
A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
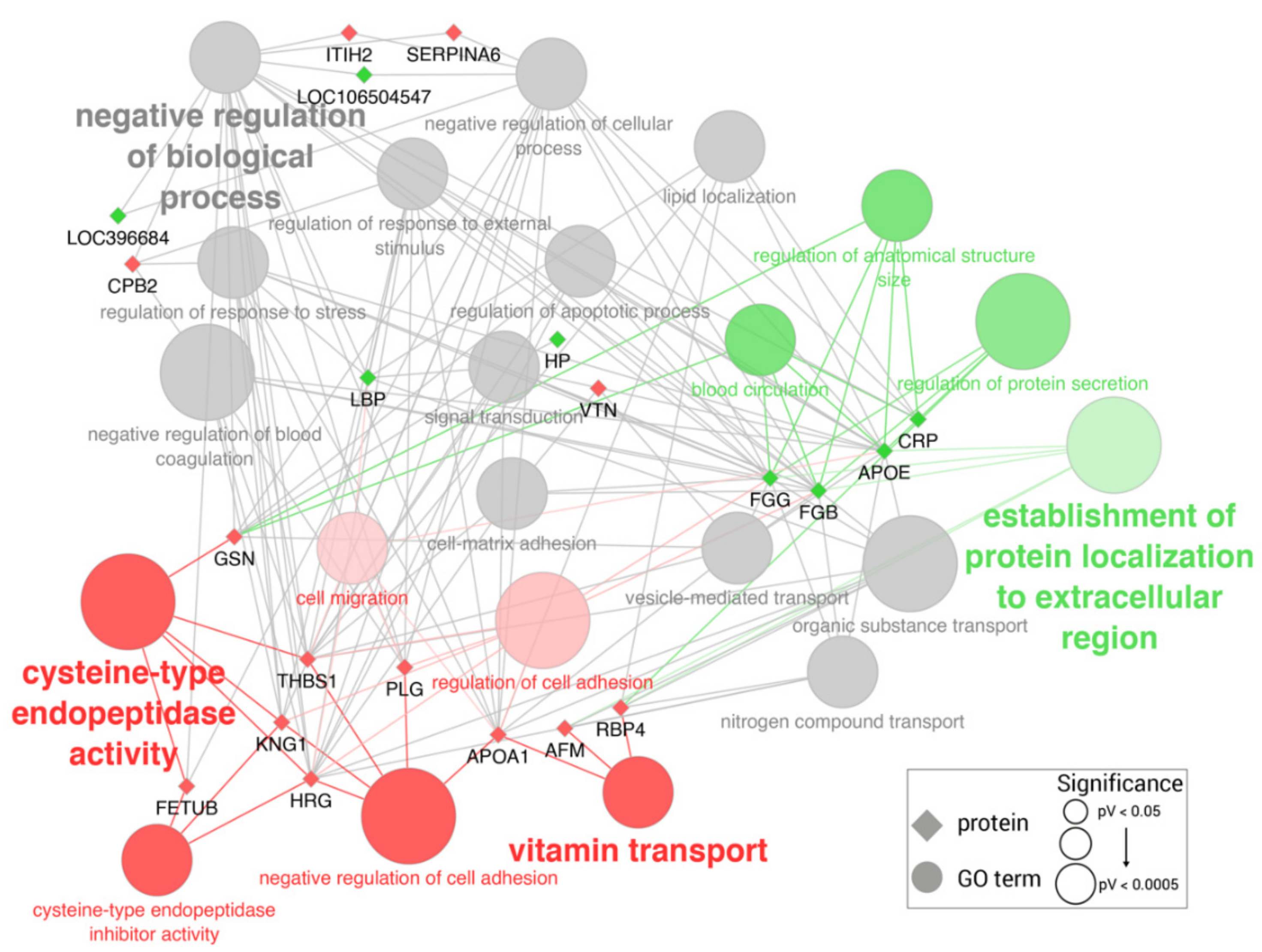
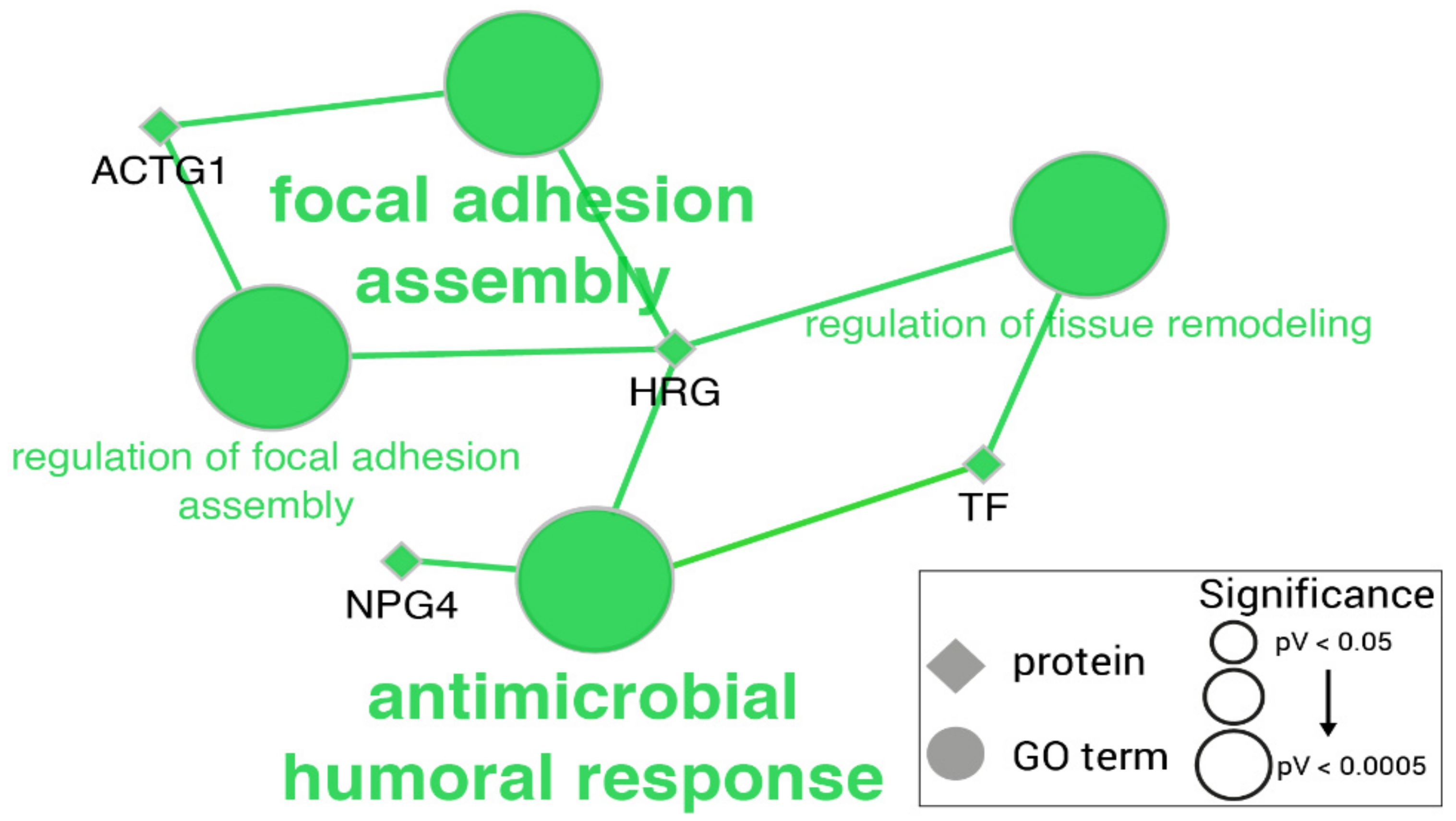
2.1. Proteomic Changes in LPS-Challenged Pigs
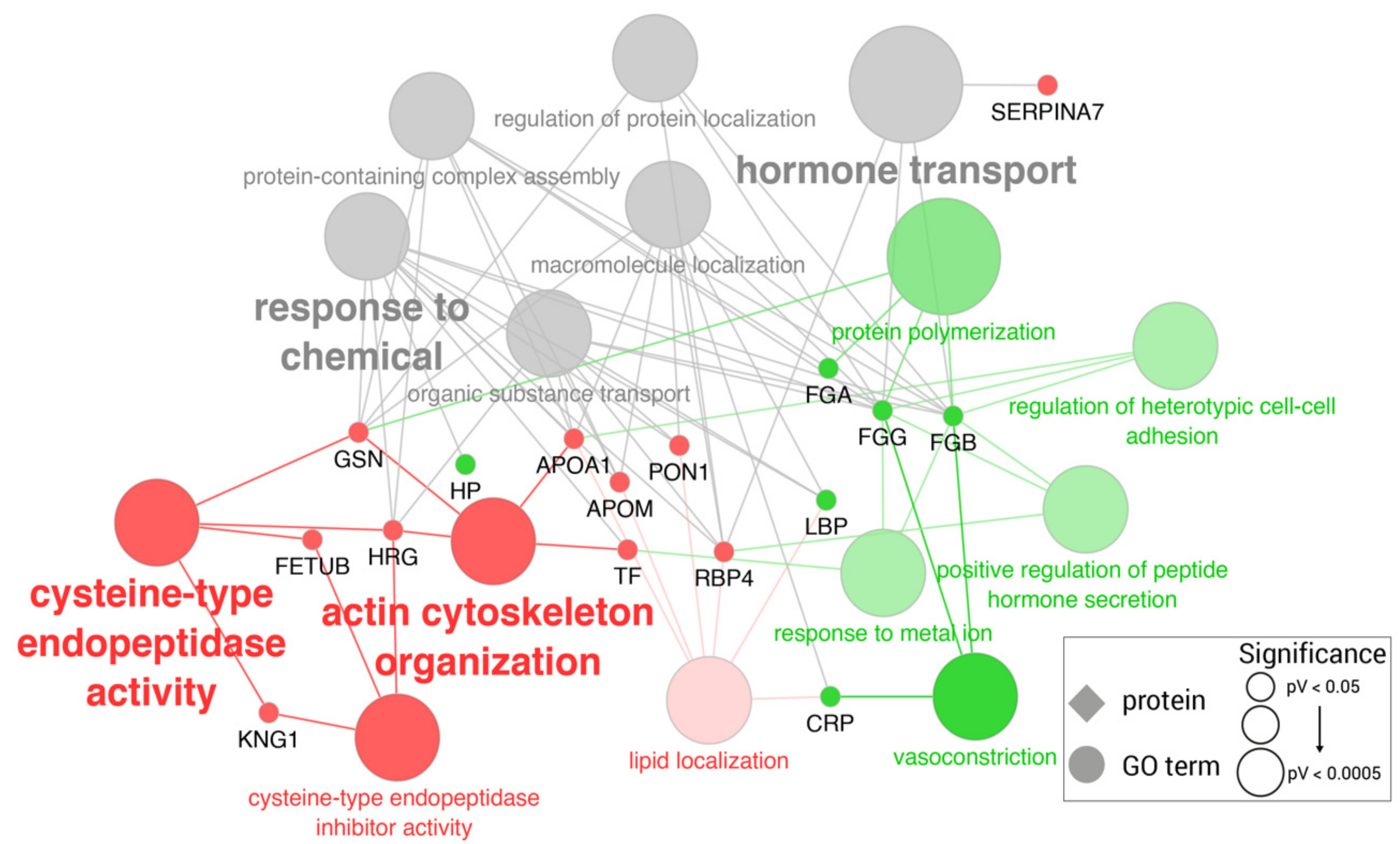
