
Molecular Mechanisms of Phenylpropane-Synthesis-Related Genes Regulating the Shoot Blight Resistance of Bambusa pervariabilis × Dendrocalamopsis grandis


Abstract
:1. Introduction
2. Results
2.1. Gene Sequence Analysis and Protein Structure Prediction
2.2. B. pervariabilis × D. grands Total RNA Extraction and Reverse Transcription
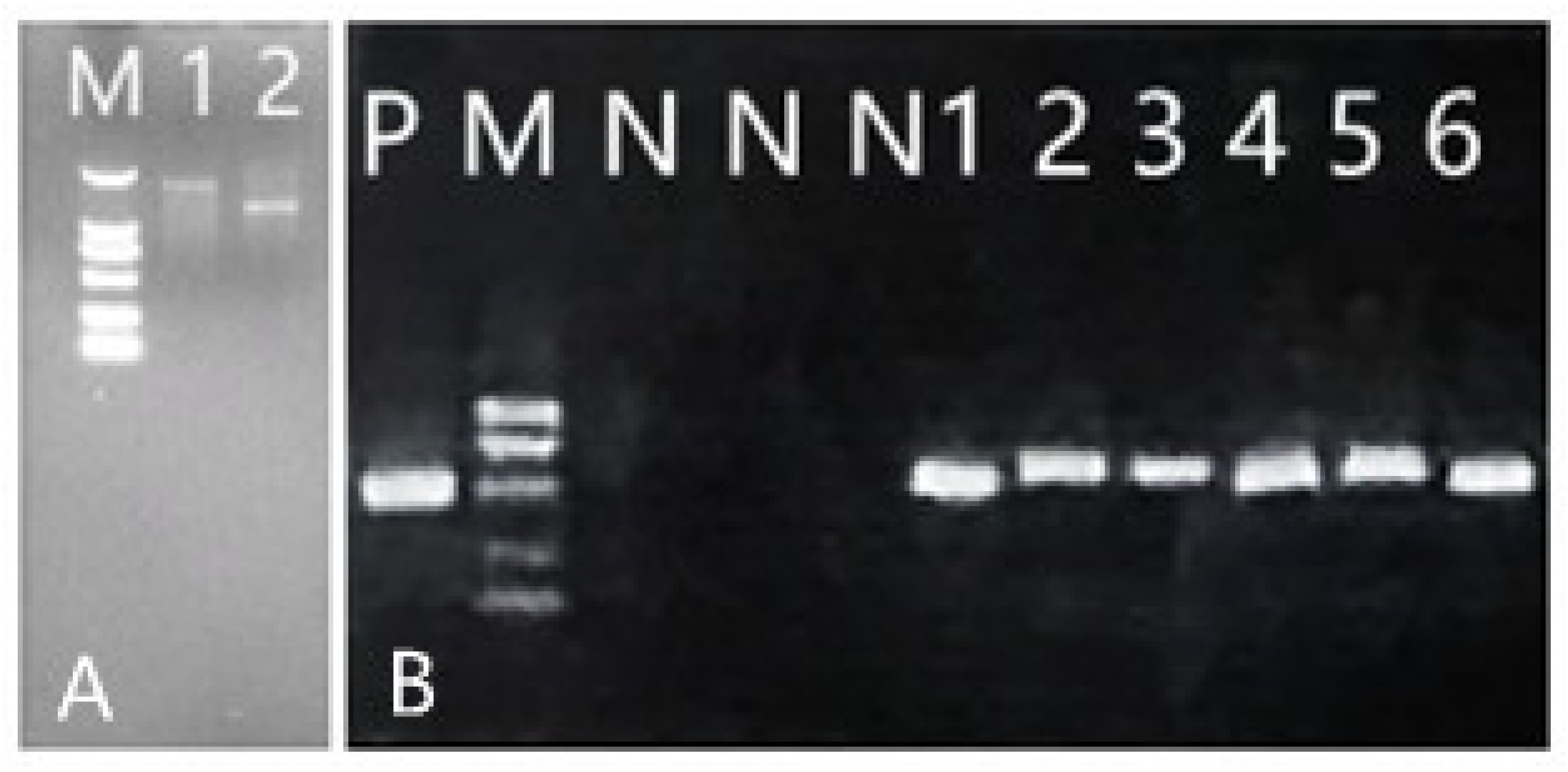
2.3. Construction and Transformation of RNAi Expression Plasmids of CCoAOMT2 and CAD5 Genes
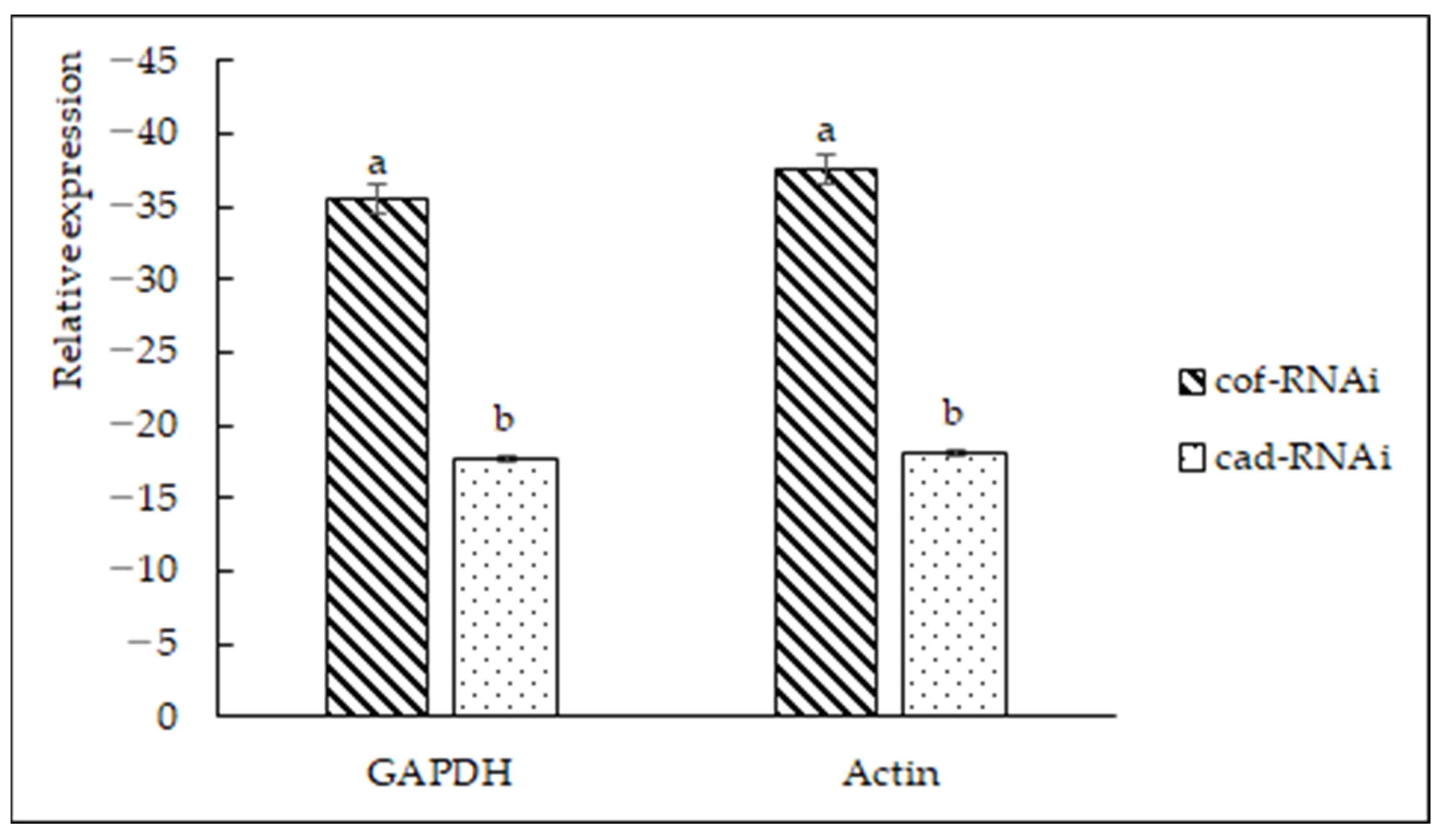
2.4. Expression Analysis of CCoAOMT2 Gene and CAD5 Gene
2.5. Disease Resistance Level of Transgenic Plants
2.6. Total Lignin Analysis and Total Flavonoids
2.6.1. Total Lignin Analysis
2.6.2. Total Flavonoids Analysis
2.6.3. Correlation Analysis
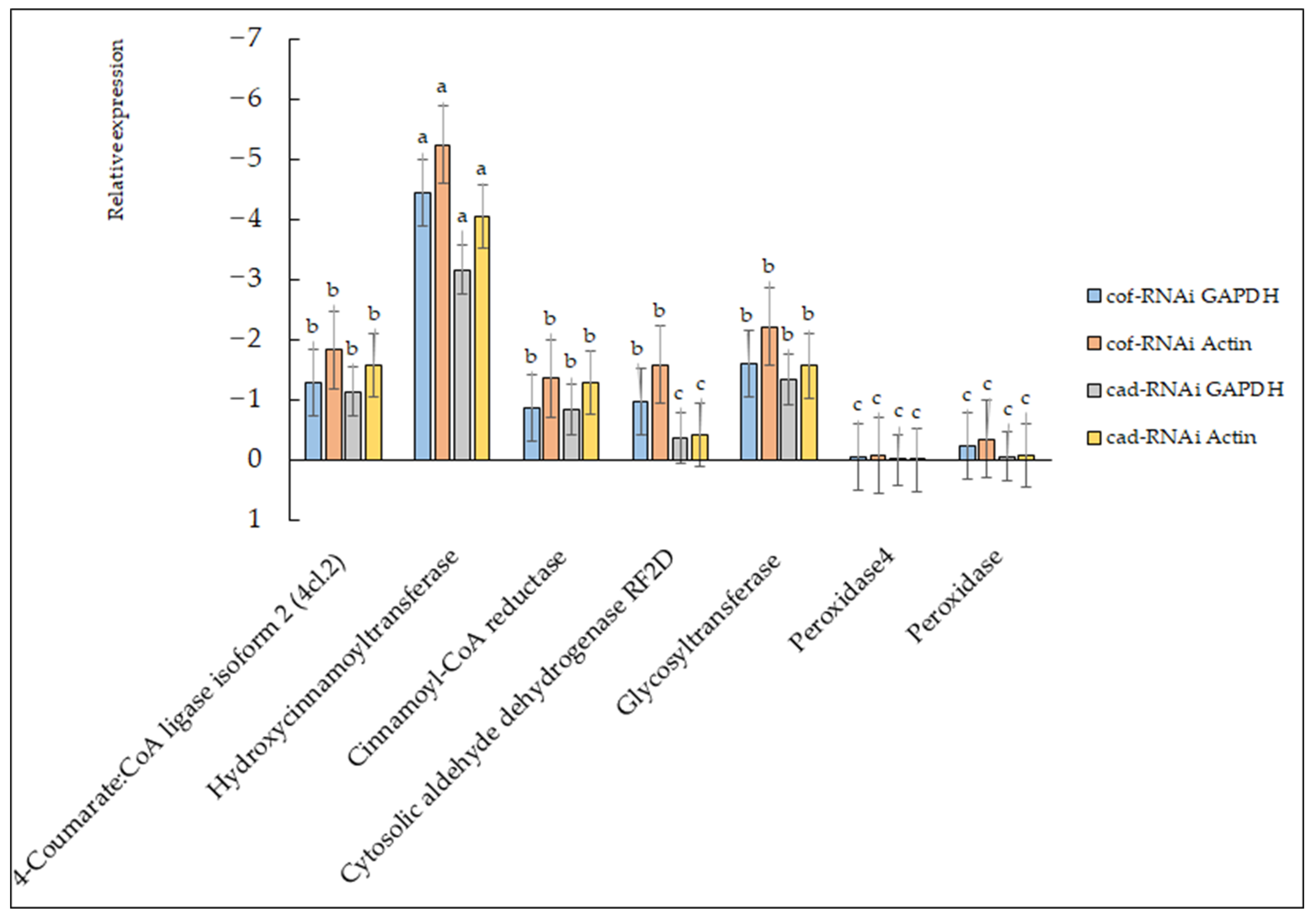
2.7. Gene Expression Analysis of Transformants
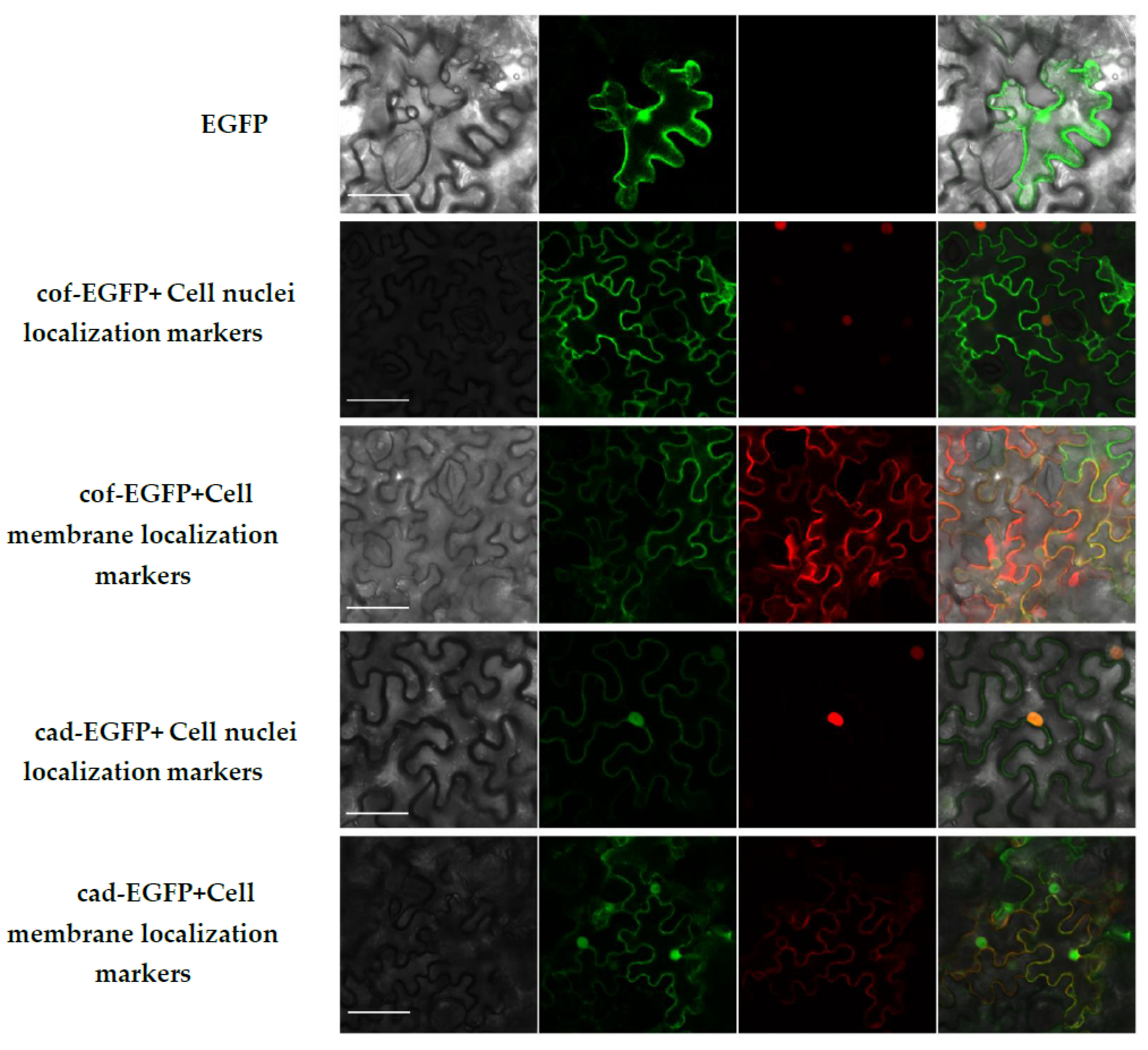
2.8. Subcellular Localization of CCoAOMT2 Gene and CAD5 Gene
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Gene Sequence Analysis, Protein Structure Prediction
4.2.2. Total RNA Extraction and cDNA Synthesis of B. pervariabilis × D. grandis
4.2.3. Construction and Transformation of RNAi Expression Plasmids of CCoAOMT2 and CAD5 Genes
4.2.4. Agrobacterium Transformation and B. pervariabilis × D. grandis Tissue Culture
4.2.5. Quantitative Detection of Gene Expression of CCoAOMT2 and CAD5 in RNAi Positive Transformed Vaccine by q-PCR
4.2.6. Pathogen Inoculation and Disease Resistance Detection
4.2.7. The Content of Total Lignin and Total Flavonoids
4.2.8. Expression Analysis of Related Genes
4.2.9. Subcellular Localization of CCoAOMT2 and CAD5 Gene
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, T.H. Investigation and spatial distribution of Bambusa pervariabilis × Dendrocalamopsis grandis. J. Sichuan For. Sci. Technol. 2009, 30, 44–48. [Google Scholar]
