
The Periventricular Nucleus as a Brain Center Containing Dopaminergic Neurons and Neurons Expressing Individual Enzymes of Dopamine Synthesis

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
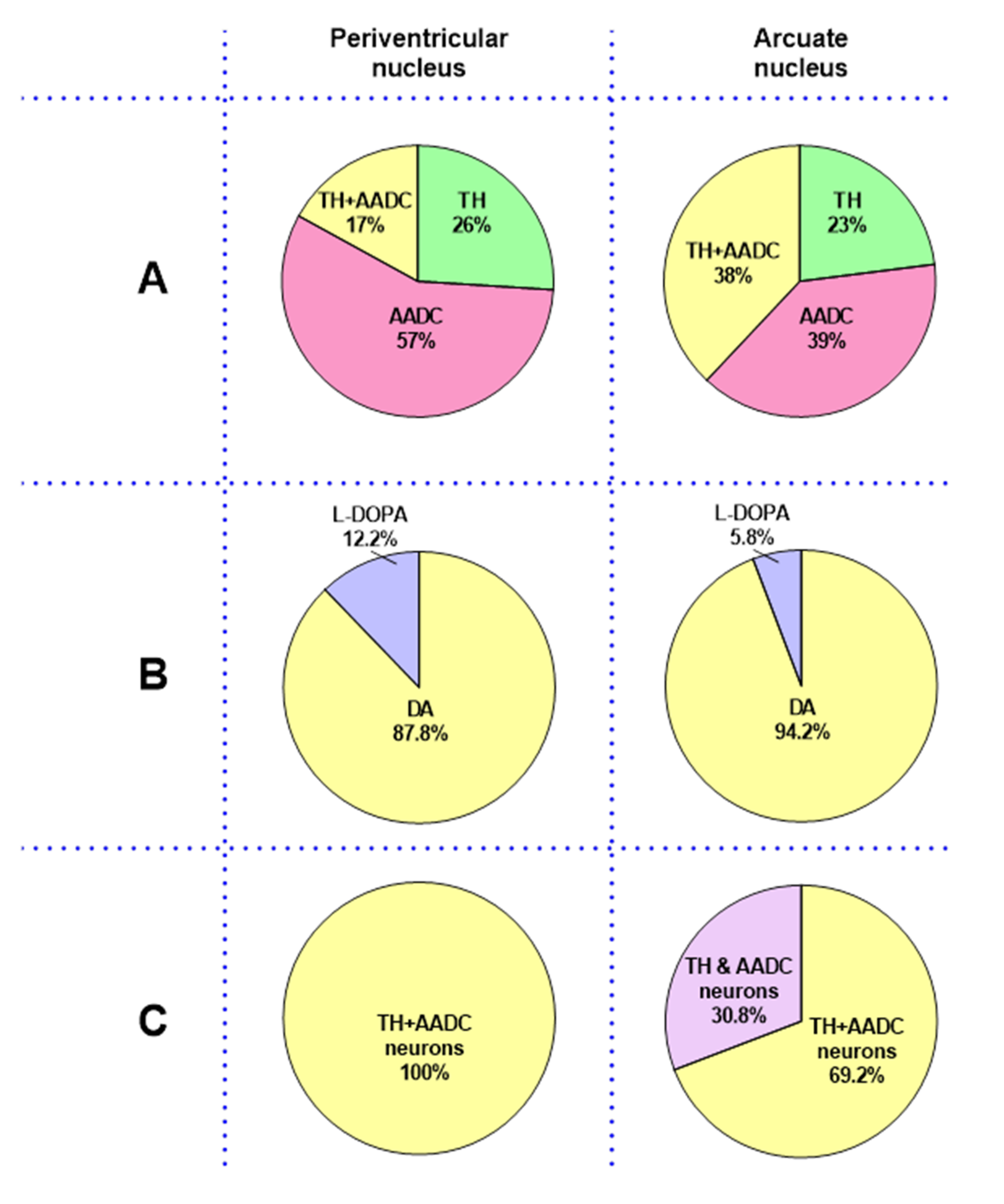
2.1. Dopamine and L-DOPA Levels in the Cerebrospinal Fluid and in the Periventricular and Arcuate Nuclei
2.2. Dopamine Content in Vibratome Sections of the Periventricular Nucleus, Arcuate Nucleus, and Substantia Nigra and in the Incubation Medium after Incubating the Sections with 2-Amino-2-Norbornanecarboxylic Acid, as Well as in the Control
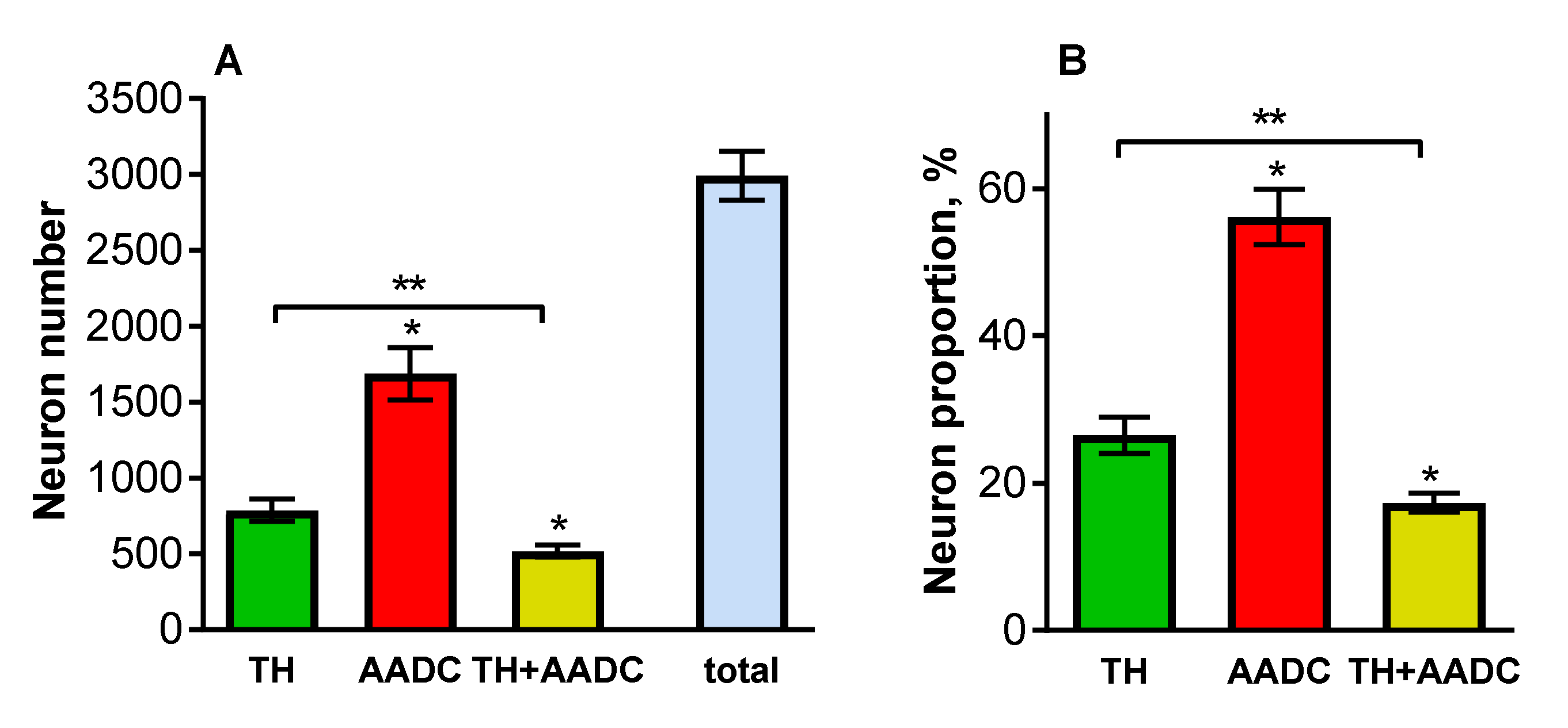
2.3. Neurons That Are Immunopositive for Tyrosine Hydroxylase, Aromatic L-Amino Acid Decarboxylase, and Both Enzymes in the Periventricular Nucleus
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experiments and Obtaining Samples for Analysis
4.2.1. Cerebrospinal Fluid Collection
4.2.2. Preparation of Vibratome Brain Sections and Their Incubation
4.2.3. Obtaining Samples of Periventricular and Arcuate Nucleuses for Biochemical Analysis
4.2.4. Preparing the Brain for Immunohistochemistry
4.3. Methods
4.3.1. High-Performance Liquid Chromatography
4.3.2. Immunohistochemistry of Sections on Slides
4.3.3. Immunohistochemistry of Floating Sections
4.3.4. Microscopy of Periventricular Nucleus Thin Sections on Slides and Quantification of Neurons Expressing Dopamine Synthesis Enzymes
4.3.5. Confocal Microscopy of the Periventricular Nucleus, Thick Floating Sections and 3D Analysis of the Neuron Topography: Cell Bodies and Processes Containing Dopamine Synthesis Enzymes
4.4. Statistical Analysis
5. Conclusions
- (1)
- The PeVN of the hypothalamus is one of the powerful dopaminergic centers in the brain, which contains numerous neurons expressing both or individual complementary enzymes of dopamine synthesis;
- (2)
- L-DOPA and dopamine are produced in the PeVN as the final synthetic products, most probably in monoenzymatic TH neurons and monoenzymatic AADC neurons and/or bienzymatic neurons;
- (3)
- Bienzymatic and monoenzymatic neurons (cell bodies and fibers) are topographically closely related to each other, as well as to the third ventricle, providing L-DOPA and dopamine delivery pathways to the CSF.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AADC | aromatic L-amino acid decarboxylase |
