
Intestinal Epithelial Cells Modulate the Production of Enterotoxins by Porcine Enterotoxigenic E. coli Strains


Abstract
:1. Introduction
2. Results
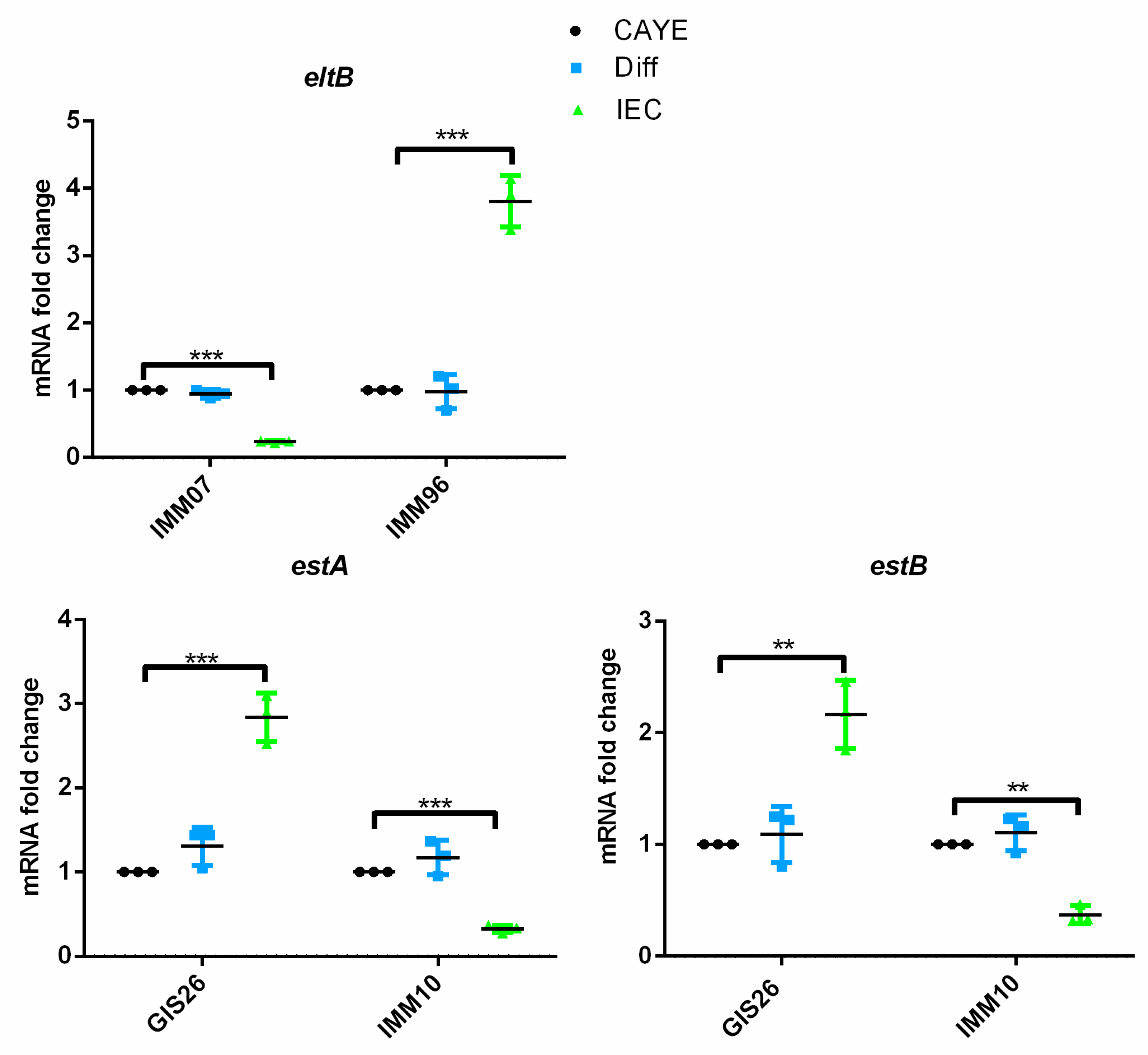
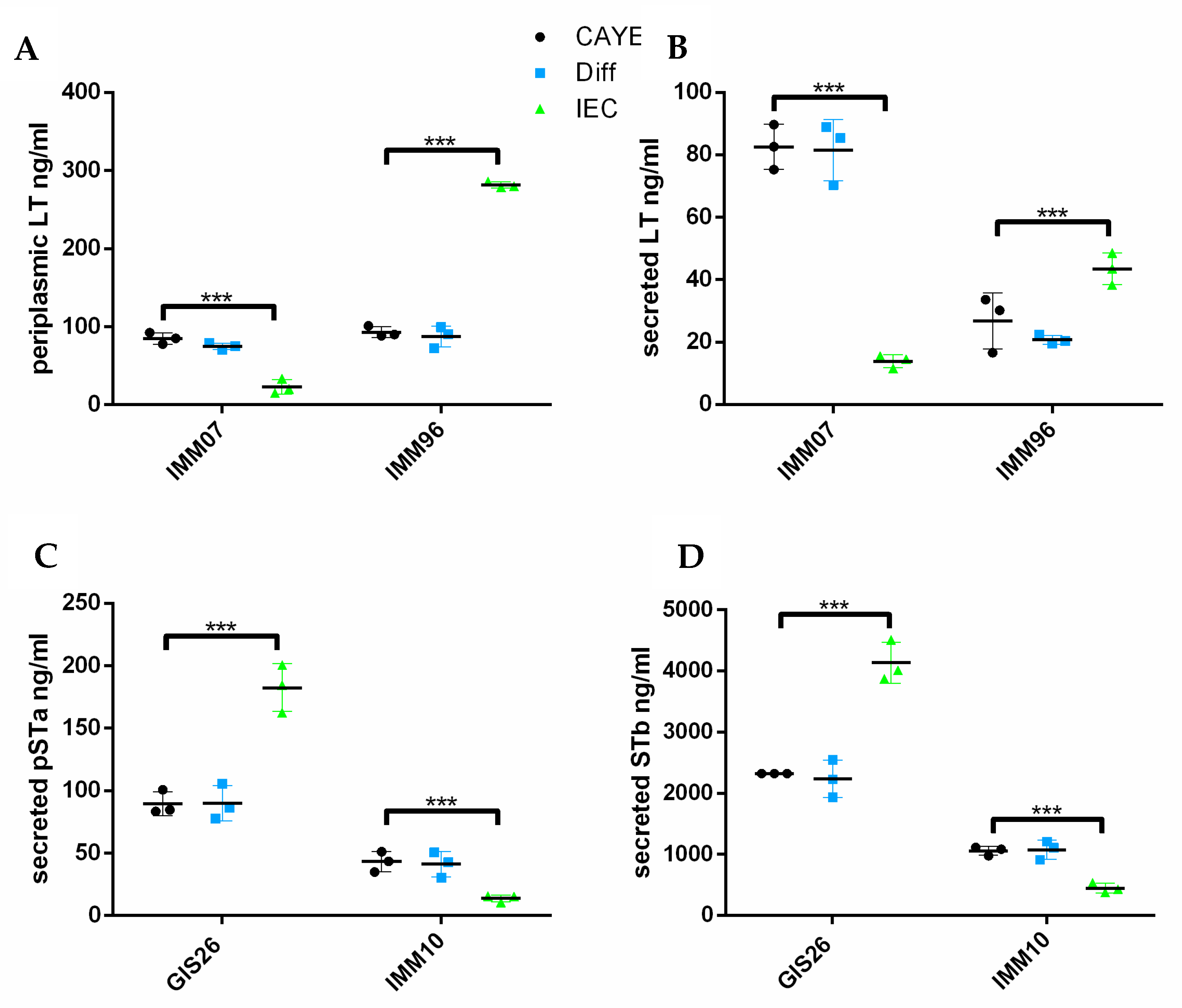