- Li, W.; Deng, Y.; Ning, Y.; He, Z.; Wang, G.-L. Exploiting broad-spectrum disease resistance in crops: From molecular dissection to breeding. Annu. Rev. Plant Biol. 2020, 71, 575–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Fang, X.; Han, S.; Zhu, T.; Zhu, H. Differential proteome analysis of hybrid Bamboo (Bambusa pervariabilis× Dendrocal-amopsis grandis) under fungal stress (Arthrinium phaeospermum). Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Luo, F.; Fang, X.; Liu, H.; Zhu, T.; Han, S.; Peng, Q.; Li, S. Differential transcriptome analysis and identification of genes related to resistance to blight in three varieties of Bambusa pervariabilis× Dendrocalamopsis grandis. Peer J. 2021, 9, e12301. [Google Scholar] [CrossRef]
- Li, C.L. Relationship between Shikimate Acid Pathway and Polyphenol Accumulation in Grape Berries. Mod. Agric. Sci. Technol. 2010, 12, 33–38. [Google Scholar]
- Geng, D.; Shen, X.; Xie, Y.; Yang, Y.; Bian, R.; Gao, Y.; Li, P.; Sun, L.; Feng, H.; Ma, F.; et al. Regulation of phenylpropanoid biosynthesis by MdMYB88 and MdMYB124 contributes to pathogen and drought resistance in apple. Hortic. Res. 2020, 7, 102. [Google Scholar] [CrossRef]
- Chen, J.-Y.; He, L.-H.; Jiang, Y.-M.; Kuang, J.-F.; Lu, C.-B.; Joyce, D.C.; Macnish, A.; He, Y.-X.; Lu, W.-J. Expression of PAL and HSPs in fresh-cut banana fruit. Environ. Exp. Bot. 2009, 66, 31–37. [Google Scholar] [CrossRef]
- Gao, X.H.; Bi, Y.; Wen, X.L.; Zheng, X.Y. Induction of postharvest malic acid treatment on activity of related enzymes and accumulation of final substances of benzene propance pathway in pears. J. Gansu Agric. Univ. 2009, 44, 132–136. [Google Scholar]
- Liu, J.Y.; Osbourn, A.; Ma, P.D. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kim, K.; Cho, M.; Franceschi, V.; Davin, L.; Lewis, N. Expression of cinnamyl alcohol dehydrogenases and their putative homologues during Arabidopsis thaliana growth and development: Lessons for database annotations? Phytochemistry 2007, 68, 1957–1974. [Google Scholar] [CrossRef]
- Gu, Z.J.; Zhang, D.Q.; Huang, Q.Y. Transgenic modification on pulp plants by key genes regulating lignin biosynthesis. J. Cent. South Univ. For. Technol. 2010, 30, 67–74. [Google Scholar]