| AN | arcuate nucleus |
| BCH | 2-amino-2-norbornanecarboxylic acid |
| CSF | cerebrospinal fluid |
| HPLC | high performance liquid chromatography |
| KRS | Krebs-Ringer solution |
| LAT1 | L-type amino acid transporter 1 |
| PD | Parkinson’s disease |
| PeVN | periventricular nucleus |
| TH | tyrosine hydroxylase |
| TPH | tryptophan hydroxylase |
References
- Dahlstroem, A.; Fuxe, K. Evidence for the Existence of Monoamine-Containing Neurons in the Central Nervous System. Acta Physiol. Scand. Suppl. 1964, SUPPL 232, 1–55. [Google Scholar]
- Hökfelt, T.; Johansson, O.; Goldstein, M. Central Catecholamine Neurons as Revealed by Immunocytochemistry with Special Reference to Adrenaline Neurons. In Handbook of Chemical Neuroanatomy (Classical Nuerotransmitters in the CNS, Part 1: Vol. 2); Björklund, A., Hökfelt, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1984; pp. 157–276. [Google Scholar]
- Fuxe, K.; Dahlström, A.B.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The Discovery of Central Monoamine Neurons Gave Volume Transmission to the Wired Brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Palkovits, M.; Brownstein, M.J. Catecholamines in the Central Nervous System. In Handbook of chemical neuroanatomy (Catecholamines: Vol. 90/2); Trendelenburg, U., Winer, N., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 1–26. [Google Scholar]
- Meister, B.; Hökfelt, T.; Steinbusch, H.W.; Skagerberg, G.; Lindvall, O.; Geffard, M.; Joh, T.H.; Cuello, A.C.; Goldstein, M. Do Tyrosine Hydroxylase-Immunoreactive Neurons in the Ventrolateral Arcuate Nucleus Produce Dopamine or Only L-Dopa? J. Chem. Neuroanat. 1988, 1, 59–64. [Google Scholar] [PubMed]
- Ikemoto, K.; Kitahama, K.; Nishimura, A.; Jouvet, A.; Nishi, K.; Arai, R.; Jouvet, M.; Nagatsu, I. Tyrosine Hydroxylase and Aromatic L-Amino Acid Decarboxylase Do Not Coexist in Neurons in the Human Anterior Cingulate Cortex. Neurosci. Lett. 1999, 269, 37–40. [Google Scholar] [CrossRef]
- Ahmed, E.I.; Northcutt, K.V.; Lonstein, J.S. L-Amino Acid Decarboxylase- and Tyrosine Hydroxylase-Immunoreactive Cells in the Extended Olfactory Amygdala and Elsewhere in the Adult Prairie Vole Brain. J. Chem. Neuroanat. 2012, 43, 76–85. [Google Scholar] [CrossRef]
- Ugrumov, M.V. Non-Dopaminergic Neurons Partly Expressing Dopaminergic Phenotype: Distribution in the Brain, Development and Functional Significance. J. Chem. Neuroanat. 2009, 38, 241–256. [Google Scholar] [CrossRef]