2.2. Proteomic Changes in Turpentine-Challenged Pigs
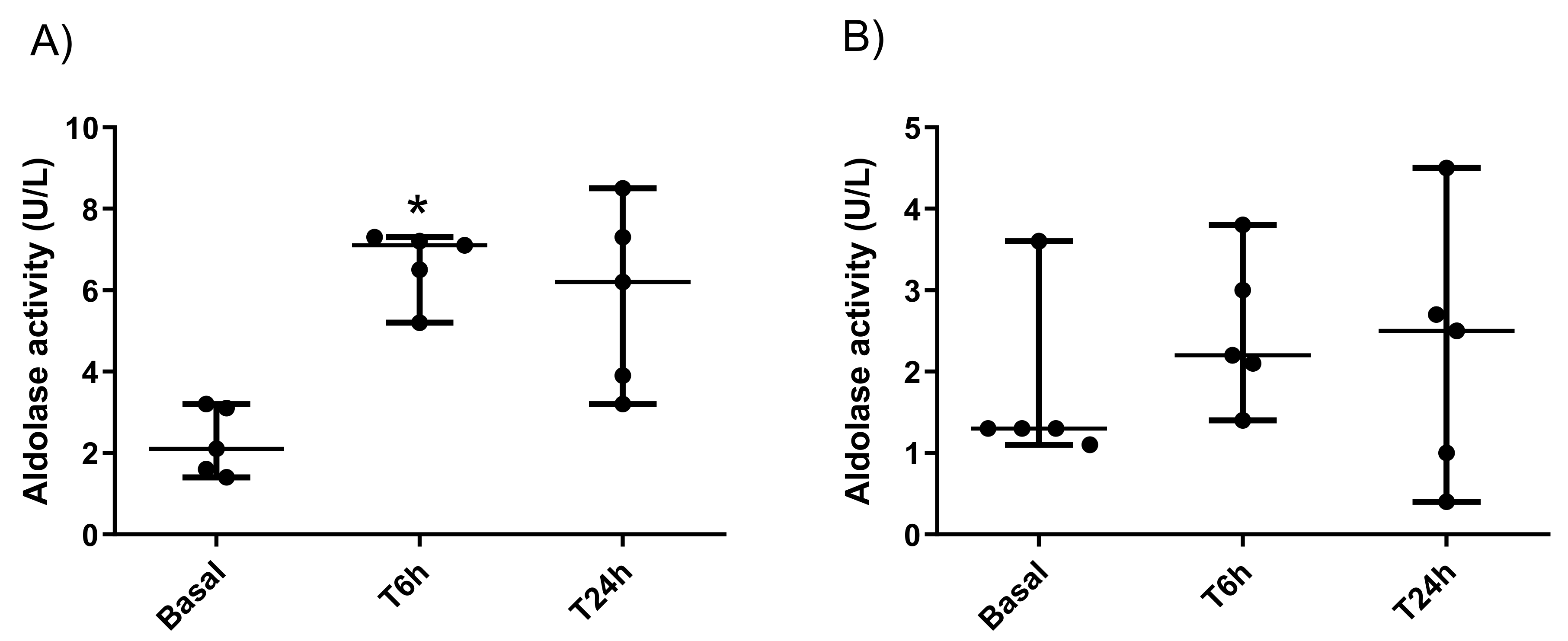
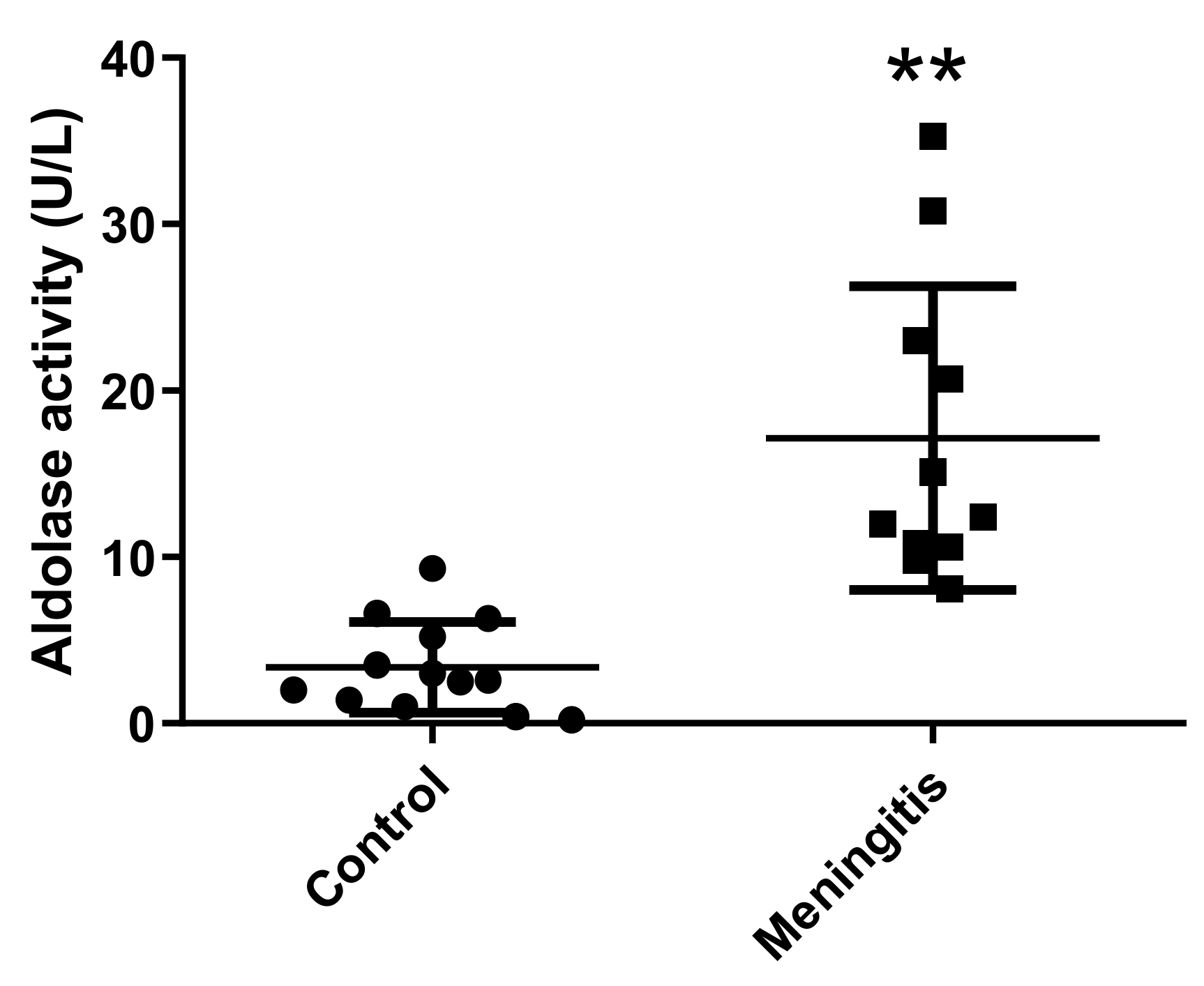
2.3. Measurement of Aldolase Activity in Porcine Saliva
3. Discussion
4. Materials and Methods
4.1. Animals
4.1.1. Proteomic Study
- Lipopolisacharide (LPS) group (n = 5). Pigs were individually administered LPS from Escherichia coli (Sigma-Aldrich, St. Louis, MO, USA) reconstituted in sterile saline solution in a single dose of 30 ug/kg by intramuscular route as previously reported [45].