- Liu, S.J.; Huang, Y.H.; He, C.J.; Fang, C.; Zhang, Y.W. Cloning, bioinformatics and transcriptional analysis of caffeoyl-coenzyme A 3-O-methyltransferase in switchgrass under abiotic stress. J. Integr. Agric. 2016, 15, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, J.; Fu, C. Comparative physiology and transcriptome analysis reveals the regulatory mechanism of genome duplication enhancing cold resistance in Fragaria nilgerrensis. Environ. Exp. Bot. 2021, 188, 104509. [Google Scholar] [CrossRef]
- Chen, C.; Wang, C.; Liu, Z.; Cai, Z.; Hua, Y.; Mei, Y.; Wei, L.; Liu, X. iTRAQ-based proteomic technique provides insights into salt stress responsive proteins in Apocyni Veneti Folium (Apocynum venetum L.). Environ. Exp. Bot. 2020, 180, 104247. [Google Scholar] [CrossRef]
- Ge, Q.; Kuang, J.; Wu, Y.-C.; Zhang, Y.; Xiao, Y.-P.; Wang, Z.-Z. Cloning and Expression Analysis of Cinnamyl Alcohol Dehydrogenase (SmCAD) Gene in Salvia miltiorrhiza Bunge. Plant Sci. J. 2013, 31, 261–268. [Google Scholar] [CrossRef]
- Liu, H.J.; Yue-Peng, L.I.; Liu, W.; Shao, Q.; Hong-Yan, Q.I. Effects of Fusarium oxysporum f. sp. melonis on Lignin, Activities of Lingin-Related Enzymes and Genes Expressions of CmCADs in Oriental Melon (Cucumis melo var. makuwa Makino). Sci. Agric. Sin. 2016, 49, 2153–2163. [Google Scholar]
- Somegowda, M.; Vigneshwaran, V.; Sunil, S.V.; Shivashankar, S.; Babu, H.R.; Pramod, S.; Rajeshwar, A.N. Comparison of Cellular Antioxidants, Enzyme Activities in Resistance and Susceptible Cucurbits Fruits Response to Melon Fly Infestation to Elucidate Possible Mechanism. Der Pharma Chem. 2018, 10, 55–64. [Google Scholar]
- Zhang, C.Q. Functional Research of Cinnamyl Alcohol Dehydrogenase in Lignin of Bambusa multiplex (Lour.). Master’s Thesis, Anhui Agricultural University, Hefei, China, 2013. [Google Scholar]
- Wassenegger, M.; Pelissier, T. A model for RNA-mediated gene silencing in higher plants. Plant Mol. Biol. 1998, 37, 349–362. [Google Scholar] [CrossRef]