- Ugrumov, M.V. Brain Neurons Partly Expressing Dopaminergic Phenotype. Location, Development, Functional Significance, and Regulation. In Advances in Pharmacology; 2013; Volume 68, pp. 37–91. [Google Scholar]
- Okamura, H.; Kitahama, K.; Mons, N.; Ibata, Y.; Jouvet, M.; Geffard, M. L-DOPA-Immunoreactive Neurons in the Rat Hypothalamic Tuberal Region. Neurosci. Lett. 1988, 95, 42–46. [Google Scholar] [CrossRef]
- Komori, K.; Fujii, T.; Nagatsu, I. Do Some Tyrosine Hydroxylase-Immunoreactive Neurons in the Human Ventrolateral Arcuate Nucleus and Globus Pallidus Produce Only l-DOPA? Neurosci. Lett. 1991, 133, 203–206. [Google Scholar] [CrossRef]
- Clarkson, J.; Herbison, A.E. Dual Phenotype Kisspeptin-Dopamine Neurones of the Rostral Periventricular Area of the Third Ventricle Project to Gonadotrophin-Releasing Hormone Neurones. J. Neuroendocrinol. 2011, 23, 293–301. [Google Scholar] [CrossRef]
- Stephens, S.B.Z.; Rouse, M.L.; Tolson, K.P.; Liaw, R.B.; Parra, R.A.; Chahal, N.; Kauffman, A.S. Effects of Selective Deletion of Tyrosine Hydroxylase from Kisspeptin Cells on Puberty and Reproduction in Male and Female Mice. eNeuro 2017, 4, ENEURO.0150-17.2017. [Google Scholar] [CrossRef] [Green Version]
- Kamkrathok, B.; Chaiseha, Y. Distribution of Dopamine-Immunoreactive Neurons in the Brain of the Male Native Thai Chicken. Folia Histochem. Cytobiol. 2022, 60, 1–12. [Google Scholar] [CrossRef]
- Ershov, P.V.; Ugrumov, M.V.; Calas, A.; Krieger, M.; Thibault, J. Differentiation of Tyrosine Hydroxylase-Synthesizing and/or Aromatic L-Amino Acid Decarboxylase-Synthesizing Neurons in the Rat Mediobasal Hypothalamus: Quantitative Double-Immunofluorescence Study. J. Comp. Neurol. 2002, 446, 114–122. [Google Scholar] [CrossRef]
- Ugrumov, M.V.; Melnikova, V.I.; Lavrentyeva, A.V.; Kudrin, V.S.; Rayevsky, K.S. Dopamine Synthesis by Non-Dopaminergic Neurons Expressing Individual Complementary Enzymes of the Dopamine Synthetic Pathway in the Arcuate Nucleus of Fetal Rats. Neuroscience 2004, 124, 629–635. [Google Scholar] [CrossRef]
- Ugrumov, M.; Taxi, J.; Pronina, T.; Kurina, A.; Sorokin, A.; Sapronova, A.; Calas, A. Neurons Expressing Individual Enzymes of Dopamine Synthesis in the Mediobasal Hypothalamus of Adult Rats: Functional Significance and Topographic Interrelations. Neuroscience 2014, 277, 45–54. [Google Scholar] [CrossRef]