- Turpentine group (n = 5). Each pig was administered a total of 8 mL subcutaneous injections of turpentine oil (oil of turpentine purified, Sigma–Aldrich, St. Louis, MO, USA), 4 mL in each front flank, as previously described [46].
4.1.2. Validation Study
- Septic and non-septic experimentally-induced inflammation: An aliquot of each saliva sample of the LPS and turpentine groups used in the proteomic study was analyzed.
- Sepsis in field conditions: Two groups of Large White weaning pigs from 6 to 9 weeks old were selected from a commercial farm located in the same geographical area. One was a group of pigs diagnosed with meningitis (n = 11, six males and five females), and the other were clinically healthy pigs (n = 13, seven males and six females). The animals with meningitis had clinical signs compatible with this disease (ataxia, anorexia, lateral recumbency, and padding) [47] and were positive for the presence of Streptococcus suis in bacteriological cultures performed in blood agar plates following standard procedures [48]. Only saliva was obtained in this trial, aiming to avoid the stress associated with blood extraction.
4.2. Sample Collection
4.3. Sample Preparation for Proteomic Analysis
4.4. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) Analysis
4.5. Bioinformatics
4.6. Validation Study
- Precision: The intra- and inter-assay coefficient of variation (CV) were calculated after analyzing two saliva samples of high and low concentration, respectively.
- Accuracy: The indirect evaluation by the linearity under the dilution of a saliva sample with a high ALDOA level.
- LLOQ: The lowest analyte concentration that could be measured with an intra-assay CV < 20%.
- LD: The lowest analyte concentration that could be distinguished from zero value. It was calculated based on data from ten replicate measurements of the zero standard (saline solution) as a mean value plus three standard deviations (SD).
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smyth, M.A.; McDonnell, S.; Perkins, G. Identification of adults with sepsis in the prehospital environment: A systematic review. BMJ Open 2016, 6, e011218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Improving the Prevention, Diagnosis and Clinical Management of Sepsis; Report by the Secretariat; WHO Executive Publication: Geneva, Switzerland, 2017; pp. 1–6. [Google Scholar]
- Alberti, C.; Brun-Buisson, C.; Goodman, S.V.; Guidici, D.; Granton, J.; Moreno, R.; Smithies, M.; Thomas, O.; Artigas, A.; Le Gall, J.R. Influence of Systemic Inflammatory Response Syndrome and Sepsis on Outcome of Critically Ill Infected Patients. Am. J. Respir. Crit. Care Med. 2003, 168, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S. A review of equine sepsis. Equine Vet. Educ. 2015, 27, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Balk, R.A. Systemic inflammatory response syndrome (SIRS). Virulence 2014, 5, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fevre, E.; Gilbert, M.; Grace, D.; Hay, S.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic resistance is the quintessential One Health issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef]
- Grondman, I.; Pirvu, A.; Riza, A.; Ioana, M.; Netea, M.G. Biomarkers of inflammation and the etiology of sepsis. Biochem. Soc. Trans. 2020, 48, 1–14. [Google Scholar] [CrossRef]
- Pierrakos, C.; Vincent, J.-L. Sepsis biomarkers: A review. Crit. Care 2010, 14, R15. [Google Scholar] [CrossRef] [Green Version]
- Cantey, J.B.; Lee, J.H. Biomarkers for the Diagnosis of Neonatal Sepsis. Clin. Perinatol. 2021, 48, 215–227. [Google Scholar] [CrossRef]
- Branche, A.; Neeser, O.; Mueller, B.; Schuetz, P. Procalcitonin to guide antibiotic decision making. Curr. Opin. Infect. Dis. 2019, 32, 130–135. [Google Scholar] [CrossRef]
- EMA (European Medicines Agency); EFSA (European Food Safety Authority). EMA and EFSA Joint Scientific Opinion on measures to reduce the need to use antimicrobial agents in animal husbandry in the European Union, and the resulting impacts on food safety (RONAFA). EFSA J. 2017, 15, e04666. [Google Scholar] [CrossRef]
- Dickson, K.; Lehmann, C. Inflammatory response to different toxins in experimental sepsis models. Int. J. Mol. Sci. 2019, 20, 4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakhshandeh, A.; de Lange, C. Evaluation of chronic immune system stimulation models in growing pigs. Animal 2012, 6, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovici, R.; Neville, L.; Feuerstein, G. Current understanding of sepsis: Criticism and a proposal. J. Endotoxin Res. 1995, 2, 163–168. [Google Scholar] [CrossRef]