- Zhao, Z.; Cao, Y.; Li, M.; Meng, A. Double-stranded RNA injection produces nonspecific defects in zebrafish. Dev. Biol. 2001, 229, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Kennerdell, J.R.; Carthew, R.W. Heritable gene silencing in Drosophila using double-stranded RNA. Nat. Biotechnol. 2000, 18, 896–898. [Google Scholar] [CrossRef]
- Cogoni, C.; Macino, G. Homology-dependent gene silencing in plants and fungi: A number of variations on the same theme. Curr. Opin. Microbiol. 1999, 2, 657–662. [Google Scholar] [CrossRef]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, P.; Stein, P.; Hayashi, H.; Schultz, R.M. Selective reduction of dormant maternal mRNAs in mouse oocytes by RNA interference. Development 2000, 127, 4147–4156. [Google Scholar] [CrossRef]
- Aly, R.; Cholakh, H.; Joel, D.; Leibman, D.; Steinitz, B.; Zelcer, A.; Naglis, A.; Yarden, O.; Gal-On, A. Gene silencing of mannose 6-phosphate reductase in the parasitic weed Orobanche aegyptiaca through the production of homologous dsRNA sequences in the host plant. Plant Biotechnol. J. 2009, 7, 487–498. [Google Scholar] [CrossRef]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the Mechanism of Action of Double-Stranded RNA Activity against Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef]
- Mumbanza, F.M.; Kiggundu, A.; Tusiime, G.; Tushemereirwe, W.K.; Niblett, C.; Bailey, A. In vitro antifungal activity of synthetic dsRNA molecules against two pathogens of banana, Fusarium oxysporum f. sp cubense and Mycosphaerella fijiensis. Pest Manag. Sci. 2013, 69, 1155–1162. [Google Scholar] [CrossRef]
- Sindhu, A.S.; Maier, T.R.; Mitchum, M.G.; Hussey, R.S.; Davis, E.L.; Baum, T.J. Effective and specific in planta RNAi in cyst nematodes: Expression interference of four parasitism genes reduces parasitic success. J. Exp. Bot. 2009, 60, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.L.; Gu, X.P.; Li, L.B.; Yue, J.J.; Yao, N.; Guo, G.P. Callus Induction and Plantlet Regeneration of Bambusa multiplex. Sci. Silvae Sin. 2009, 45, 34–39. [Google Scholar]