- Goshima, Y.; Nakamura, F.; Masukawa, D.; Chen, S.; Koga, M. The Cardiovascular Actions of DOPA Mediated by the Gene Product of Ocular Albinism 1. J. Pharmacol. Sci. 2014, 126, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Goshima, Y.; Masukawa, D.; Kasahara, Y.; Hashimoto, T.; Aladeokin, A.C. L-DOPA and Its Receptor GPR143: Implications for Pathogenesis and Therapy in Parkinson’s Disease. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, N.; Naito, S.; Masukawa, D.; Kaneda, M.; Miyamoto, H.; Abe, T.; Yamashita, Y.; Endo, I.; Nakamura, F.; Goshima, Y. Expression of Ocular Albinism 1 (OA1), 3, 4- Dihydroxy- L-Phenylalanine (DOPA) Receptor, in Both Neuronal and Non-Neuronal Organs. Brain Res. 2015, 1602, 62–74. [Google Scholar] [CrossRef]
- Masukawa, D.; Nishizawa, D.; Kanai, K.; Kitamura, S.; Kasahara, Y.; Hashimoto, T.; Takahagi, R.; Hasegawa, J.; Nakayama, K.; Sato, N.; et al. Genetic Associations of Single Nucleotide Polymorphisms in the L-DOPA Receptor (GPR143) Gene with Severity of Nicotine Dependence in Japanese Individuals, and Attenuation of Nicotine Reinforcement in Gpr143 Gene-Deficient Mice. J. Pharmacol. Sci. 2020, 144, 89–93. [Google Scholar] [CrossRef]
- Ershov, P.V.; Ugrumov, M.V.; Calas, A.; Krieger, M.; Thibault, J. Degeneration of Dopaminergic Neurons Triggers an Expression of Individual Enzymes of Dopamine Synthesis in Non-Dopaminergic Neurons of the Arcuate Nucleus in Adult Rats. J. Chem. Neuroanat. 2005, 30, 27–33. [Google Scholar] [CrossRef]
- Tepper, J.M.; Koós, T.; Ibanez-Sandoval, O.; Tecuapetla, F.; Faust, T.W.; Assous, M. Heterogeneity and Diversity of Striatal GABAergic Interneurons: Update 2018. Front. Neuroanat. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Ikemoto, K.; Nagatsu, I.; Nishimura, A.; Nishi, K.; Arai, R. Do All of Human Midbrain Tyrosine Hydroxylase Neurons Synthesize Dopamine? Brain Res. 1998, 805, 255–258. [Google Scholar] [CrossRef]
- Karasawa, N.; Hayashi, M.; Yamada, K.; Nagatsu, I.; Iwasa, M.; Takeuchi, T.; Uematsu, M.; Watanabe, K.; Onozuka, M. Tyrosine hydroxylase (TH)- and aromatic-L-amino acid decarboxylase (AADC)-immunoreactive neurons of the common marmoset (Callithrix jacchus) brain: An immunohistochemical analysis. Acta Histochem Cytochem. 2017, 40, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, C.B.; Ruggiero, D.A.; Albert, V.R.; Park, D.H.; Joh, T.H.; Reis, D.J. Aromatic L-amino acid decarboxylase in the rat brain: Immunocytochemical localization in neurons of the brain stem. Neuroscience 1984, 11, 691–713. [Google Scholar] [CrossRef]
