol. 2001, 85, 2423–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.C.; Huddleston, D.E.; Brickman, A.M.; Sosunov, A.A.; Hen, R.; McKhann, G.M.; Sloan, R.; Gage, F.H.; Brown, T.R.; Small, S.A. An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proc. Natl. Acad. Sci. USA 2007, 104, 5638–5643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uda, M.; Ishido, M.; Kami, K.; Masuhara, M. Effects of chronic treadmill running on neurogenesis in the dentate gyrus of the hippocampus of adult rat. Brain Res. 2006, 1104, 64–72. [Google Scholar] [CrossRef]
- Wu, C.W.; Chang, Y.T.; Yu, L.; Chen, H.I.; Jen, C.J.; Wu, S.Y.; Lo, C.P.; Kuo, Y.M. Exercise enhances the proliferation of neural stem cells and neurite growth and survival of neuronal progenitor cells in dentate gyrus of middle-aged mice. J. Appl. Physiol. (1985) 2008, 105, 1585–1594. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, N.C.; Castello, N.; Cotman, C.W. Exercise and time-dependent benefits to learning and memory. Neuroscience 2010, 167, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Creer, D.J.; Romberg, C.; Saksida, L.M.; van Praag, H.; Bussey, T.J. Running enhances spatial pattern separation in mice. Proc. Natl. Acad. Sci. USA 2010, 107, 2367–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Bolz, L.; Heigele, S.; Bischofberger, J. Running Improves Pattern Separation during Novel Object Recognition. Brain Plast. 2015, 1, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Kamijo, S.; Ishii, Y.; Horigane, S.I.; Suzuki, K.; Ohkura, M.; Nakai, J.; Fujii, H.; Takemoto-Kimura, S.; Bito, H. A Critical Neurodevelopmental Role for L-Type Voltage-Gated Calcium Channels in Neurite Extension and Radial Migration. J. Neurosci. 2018, 38, 5551–5566. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, C.; Wong, R.O. Regulation of dendritic growth and plasticity by local and global calcium dynamics. Cell Calcium 2005, 37, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Bando, Y.; Irie, K.; Shimomura, T.; Umeshima, H.; Kushida, Y.; Kengaku, M.; Fujiyoshi, Y.; Hirano, T.; Tagawa, Y. Control of Spontaneous Ca2+ Transients Is Critical for Neuronal Maturation in the Developing Neocortex. Cereb. Cortex 2016, 26, 106–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horigane, S.I.; Ozawa, Y.; Yamada, H.; Takemoto-Kimura, S. Calcium signalling: A key regulator of neuronal migration. J. Biochem. 2019, 165, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Teng, E.M.; Summers, R.G., Jr.; Ming, G.L.; Gage, F.H. Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. J. Neurosci. 2006, 26, 3–11. [Google Scholar] [CrossRef]
- Hensch, T.K. Critical period regulation. Annu. Rev. Neurosci. 2004, 27, 549–579. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Yang, C.H.; Hsu, K.S.; Ming, G.L.; Song, H. A critical period for enhanced synaptic plasticity in newly generated neurons of the adult brain. Neuron 2007, 54, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Scott, B.W.; Wojtowicz, J.M. Heterogenous properties of dentate granule neurons in the adult rat. J. Neurobiol. 2000, 42, 248–257. [Google Scholar] [CrossRef]
- Schmidt-Hieber, C.; Jonas, P.; Bischofberger, J. Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Nature 2004, 429, 184–187. [Google Scholar] [CrossRef]
- Kempermann, G. What Is Adult Hippocampal Neurogenesis Good for? Front. Neurosci. 2022, 16, 852680. [Google Scholar] [CrossRef]
- Jennen, L.; Mazereel, V.; Lecei, A.; Samaey, C.; Vancampfort, D.; van Winkel, R. Exercise to spot the differences: A framework for the effect of exercise on hippocampal pattern separation in humans. Rev. Neurosci. 2022. [Google Scholar] [CrossRef]
- Stuchlik, A. Dynamic learning and memory, synaptic plasticity and neurogenesis: An update. Front. Behav. Neurosci. 2014, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anacker, C.; Hen, R. Adult hippocampal neurogenesis and cognitive flexibility-linking memory and mood. Nat. Rev. Neuroscience 2017, 18, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Aimone, J.B.; Wiles, J.; Gage, F.H. Potential role for adult neurogenesis in the encoding of time in new memories. Nat. Neurosci. 2006, 9, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Aimone, J.B.; Wiles, J.; Gage, F.H. Computational influence of adult neurogenesis on memory encoding. Neuron 2009, 61, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Piatti, V.C.; Davies-Sala, M.G.; Espósito, M.S.; Mongiat, L.A.; Trinchero, M.F.; Schinder, A.F. The timing for neuronal maturation in the adult hippocampus is modulated by local network activity. J. Neurosci. 2011, 31, 7715–7728. [Google Scholar] [CrossRef] [Green Version]
- Vivar, C.; Potter, M.C.; Choi, J.; Lee, J.Y.; Stringer, T.P.; Callaway, E.M.; Gage, F.H.; Suh, H.; van Praag, H. Monosynaptic inputs to new neurons in the dentate gyrus. Nat. Commun. 2012, 3, 1107. [Google Scholar] [CrossRef] [Green Version]