- Chen, L.; Guo, X.; Yang, H. Cloning and Expression Analysis of CCoAOMT, a Key Gene in Lignin Biosynthesis of Dendrocalamus sinicus. J. Plant Genet. Resour. 2019, 20, 467–484. [Google Scholar]
- Zhu, J.; Sun, J.; Yuan, X.; Wang, J.; Yang, Y.; Wang, Y. Cloning and Sequence Analysis of Cinnamoyl Alcohol Dehydrogenase Gene from Guadua paraguayana. J. West China For. Sci. 2017, 46, 6–11. [Google Scholar]
- Ferrer, J.; Zubieta, C.; Dixon, R.; Noel, J. Crystal structures of alfalfa caffeoyl coenzyme A 3-O-methyltransferase. Plant Physiol. 2005, 137, 1009–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.W. Cloning of Key Genes Involved in Lignin Monomer Biosynthesis from Eucalvptus urophylla and Transformation in Tobacco. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2017. [Google Scholar]
- Luo, B.; Han, Y.X.; Zhang, X.Q.; Guo, H.; Li, H.M.; Wang, J.; Yang, Z.G.; Sun, H.Y. Construction and Genetic Transformation of RNAi Expression Vector Targ-eting UDP-N-acetylglucosamine Acyltransferase (OsLpxA) Gene in Rice. Mol. Plant Breed. 2018, 16, 5311–5317. [Google Scholar]
- Cao, Y.H.; Zhang, Z.; Yao, Q.H.; Peng, R.H.; Xiong, A.S.; Li, X. Study on double stranded RNA interference (RNAi) of apple polyphenol oxidase. Acta Biol. Exp. Sinioa 2004, 37, 487–493. [Google Scholar]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.Y.; Sun, X.J.; Liu, Z.Y. A Research Review of RNA Interference Mechanisms and Its Applications in Plants. J. Jilin Agric. Univ. 2013, 35, 160–166. [Google Scholar]
- Jin, L.; Sun, Z.Y.; Liu, Y.; Tian-Hong, L.I. Cloning of Carnation GA20-oxidase Gene and Construction of Plant RNAi Vector. J. Agric. Biotechnol. 2009, 17, 107–113. [Google Scholar]
- Li, T.T. Identification and Functional Analysis of Conserved Target CAD7 by Phytophthora Avr3 Family Effectors. Ph.D. Thesis, Northwest University of Agriculture and Forestry Science and Technology, Xianyang, China, 2017. [Google Scholar]