- Nagatsu, I.; Sakai, M.; Yoshida, M.; Nagatsu, T. Aromatic L-Amino Acid Decarboxylase-Immunoreactive Neurons in and around the Cerebrospinal Fluid-Contacting Neurons of the Central Canal Do Not Contain Dopamine or Serotonin in the Mouse and Rat Spinal Cord. Brain Res. 1988, 475, 91–102. [Google Scholar] [CrossRef]
- Zhang, M. Two-Step Production of Monoamines in Monoenzymatic Cells in the Spinal Cord: A Different Control Strategy of Neurotransmitter Supply? Neural Regen. Res. 2016, 11, 1904–1909. [Google Scholar] [CrossRef]
- Kitahama, K.; Ikemoto, K.; Jouvet, A.; Nagatsu, I.; Sakamoto, N.; Pearson, J. Aromatic L-Amino Acid Decarboxylase- and Tyrosine Hydroxylase- Immunohistochemistry in the Adult Human Hypothalamus. J. Chem. Neuroanat. 1998, 16, 43–55. [Google Scholar] [CrossRef]
- Lehman, M.N.; Hileman, S.M.; Goodman, R.L. Neuroanatomy of the Kisspeptin Signaling System in Mammals: Comparative and Developmental Aspects. Adv. Exp. Med. Biol. 2013, 784, 27–62. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.H.; Colledge, W.H. The Role of Kiss1 Neurons as Integrators of Endocrine, Metabolic, and Environmental Factors in the Hypothalamic-Pituitary-Gonadal Axis. Front. Endocrinol. (Lausanne) 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Navarro, V.M. Metabolic Regulation of Kisspeptin — the Link between Energy Balance and Reproduction. Nat. Rev. Endocrinol. 2020, 16, 407–420. [Google Scholar] [CrossRef]
- Negrón, A.L.; Yu, G.; Boehm, U.; Acosta-Martínez, M. Targeted Deletion of PTEN in Kisspeptin Cells Results in Brain Region-and Sex-Specific Effects on Kisspeptin Expression and Gonadotropin Release. Int. J. Mol. Sci. 2020, 21, 2107. [Google Scholar] [CrossRef] [Green Version]
- Misu, Y.; Goshima, Y.; Ueda, H.; Okamura, H. Neurobiology of L-DOPAergic Systems. Prog. Neurobiol. 1996, 49, 415–454. [Google Scholar] [CrossRef]