- Nemzek, J.A.; Hugunin, K.M.S.; Opp, M.R. Modeling sepsis in the laboratory: Merging sound science with animal well-being. Comp. Med. 2008, 58, 120–128. [Google Scholar] [PubMed]
- Liu, H.; Feye, K.M.; Nguyen, Y.T.; Rakhshandeh, A.; Loving, C.L.; Dekkers, J.C.M.; Gabler, N.K.; Tuggle, C.K. Acute systemic inflammatory response to lipopolysaccharide stimulation in pigs divergently selected for residual feed intake. BMC Genom. 2019, 20, 728. [Google Scholar] [CrossRef] [PubMed]
- Nordgreen, J.; Munsterhjelm, C.; Aae, F.; Popova, A.; Boysen, P.; Ranheim, B.; Heinonen, M.; Raszplewicz, J.; Piepponen, P.; Lervik, A.; et al. The effect of lipopolysaccharide (LPS) on inflammatory markers in blood and brain and on behavior in individually-housed pigs. Physiol. Behav. 2018, 195, 98–111. [Google Scholar] [CrossRef]
- Escribano, D.; Tvarijonaviciute, A.; Tecles, F.; Cerón, J.J. Serum paraoxonase type-1 activity in pigs: Assay validation and evolution after an induced experimental inflammation. Vet. Immunol. Immunopathol. 2015, 163, 210–215. [Google Scholar] [CrossRef]
- Cerón, J.J.; Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Miró, S.; López-Martínez, M.J.; Ortín-Bustillo, A.; Franco-Martínez, L.; Rubio, C.P.; Muñoz-Prieto, A.; Tvarijonaviciute, A.; et al. Basics for the potential use of saliva to evaluate stress, inflammation, immune system, and redox homeostasis in pigs. BMC Vet. Res. 2022, 18, 81. [Google Scholar] [CrossRef]
- Escribano, D.; Campos, P.H.; Gutiérrez, A.M.; Le Floc’H, N.; Cerón, J.J.; Merlot, E. Effect of repeated administration of lipopolysaccharide on inflammatory and stress markers in saliva of growing pigs. Vet. J. 2014, 200, 393–397. [Google Scholar] [CrossRef]
- Tecles, F.; Escribano, D.; Martínez-Miró, S.; Cerón, J. Homocysteine measurement in pig saliva, assay validation and changes after acute stress and experimental inflammation models: A pilot study. Res. Vet. Sci. 2017, 112, 75–80. [Google Scholar] [CrossRef]
- Bilić, P.; Kuleš, J.; Galan, A.; De Pontes, L.G.; Guillemin, N.; Horvatić, A.; Sabes, A.F.; Mrljak, V.; Eckersall, P.D. Proteomics in Veterinary Medicine and Animal Science: Neglected Scientific Opportunities with Immediate Impact. Proteomics 2018, 18, e1800047. [Google Scholar] [CrossRef] [PubMed]
- Olumee-Shabon, Z.; Chattopadhaya, C.; Myers, M.J. Proteomics profiling of swine serum following lipopolysaccharide stimulation. Rapid Commun. Mass Spectrom. 2019, 34, e8639. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.; Dumas-Gaudot, E.; Renaut, J.; Sergeant, K. Gel-based and gel-free quantitative proteomics approaches at a glance. Int. J. Plant Genom. 2012, 2012, 494572. [Google Scholar] [CrossRef] [Green Version]
- Baeumlisberger, D.; Arrey, T.N.; Rietschel, B.; Rohmer, M.; Papasotiriou, D.G.; Mueller, B.; Beckhaus, T.; Karas, M. Labeling elastase digests with TMT: Informational gain by identification of poorly detectable peptides with MALDI-TOF/TOF mass spectrometry. Proteomics 2010, 10, 3905–3909. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Shi, Q.; Chen, Z.; Zhuang, R.; Chen, H.; Zhu, D.; Lou, Y. Skeletal muscle aldolase an overexpression in endotoxemic rats and inhibited by GSNO via potential role for S-nitrosylation in vitro. J. Surg. Res. 2011, 170, e57–e63. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Yeretssian, G.; Doiron, K.; Hussain, S.N.; Saleh, M. The Caspase-1 Digestome Identifies the Glycolysis Pathway as a Target during Infection and Septic Shock. J. Biol. Chem. 2007, 282, 36321–36329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunio, S.; Oldfield, N.; Berry, A.; Ala’Aldeen, D.A.A.; Wooldridge, K.; Turner, D.P.J. The moonlighting protein fructose-1, 6-bisphosphate aldolase of Neisseria meningitidis: Surface localization and role in host cell adhesion. Mol. Microbiol. 2010, 76, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Zhang, W.; Lu, C. Immunoproteomic assay of surface proteins of Streptococcus suis serotype 9. FEMS Immunol. Med. Microbiol. 2008, 53, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirovich, D.B.; Da’Dara, A.A.; Skelly, P.J. Multifunctional fructose 1, 6-bisphosphate aldolase as a therapeutic target. Front. Mol. Biosci. 2021, 8, 719678. [Google Scholar] [CrossRef]