- Li, Z. Genetic manipulation of Lignin Biosynthesis in Poplar. Master’s Thesis, Capital Normal University, Beijing, China, 2009. [Google Scholar]
- Zhou, J.; Lee, E.; Kanapathy-Sinnaiaha, F.; Park, Y.; Kornblatt, J.; Lim, Y.; Ibrahim, R. Structure-function relationships of wheat flavone O-methyltransferase: Homology modeling and site-directed mutagenesis. BMC Plant Biol. 2010, 10, e156. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Zhou, R.; Louie, G.; Mühlemann, J.; Bomati, E.; Bowman, M.; Dudareva, N.; Dixon, R.; Noel, J.; Wang, X. Structural studies of cinnamoyl-CoA reductase and cinnamyl-alcohol dehydrogenase, key enzymes of monolignol biosynthesis. Plant Cell 2014, 26, 3709–3727. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Cheng, X.; Leshkevich, J.; Umezawa, T.; Harding, S.; Chiang, V. The last step of syringyl monolignol biosynthesis in angiosperms is regulated by a novel gene encoding sinapyl alcohol dehydrogenase. Plant Cell 2001, 13, 1567–1586. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Chen, F.; Inoue, K.; Blount, J.; Dixon, R. Downregulation of caffeic acid 3-O-methyltransferase and caffeoyl CoA 3-O-methyltransferase in transgenic alfalfa: Impacts on lignin structure and implications for the biosynthesis of G and S lignin. Plant Cell 2001, 13, 73–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, N.; Liu, S.; Liu, B.; Li, N.; Jiang, X.; Gai, Y. Molecular cloning and functional analysis of nine cinnamyl alcohol dehydrogenase family members in Populus tomentosa. Planta 2014, 240, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Bagniewska-Zadworna, A.; Barakat, A.; Lakomy, P.; Smolinski, D.J.; Zadworny, M. Lignin and lignans in plant defence: Insight from expression profiling of cinnamyl alcohol dehydrogenase genes during development and following fungal infection in Populus. Plant Sci. 2014, 229, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.Y.; Qiao, J.J.; Yan-Ni, L.I. Structure, Function and Application of Hydroxycinnamoyl Transferase. Chin. J. Biochem. Mol. Biol. 2019, 35, 1058–1066. [Google Scholar]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of hydroxycinnamoyl-coenzyme A shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [Green Version]