- Waters, E.M.; Simerly, R.B. Estrogen Induces Caspase-Dependent Cell Death during Hypothalamic Development. J. Neurosci. 2009, 29, 9714–9718. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Ikegami, K.; Nakamura, S.; Uenoyama, Y.; Ozawa, H.; Maeda, K.I.; Tsukamura, H.; Inoue, N. Mating-Induced Increase in Kiss1 MRNA Expression in the Anteroventral Periventricular Nucleus Prior to an Increase in LH and Testosterone Release in Male Rats. J. Reprod. Dev. 2020, 66, 579–586. [Google Scholar] [CrossRef]
- Mohr, M.A.; Wong, A.M.; Sukumar, G.; Dalgard, C.L.; Hong, W.; Wu, T.J.; Wu, Y.E.; Micevych, P.E. RNA-Sequencing of AVPV and ARH Reveals Vastly Different Temporal and Transcriptomic Responses to Estradiol in the Female Rat Hypothalamus. PLoS One 2021, 16. [Google Scholar] [CrossRef]
- Semaan, S.J.; Tolson, K.P.; Kauffman, A.S. The Development of Kisspeptin Circuits in the Mammalian Brain. Adv. Exp. Med. Biol. 2013, 784, 221–252. [Google Scholar] [CrossRef]
- Simerly, R.B. Organization of the Hypothalamus. In The Rat Nervous System, 4th ed.; 2015; pp. 267–294. ISBN 9780080921372. [Google Scholar]
- Rice, D.; Barone, S. Critical Periods of Vulnerability for the Developing Nervous System: Evidence from Humans and Animal Models. Environ. Health Perspect. 2000, 108, 511–533. [Google Scholar] [CrossRef]
- Engelhardt, B. Development of the Blood-Brain Barrier. Cell Tissue Res. 2003, 314, 119–129. [Google Scholar] [CrossRef]
- Ugrumov, M.V. Developing Brain as an Endocrine Organ: A Paradoxical Reality. Neurochem. Res. 2010, 35, 837–850. [Google Scholar]
- Gomes, P.; Soares-da-Silva, P. Na+-Independent Transporters, LAT-2 and B0,+, Exchange L-DOPA with Neutral and Basic Amino Acids in Two Clonal Renal Cell Lines. J. Membr. Biol. 2002, 186, 63–80. [Google Scholar] [CrossRef]
- Kageyama, T.; Nakamura, M.; Matsuo, A.; Yamasaki, Y.; Takakura, Y.; Hashida, M.; Kanai, Y.; Naito, M.; Tsuruo, T.; Minato, N.; et al. The 4F2hc/LAT1 Complex Transports L-DOPA across the Blood-Brain Barrier. Brain Res. 2000, 879. [Google Scholar] [CrossRef]
- Ugrumov, M.V.; Tixier-Vidal, A.; Taxi, J.; Thibault, J.; Mitskevich, M.S. Ontogenesis of Tyrosine Hydroxylase-Immunopositive Structures in the Rat Hypothalamus. Fiber Pathways and Terminal Fields. Neuroscience 1989, 29, 157–166. [Google Scholar] [CrossRef]
- Ugrumov, M.V. Development of the Median Eminence during Ontogenesis (Morpho-Functional Aspects). Prog. Brain Res. 1992, 91, 349–356. [Google Scholar] [CrossRef]