- Farmer, J.; Zhao, X.; van Praag, H.; Wodtke, K.; Gage, F.H.; Christie, B.R. Effects of voluntary exercise on synaptic plasticity and gene expression in the dentate gyrus of adult male Sprague-Dawley rats in vivo. Neuroscience 2004, 124, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Vaynman, S.; Ying, Z.; Gomez-Pinilla, F. Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur. J. Neurosci. 2004, 20, 2580–2590. [Google Scholar] [CrossRef]
- O’Callaghan, R.M.; Ohle, R.; Kelly, A.M. The effects of forced exercise on hippocampal plasticity in the rat: A comparison of LTP, spatial- and non-spatial learning. Behav. Brain Res. 2007, 176, 362–366. [Google Scholar] [CrossRef]
- Prakash, R.S.; Voss, M.W.; Erickson, K.I.; Kramer, A.F. Physical activity and cognitive vitality. Annu. Rev. Psychol. 2015, 66, 769–797. [Google Scholar] [CrossRef] [Green Version]
- Soya, H.; Nakamura, T.; Deocaris, C.C.; Kimpara, A.; Iimura, M.; Fujikawa, T.; Chang, H.; McEwen, B.S.; Nishijima, T. BDNF induction with mild exercise in the rat hippocampus. Biochem. Biophys. Res. Commun. 2007, 358, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.; Tavazoie, S.F.; Maloratsky, A.; Jacobs, K.M.; Harris, K.M.; Greenberg, M.E. CREB: A major mediator of neuronal neurotrophin responses. Neuron 1997, 19, 1031–1047. [Google Scholar] [CrossRef] [Green Version]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 237–258. [Google Scholar] [PubMed]
- Obrietan, K.; Gao, X.B.; Van Den Pol, A.N. Excitatory actions of GABA increase BDNF expression via a MAPK-CREB-dependent mechanism--a positive feedback circuit in developing neurons. J. Neurophysiol. 2002, 88, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; McCorkle, S.R.; Cha-Molstad, H.; Dwyer, J.M.; Yochum, G.S.; Boss, J.M.; McWeeney, S.; Dunn, J.J.; Mandel, G.; Goodman, R.H. Defining the CREB regulon: A genome-wide analysis of transcription factor regulatory regions. Cell 2004, 119, 1041–1054. [Google Scholar]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Merz, K.; Herold, S.; Lie, D.C. CREB in adult neurogenesis--master and partner in the development of adult-born neurons? Eur. J. Neurosci. 2011, 33, 1078–1086. [Google Scholar] [CrossRef]
- Magill, S.T.; Cambronne, X.A.; Luikart, B.W.; Lioy, D.T.; Leighton, B.H.; Westbrook, G.L.; Mandel, G.; Goodman, R.H. microRNA-132 regulates dendritic growth and arborization of newborn neurons in the adult hippocampus. Proc. Natl. Acad. Sci. USA 2010, 107, 20382–20387. [Google Scholar] [CrossRef] [Green Version]
- Luikart, B.W.; Bensen, A.L.; Washburn, E.K.; Perederiy, J.V.; Su, K.G.; Li, Y.; Kernie, S.G.; Parada, L.F.; Westbrook, G.L. miR-132 mediates the integration of newborn neurons into the adult dentate gyrus. PLoS ONE 2011, 6, e19077. [Google Scholar] [CrossRef]
- Mojtahedi, S.; Shabkhiz, F.; Ravasi, A.A.; Rosenkranz, S.; Soori, R.; Soleimani, M.; Tavakoli, R. Voluntary wheel running promotes improvements in biomarkers associated with neurogenic activity in adult male rats. Biochem. Biophys. Res. Commun. 2020, 533, 1505–1511. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Real, C.C.; Rodrigues, A.C.; Alves, A.S.; Britto, L.R. Short-term, moderate exercise is capable of inducing structural, BDNF-independent hippocampal plasticity. Brain Res. 2011, 1425, 111–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, M.G.; Bruce, S.R.; Kohman, R.A. Exercise Duration Differentially Effects Age-related Neuroinflammation and Hippocampal Neurogenesis. Neuroscience 2022, 490, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Cull-Candy, S.G.; Leszkiewicz, D.N. Role of distinct NMDA receptor subtypes at central synapses. Sci. STKE 2004, 255, re16. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, T.A.; Blackman, A.V.; Oyrer, J.; Jayabal, S.; Chung, A.J.; Watt, A.J.; Sjöström, P.J.; van Meyel, D.J. Neuronal morphometry directly from bitmap images. Nat. Methods 2014, 11, 982–984. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lattanzi, D.; Savelli, D.; Pagliarini, M.; Cuppini, R.; Ambrogini, P. Short-Term, Voluntary Exercise Affects Morpho-Functional Maturation of Adult-Generated Neurons in Rat Hippocampus. Int. J. Mol. Sci. 2022, 23, 6866. https://doi.org/10.3390/ijms23126866
Lattanzi D, Savelli D, Pagliarini M, Cuppini R, Ambrogini P. Short-Term, Voluntary Exercise Affects Morpho-Functional Maturation of Adult-Generated Neurons in Rat Hippocampus. International Journal of Molecular Sciences. 2022; 23(12):6866. https://doi.org/10.3390/ijms23126866
Chicago/Turabian StyleLattanzi, Davide, David Savelli, Marica Pagliarini, Riccardo Cuppini, and Patrizia Ambrogini. 2022. "Short-Term, Voluntary Exercise Affects Morpho-Functional Maturation of Adult-Generated Neurons in Rat Hippocampus" International Journal of Molecular Sciences 23, no. 12: 6866. https://doi.org/10.3390/ijms23126866