- Ruelland, E.; Campalans, A.; Selman-Housein, G.; Puigdomènech, P.; Rigau, J. Cellular and subcellular localization of the lignin biosynthetic enzymes caffeic acid-O-methyltransferase, cinnamyl alcohol dehydrogenase and cinnamoyl-coenzyme A reductase in two monocots, sugarcane and maize. Physiol. Plant. 2003, 117, 93–99. [Google Scholar] [CrossRef]
- Tang, R.; Zhang, B.; Peng, X.; Zhang, X.; Yuan, J.; Xie, X. Subcellular Localization of PpCAD from Elephant Grass (Pennisetum purpureum cv.Huanan) and Its Heterologous Expression in Transgenic Tobacco. Acta Bot. Boreali-Occident. Sin. 2015, 35, 1069–1077. [Google Scholar]
- Nakashima, J.; Awano, T.; Takabe, K.; Fujita, M.; Saiki, H. Immunocytochemical Localization of Phenylalanine Ammonia-Lyase and Cinnamyl Alcohol Dehydrogenase in Differentiating Tracheary Elements Derived from Zinnia Mesophyll Cells. Plant Cell Physiol. 1997, 38, 113–123. [Google Scholar] [CrossRef]
- Ran, X. The Development of Enzymology Studies in the Metabolism of Lignin Biosynthesis. Chin. Agric. Sci. Bull. 2009, 25, 23–27. [Google Scholar]
- Xiaoyan, L.; Wang, Y.; Jin, Y.; University, H. Cloning, expression and bioinformatics analyses of BvM14-CCoAOMT gene. J. Nat. Sci. Heilongjiang Univ. 2018, 35, 317–323. [Google Scholar]
- Zhang, Q.; Gong, X.; Zhang, T.Z.; Chen, Y.; Wang, H.; Tao, S.T.; Bao, J.P. Subcellular Localization and Expression Analysis of PsiCAD1 and PsiPRX6 Genes in Kuerle Pear. Mol. Plant Breed. 2022; accepted. [Google Scholar]
- Liu, P.P. Effects of Boron on Lignin Metabolism Related Enzymes and Stone Cell Formation in Pear. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Fang, Z.D. Plant Disease Research Methods, 3rd ed.; China Agricultural Press: Beijing, China, 1998. [Google Scholar]
- Molecular Evolutionary Genetics Analysis. Available online: https://megasoftware.net/ (accessed on 16 May 2022).