- Ishida, Y.; Yokoyama, C.; Inatomi, T.; Yagita, K.; Dong, X.; Yan, L.; Yamaguchi, S.; Nagatsu, I.; Komori, T.; Kitahama, K.; et al. Circadian Rhythm of Aromatic L-Amino Acid Decarboxylase in the Rat Suprachiasmatic Nucleus: Gene Expression and Decarboxylating Activity in Clock Oscillating Cells. Genes to Cells 2002, 7, 447–459. [Google Scholar] [CrossRef]
- Ugrumov, M.V. Magnocellular Vasopressin System in Ontogenesis: Development and Regulation. Microsc. Res. Tech. 2002, 56, 164–171. [Google Scholar] [CrossRef]
- Liu, L.; Duff, K. A Technique for Serial Collection of Cerebrospinal Fluid from the Cisterna Magna in Mouse. J. Vis. Exp. 2008. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Compact 6th ed. 2009; Volume 170, ISBN 9780125476126. [Google Scholar]
- Yanagida, O.; Kanai, Y.; Chairoungdua, A.; Kim, D.K.; Segawa, H.; Nii, T.; Cha, S.H.; Matsuo, H.; Fukushima, J.I.; Fukasawa, Y.; et al. Human L-Type Amino Acid Transporter 1 (LAT1): Characterization of Function and Expression in Tumor Cell Lines. Biochim. Biophys. Acta - Biomembr. 2001, 1514, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of Protein Using Bicinchoninic Acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Abercrombie, M. Estimation of Nuclear Population from Microtome Sections. Anat. Rec. 1946, 94, 239–247. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ugrumov, M.V.; Pavlova, E.N.; Kolacheva, A.A.; Dil’mukhametova, L.K.; Bogdanov, V.V.; Blokhin, V.; Pronina, T.S. The Periventricular Nucleus as a Brain Center Containing Dopaminergic Neurons and Neurons Expressing Individual Enzymes of Dopamine Synthesis. Int. J. Mol. Sci. 2022, 23, 6739. https://doi.org/10.3390/ijms23126739
Ugrumov MV, Pavlova EN, Kolacheva AA, Dil’mukhametova LK, Bogdanov VV, Blokhin V, Pronina TS. The Periventricular Nucleus as a Brain Center Containing Dopaminergic Neurons and Neurons Expressing Individual Enzymes of Dopamine Synthesis. International Journal of Molecular Sciences. 2022; 23(12):6739. https://doi.org/10.3390/ijms23126739
Chicago/Turabian StyleUgrumov, Michael V., Ekaterina N. Pavlova, Anna A. Kolacheva, Liliya K. Dil’mukhametova, Vsevolod V. Bogdanov, Victor Blokhin, and Tatiana S. Pronina. 2022. "The Periventricular Nucleus as a Brain Center Containing Dopaminergic Neurons and Neurons Expressing Individual Enzymes of Dopamine Synthesis" International Journal of Molecular Sciences 23, no. 12: 6739. https://doi.org/10.3390/ijms23126739