- Goldman, S.E.; Dotan, S.; Talias, A.; Lilo, A.; Azriel, S.; Malka, I.; Portnoi, M.; Ohayon, A.; Kafka, D.; Ellis, R.; et al. Streptococcus pneumoniae fructose-1, 6-bisphosphate aldolase, a protein vaccine candidate, elicits Th1/Th2/Th17-type cytokine responses in mice. Int. J. Mol. Med. 2016, 37, 1127–1138. [Google Scholar] [CrossRef] [Green Version]
- Askew, Y.S.; Pak, S.C.; Luke, C.J.; Askew, D.J.; Cataltepe, S.; Mills, D.R.; Kato, H.; Lehoczky, J.; Dewar, K.; Birren, B.; et al. SERPINB12 is a novel member of the human ov-serpin family that is widely expressed and inhibits trypsin-like serine proteinases. J. Biol. Chem. 2001, 276, 49320–49330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehaus, J.Z.; Good, M.; Jackson, L.; Ozolek, J.A.; Silverman, G.A.; Luke, C.J. Human SERPINB12 is an abundant intracellular serpin expressed in most surface and glandular epithelia. J. Histochem. Cytochem. 2015, 63, 854–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmerhorst, E.J.; Dawes, C.; Oppenheim, F.G. The complexity of oral physiology and its impact on salivary diagnostics. Oral Dis. 2017, 24, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Ekaney, M.L.; Otto, G.P.; Sossdorf, M.; Sponholz, C.; Boehringer, M.; Loesche, W.; Rittirsch, D.; Wilharm, A.; Kurzai, O.; Bauer, M.; et al. Impact of plasma histones in human sepsis and their contribution to cellular injury and inflammation. Crit. Care 2014, 18, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.K.; Ferreira, B.L.; Tashima, A.K.; Brunialti, M.K.C.; Torquato, R.J.S.; Bafi, A.; Assuncao, M.; Azevedo, L.C.P.; Salomao, R. Lipid metabolism impairment in patients with sepsis secondary to hospital acquired pneumonia, a proteomic analysis. Clin. Proteom. 2019, 16, 29. [Google Scholar] [CrossRef]
- Yin, C.; Liu, W.; Liu, Z.; Huang, Y.; Ci, L.; Zhao, R.; Yang, X. Identification of potential serum biomarkers in pigs at early stage after Lipopolysaccharide injection. Res. Vet. Sci. 2017, 111, 140–146. [Google Scholar] [CrossRef]
- Heegaard, P.M.; Stockmarr, A.; Piñeiro, M.; Carpintero, R.; Lampreave, F.; Campbell, F.M.; Eckersall, P.D.; Toussaint, M.J.; Gruys, E.; Sorensen, N.S. Optimal combinations of acute phase proteins for detecting infectious disease in pigs. Vet. Res. 2011, 42, 50. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Zhao, T.; Zhang, W.; He, J.; Lu, F.; Cai, Y.; Lai, Z.; Wei, N.; Liang, C.; Liu, L.; et al. Presence of the apolipoprotein E-ε4 allele is associated with an increased risk of sepsis progression. Sci. Rep. 2020, 10, 15735. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Horvatić, A.; Gelemanović, A.; Samardžija, M.; Mrljak, V.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Dąbrowski, R.; Tvarijonaviciute, A. Changes in the salivary proteome associated with canine pyometra. Front. Vet. Sci. 2020, 7, 277. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Gelemanović, A.; Horvatić, A.; Contreras-Aguilar, M.D.; Dąbrowski, R.; Mrljak, V.; Cerón, J.J.; Martínez-Subiela, S.; Tvarijonaviciute, A. Changes in serum and salivary proteins in canine mammary tumors. Animals 2020, 10, 741. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Muñoz-Prieto, A.; Contreras-Aguilar, M.; Želvytė, R.; Monkevičienė, I.; Horvatić, A.; Kuleš, J.; Mrljak, V.; Cerón, J.; Escribano, D. Changes in saliva proteins in cows with mastitis: A proteomic approach. Res. Vet. Sci. 2021, 140, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Prieto, A.; Contreras-Aguilar, M.D.; Cerón, J.J.; Ayala, I.; Martin-Cuervo, M.; Gonzalez-Sanchez, J.C.; Jacobsen, S.; Kuleš, J.; Beletić, A.; Rubić, I.; et al. Changes in Proteins in Saliva and Serum in Equine Gastric Ulcer Syndrome Using a Proteomic Approach. Animals 2022, 12, 1169. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, M.J.; Escribano, D.; Martínez-Miró, S.; Ramis, G.; Manzanilla, E.G.; Tecles, F.; Martínez-Subiela, S.; Cerón, J.J. Measurement of procalcitonin in saliva of pigs: A pilot study. BMC Vet. Res. 2022, 18, 139. [Google Scholar] [CrossRef] [PubMed]
- Petry, A.; McGilvray, W.; Rakhshandeh, A.R. Technical note: Assessment of an alternative technique for measuring body temperature in pigs. J. Anim. Sci. 2017, 95, 3270. [Google Scholar] [CrossRef]
- Martinez-Subiela, S.; Eckersall, P.D.; Campbell, F.M.; Parra, M.D.; Fuentes, P.; Cerón, J.J. A time-resolved immunofluorometric assay for porcine C-reactive protein quantification in whole blood. Luminescence 2007, 22, 171–176. [Google Scholar] [CrossRef]