- Protein Conserved Domain Analysis. Available online: https://meme-suite.org/meme/tools/meme (accessed on 16 May 2022).
- Protein Primary Structure Analysis. Available online: https://www.expasy.org/ (accessed on 16 May 2022).
- Protein Tertiary Structure Analysis. Available online: https://swissmodel.expasy.org/ (accessed on 16 May 2022).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Luo, D.; He, S.; Liu, F. Comparative study on four inoculation methods of rice bacterial leaf streak. Subtrop. Agric. Res. 2018, 14, 242–246. [Google Scholar]
- Liu, W.; Zhu, D.-W.; Liu, D.-H.; Geng, M.-J.; Zhou, W.-B.; Mi, W.-J.; Yang, T.-W.; Hamilton, D. Influence of nitrogen on the primary and secondary metabolism and synthesis of flavonoids in c hrysanthemum morifolium ramat. J. Plant Nutr. 2010, 33, 240–254. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, K.; Li, S.; Li, J.; Huang, J.; Chen, R.; Pang, S.; Zhou, X. Genome-wide analysis of the NAAT, DMAS, TOM, and ENA gene families in maize suggests their roles in mediating iron homeostasis. BMC Plant Biol. 2022, 22, 1–16. [Google Scholar] [CrossRef]
- ProteomeXchange Consortium Protein Database. Available online: http://proteomecentral.proteomexchange.org (accessed on 16 May 2022).
- Ma, J.; Chen, T.; Wu, S.; Yang, C.; Bai, M.; Shu, K.; Li, K.; Zhang, G.; Jin, Z.; He, F. iProX: An integrated proteome resource. Nucleic Acids Res. 2019, 47, D1211–D1217. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Incidence (%) | Disease Index |
|---|---|---|
| cof-RNAi | 86.7 ± 4.7 a | 42.5 ± 2.0 a |
| cad-RNAi | 83.3 ± 9.4 a | 55.8 ± 5.1 a |
| wild-type B. pervariabilis × D. grandis | 16.7 ± 4.7 b | 9.2 ± 3.1 b |
| Varieties | Lignin Content mg/g |
|---|---|
| wild-type B. pervariabilis × D. grandis | 4.064 ± 0.107 a |
| cof-RNAi | 2.727 ± 0.473 b |
| cad-RNAi | 3.152 ± 0.449 b |
| Varieties | Flavonoids Content mg/g |
|---|---|
| wild-type B. pervariabilis × D. grandis | 5.63 ± 1.31 a |
| cof-RNAi | 3.84 ± 0.26 b |
| cad-RNAi | 3.01 ± 0.24 b |
| Incidence | Disease Index | Lignin Content | Flavonoids Content | |
|---|---|---|---|---|
| CCoAOMT2 relative expression | −0.9865 ** | −0.9840 ** | 0.8849 * | 0.9992 ** |
| CAD5 relative expression | −0.9810 ** | −0.9877 ** | 0.8270 * | 0.9991 ** |
| lignin content | −0.7917 * | −0.7173 * | / | 0.6515 |
| flavonoids content | −0.9365 * | −0.9994 * | 0.6515 | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, F.; Yan, P.; Xie, L.; Li, S.; Zhu, T.; Han, S.; Lin, T.; Li, S. Molecular Mechanisms of Phenylpropane-Synthesis-Related Genes Regulating the Shoot Blight Resistance of Bambusa pervariabilis × Dendrocalamopsis grandis. Int. J. Mol. Sci. 2022, 23, 6760. https://doi.org/10.3390/ijms23126760
Luo F, Yan P, Xie L, Li S, Zhu T, Han S, Lin T, Li S. Molecular Mechanisms of Phenylpropane-Synthesis-Related Genes Regulating the Shoot Blight Resistance of Bambusa pervariabilis × Dendrocalamopsis grandis. International Journal of Molecular Sciences. 2022; 23(12):6760. https://doi.org/10.3390/ijms23126760
Chicago/Turabian StyleLuo, Fengying, Peng Yan, Liling Xie, Shuying Li, Tianhui Zhu, Shan Han, Tiantian Lin, and Shujiang Li. 2022. "Molecular Mechanisms of Phenylpropane-Synthesis-Related Genes Regulating the Shoot Blight Resistance of Bambusa pervariabilis × Dendrocalamopsis grandis" International Journal of Molecular Sciences 23, no. 12: 6760. https://doi.org/10.3390/ijms23126760