- Obradovic, M.R.; Segura, M.; Segalés, J.; Gottschalk, M. Review of the speculative role of co-infections in Streptococcus suis-associated diseases in pigs. Vet. Res. 2021, 52, 49. [Google Scholar] [CrossRef]
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Bowersock, T.L.; Thacker, H.L. Streptococcus suis infection in swine: A retrospective study of 256 cases. Part I. Epidemiologic factors and antibiotic susceptibility patterns. J. Vet. Diagn. Investig. 1993, 5, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Horvatić, A.; Guillemin, N.; Kaab, H.; McKeegan, D.; O’Reilly, E.; Bain, M.; Kuleš, J.; Eckersall, P.D. Quantitative proteomics using tandem mass tags in relation to the acute phase protein response in chicken challenged with Escherichia coli lipopolysaccharide endotoxin. J. Proteom. 2018, 192, 64–77. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Smuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Abundances | Fold Changes | |||||
|---|---|---|---|---|---|---|
| Gene (or Accession Number) | Protein Name | Basal | 6 h | 24 h | 6 h/Basal | 24 h/Basal |
| ALDOA | Fructose-biphosphate aldolase | 0.50 | 1.13 | 0.88 | 1.18 ** | 0.83 |
| SERPINB12 | SERPIN domain-containing protein | 0.42 | 0.92 | 0.74 | 1.12 * | 0.80 |
| ANXA2 | Annexin | 0.50 | 1.02 | 0.65 | 1.04 * | 0.39 |
| SFN | 14-3-3 sigma protein | 0.61 | 1.23 | 0.94 | 1.02 ** | 0.63 |
| MSN | Moesin | 0.67 | 1.34 | 1.12 | 0.99 * | 0.74 |
| SERPINB1 | Leukocyte elastase inhibitor | 0.74 | 1.35 | 0.97 | 0.88 * | 0.39 |
| IGHA | IgM | 0.64 | 1.16 | 0.97 | 0.86 ** | 0.59 |
| ECH1 | Galectin | 0.61 | 1.07 | 0.93 | 0.81 * | 0.60 |
| FABP5 | FABP domain-containing protein | 0.66 | 1.06 | 1.16 | 0.70 | 0.82 * |
| A2M | Alpha-2-macroglobulin isoform a | 0.80 | 1.28 | 1.01 | 0.68 ** | 0.33 |
| IGHG | IgG heavy chain | 0.82 | 1.28 | 0.80 | 0.63 * | -0.04 |
| LMNA | Lamin isoform A | 0.74 | 1.11 | 1.01 | 0.59 * | 0.45 |
| P4HB | Protein disulfide-isomerase | 0.79 | 1.19 | 1.06 | 0.59 * | 0.43 |
| TKT | Transketolase | 0.75 | 1.13 | 1.03 | 0.59 * | 0.45 |
| YWHAZ | 14-3-3 protein zeta/delta | 0.64 | 0.86 | 1.13 | 0.43 | 0.83 * |
| CSTB | Cystatin-B | 0.80 | 1.06 | 1.35 | 0.40 | 0.75 * |
| LCN2 | Neutrophil gelatinase-associated lipocalin | 0.72 | 0.81 | 1.31 | 0.18 | 0.86 * |
| P51524 (accession) | Prophenin and tritrpticin precursor (Fragment) | 0.73 | 0.54 | 1.71 | −0.43 | 1.23 * |
| Mean Abundances | Fold Changes | |||||
|---|---|---|---|---|---|---|
| Gene (or Accession Number) | Protein Name | Basal | 6 h | 24 h | 6 h/Basal | 24 h/Basal |
| LOC106504547 | SERPIN domain-containing protein | 0.78 | 1.02 | 1.66 | 0.43 | 1.13 *** |
| LOC396684 | SERPIN domain-containing protein | 0.90 | 1.07 | 1.51 | 0.25 | 0.74 ** |
| HP | Haptoglobin | 0.77 | 0.98 | 1.21 | 0.34 | 0.63 ** |
| CRP | Pentaxin/C-reactive protein | 0.56 | 1.09 | 0.86 | 0.96 ** | 0.62 |
| APOE | Apolipoprotein E | 0.97 | 1.14 | 1.04 | 0.23 * | 0.10 |
| LBP | Lipopolysaccharide-binding protein | 0.84 | 0.95 | 1.08 | 0.17 | 0.36 * |
| LUM | Lumican | 0.94 | 0.99 | 1.18 | 0.07 | 0.32 * |
| A0A480XY00 | Complement C1s subcomponent isoform 1 preproprotein | 0.98 | 0.98 | 1.10 | −0.01 | 0.16 ** |
| FGB | Fibrinogen beta chain | 0.94 | 0.78 | 1.10 | −0.26 ** | 0.22 |
| FGG | Fibrinogen C-terminal domain-containing | 0.99 | 0.81 | 1.06 | −0.29 * | 0.10 |
| FGA | Fibrinogen alpha chain | 0.96 | 0.78 | 1.02 | −0.29 * | 0.08 |
| FN1 | Fibronectin | 1.03 | 0.91 | 0.95 | −018 * | −0.12 |
| ALB | Albumin | 1.01 | 1.00 | 0.93 | −0.02 | −0.12 * |
| SERPINC1 | Antithrombin-III | 0.99 | 0.94 | 0.90 | −0.06 | −0.13 * |
| SERPINA7 | Thyroxine-binding globulin | 1.02 | 0.99 | 0.92 | −0.10 | −0.14 * |
| ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 isoform a preproprotein | 1.06 | 1.00 | 0.95 | −0.08 | −0.14 * |
| C8B | Complement component 8 subunit beta | 1.01 | 1.04 | 0.90 | 0.04 | −016 * |
| PROC | Vitamin K-dependent protein C | 1.16 | 1.08 | 1.02 | −010 | −0.19 * |
| ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 | 1.07 | 1.02 | 0.92 | −0.06 | −0.22 * |
| A0A4X1TBX0 | C1q domain-containing protein | 1.01 | 0.95 | 0.86 | −0.08 | −0.22 * |
| C8G | Complement component C8G | 1.03 | 1.04 | 0.88 | 0.01 | −0.23 * |
| AFM | Afamin | 1.09 | 1.03 | 0.92 | −0.07 | −0.24 * |
| SERPINA6 | SERPIN domain-contaning protein | 1.03 | 1.03 | 0.85 | −0.01 | −0.27 * |
| PLG | Plasminogen | 1.02 | 1.00 | 0.84 | −0.02 | −0.27 ** |
| GSN | Actin-depolymerizing factor | 1.06 | 1.05 | 0.87 | −0.01 | −0.27 * |
| FETUB | Fetuin-B isoform 1 | 1.11 | 0.99 | 0.01 | −0.17 | −0.28 ** |
| HRG | Histidine-rich glycoprotein | 1.00 | 0.97 | 0.80 | −0.03 | −0.31 ** |
| APOA1 | Apolipoprotein A-1 | 1.13 | 1.02 | 0.90 | −0.15 | −0.32 * |
| CPB2 | Carboxypeptidase B2 isoform 1 preproprotein | 1.06 | 1.06 | 0.84 | -0.01 | −0.32 ** |
| VTN | Vitronectin | 1.09 | 0.92 | 0.82 | −0.24 | −0.40 ** |
| Mean Abundances | Fold Changes | |||||
|---|---|---|---|---|---|---|
| Gene | Protein Name | Basal | 6 h | 24 h | 6 h/Basal | 24 h/Basal |
| P62802 | Histone H4 | 0.49 | 1.13 | 0.73 | 1.18 * | 0.55 |
| ALB | Albumin | 0.54 | 1.12 | 0.76 | 1.05 ** | 0.50 |
| HRG | Cystatin domain-containing protein | 0.67 | 1.25 | 0.90 | 0.88 * | 0.42 |
| A2M | Alpha-2-macroglobulin isoform a | 0.61 | 1.03 | 0.73 | 0.75 * | 0.24 |
| TF | Beta-1 metal-binding globulin | 0.72 | 1.14 | 0.93 | 0.65 ** | 0.37 |
| IGHG | IgG heavy chain | 0.67 | 1.05 | 0.90 | 0.64 * | 0.42 |
| P51524 (accession) | Prophenin and tritrpticin precursor (Fragment) | 0.54 | 0.82 | 1.46 | 0.58 | 1.41 * |
| LOC106504547 | SERPIN domain-containing protein | 0.69 | 0.90 | 1.15 | 0.37 | 0.73 ** |
| LCN2 | Neutrophil gelatinase-associated lipocalin | 0.70 | 0.89 | 1.17 | 0.34 | 0.72 * |
| Mean Abundances | Fold Changes | |||||
|---|---|---|---|---|---|---|
| Gene (or Accession Number) | Protein Name | Basal | 6 h | 24 h | 6 h/Basal | 24 h/Basal |
| CRP | Pentaxin or C-reactive protein | 0.46 | 0.66 | 1.30 | 0.50 | 1.47 * |
| LOC106504547 | SERPIN domain-containing protein | 0.688 | 0.73 | 1.74 | 0.08 | 1.32 ** |
| LOC100156325 | SERPIN domain-containing protein | 0.69 | 0.83 | 1.13 | 0.27 | 0.71 * |
| HP | Haptoglobin | 0.80 | 0.87 | 1.29 | 0.10 | 0.67 ** |
| FGA | Fibrinogen alpha-chain | 0.85 | 0.91 | 1.32 | 0.09 | 0.62 ** |
| LBP | Lipopolysaccharide-binding protein | 0.90 | 0.98 | 1.34 | 0.11 | 0.57 ** |
| FGB | Fibrinogen beta chain | 0.86 | 0.91 | 1.19 | 0.09 | 0.47 ** |
| FGG | Fibrinogen C-terminal domain-containing protein | 0.91 | 0.93 | 1.21 | 0.02 | 0.42 ** |
| A0A4X1U9T5 (accession) | Ig-like domain-containing protein | 0.97 | 0.95 | 1.11 | −0.02 | 0.19 * |
| APOA1 | Apolipoprotein A-1 | 1.25 | 1.13 | 0.72 | −0.13 | −0.8 ** |
| C8A | MACPF domain-containing protein | 1.27 | 1.11 | 0.90 | −0.19 | −0.48 * |
| RBP4 | Plasma retinol-binding protein | 1.10 | 1.07 | 0.79 | −0.03 | −0.47 * |
| SERPINA6 | SERPIN domain-containing protein | 1.12 | 1.01 | 0.82 | −0.14 | −0.45 ** |
| VTN | Vitronectin | 1.19 | 1.16 | 0.92 | −0.03 | −0.36 ** |
| APON | Ovarian and testicular apolipoprotein N | 1.08 | 0.99 | 0.85 | −0.13 | −0.34 ** |
| ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 isoform a preproprotein | 1.01 | 1.03 | 0.83 | 0.02 | −0.29 ** |
| HRG | Histidine-rich glycoprotein | 1.09 | 1.04 | 0.89 | −0.06 | −0.28 * |
| GSN | Actin-depolymerizing factor | 1.03 | 0.99 | 0.88 | −0.05 | −0.23 * |
| FETUB | Fetuin-B isoform 1 | 1.11 | 1.09 | 0.95 | −0.02 | −0.23 * |
| TF | Serotransferrin | 1.06 | 1.07 | 0.91 | 0.01 | −0.22 * |
| A0SEH3 (accession) | Complement component C8G | 1.00 | 0.98 | 0.88 | −0.03 | −0.18 * |
| ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 | 1.05 | 1.03 | 0.92 | −0.02 | −0.18 * |
| AMBP | Alpha-1-microglobulin | 1.01 | 0.98 | 0.89 | −0.04 | −0.18 * |
| PROC | Vitamin K-dependent protein | 1.10 | 1.07 | 0.99 | −0.04 | −0.15 * |
| A1BG | Alpha-1B-glycoprotein | 1.06 | 1.04 | 0.97 | −0.01 | −0.12 * |
| A1BG | Alpha-1B-glycoprotein | 1.06 | 1.04 | 0.97 | −0.01 | −0.12 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Martínez, M.J.; Cerón, J.J.; Ortín-Bustillo, A.; Escribano, D.; Kuleš, J.; Beletić, A.; Rubić, I.; González-Sánchez, J.C.; Mrljak, V.; Martínez-Subiela, S.; et al. A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation. Int. J. Mol. Sci. 2022, 23, 6738. https://doi.org/10.3390/ijms23126738
López-Martínez MJ, Cerón JJ, Ortín-Bustillo A, Escribano D, Kuleš J, Beletić A, Rubić I, González-Sánchez JC, Mrljak V, Martínez-Subiela S, et al. A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation. International Journal of Molecular Sciences. 2022; 23(12):6738. https://doi.org/10.3390/ijms23126738
Chicago/Turabian StyleLópez-Martínez, María José, José Joaquín Cerón, Alba Ortín-Bustillo, Damián Escribano, Josipa Kuleš, Anđelo Beletić, Ivana Rubić, Juan Carlos González-Sánchez, Vladimir Mrljak, Silvia Martínez-Subiela, and et al. 2022. "A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation" International Journal of Molecular Sciences 23, no. 12: 6738. https://doi.org/10.3390/ijms23126738