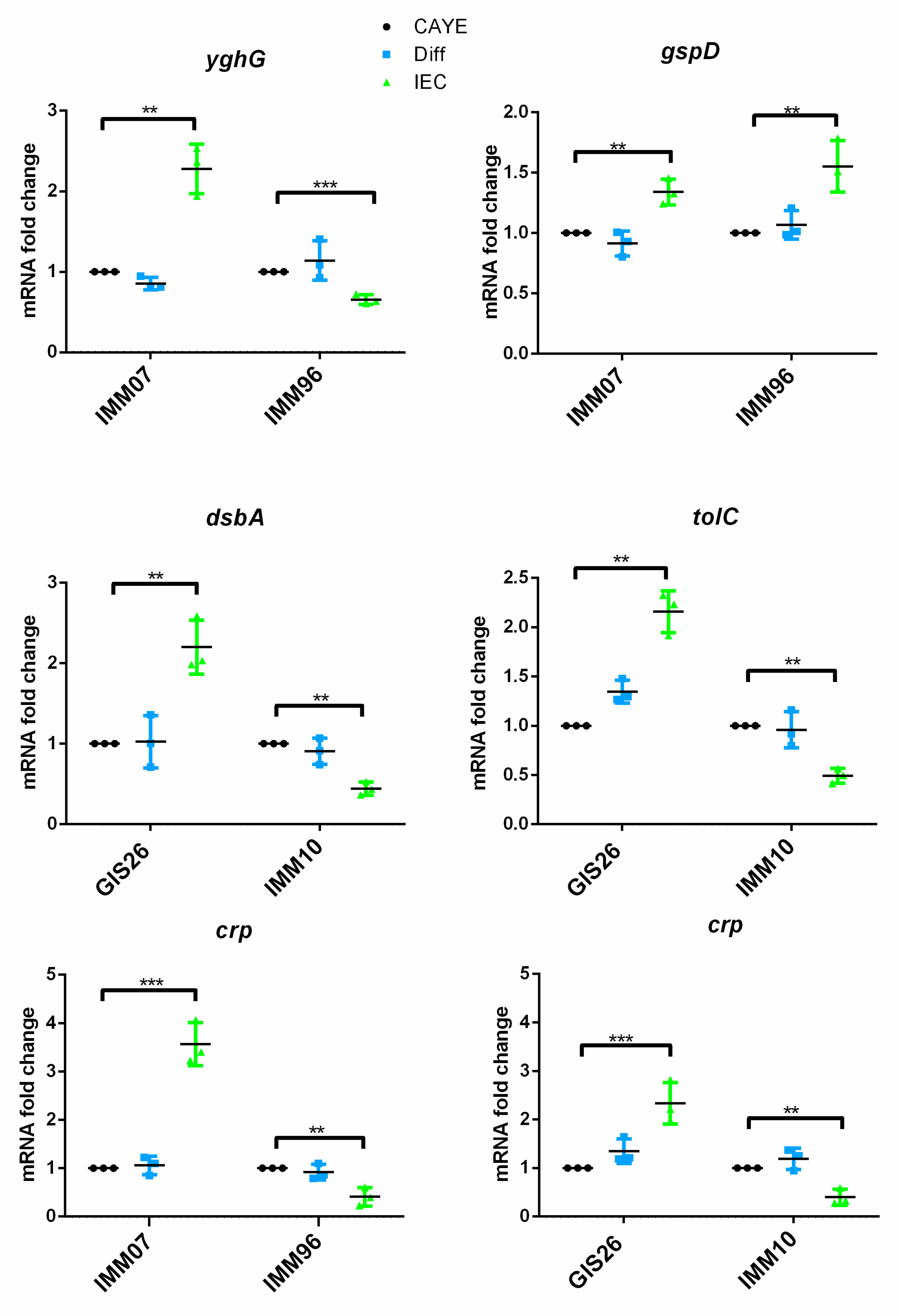
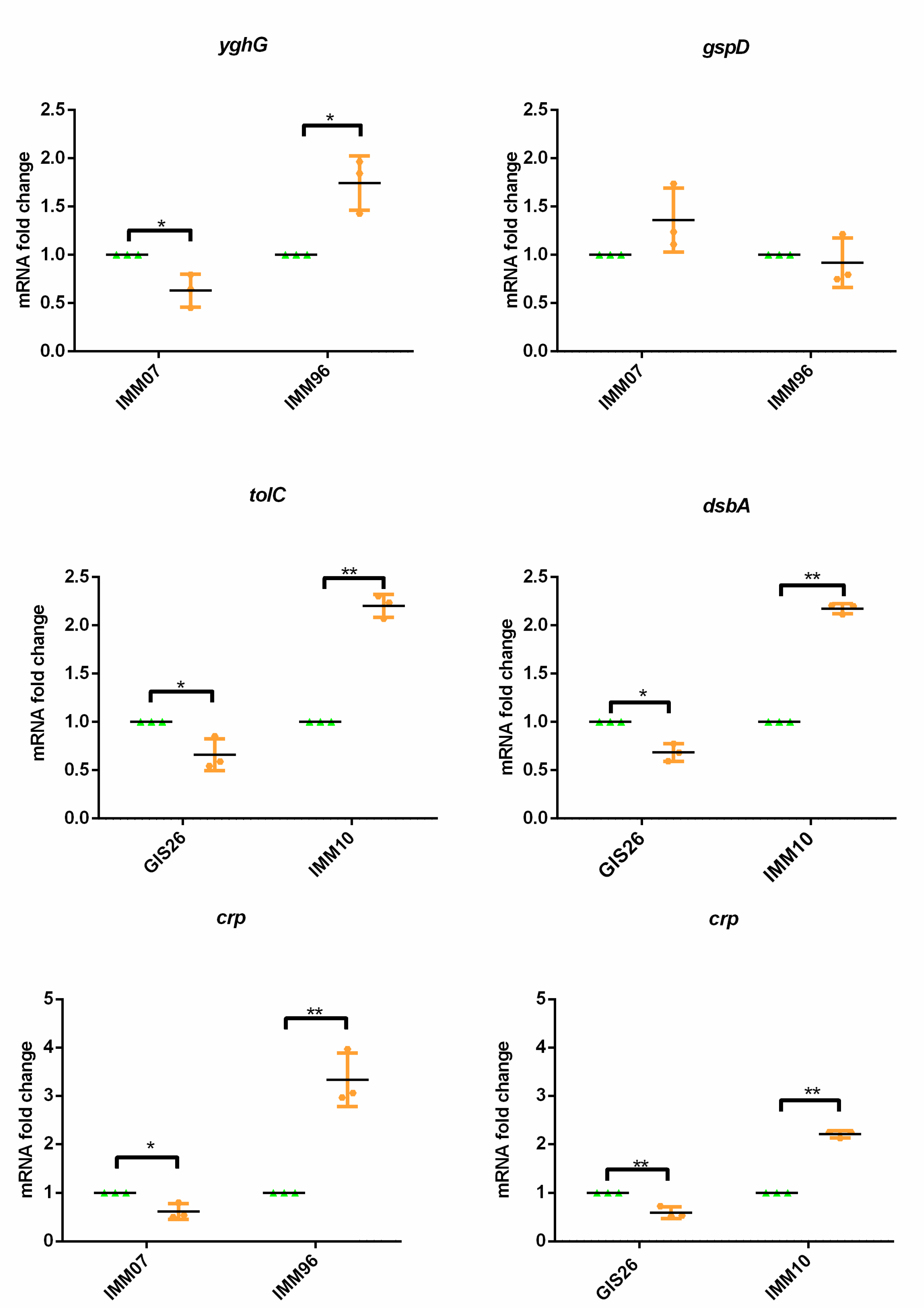
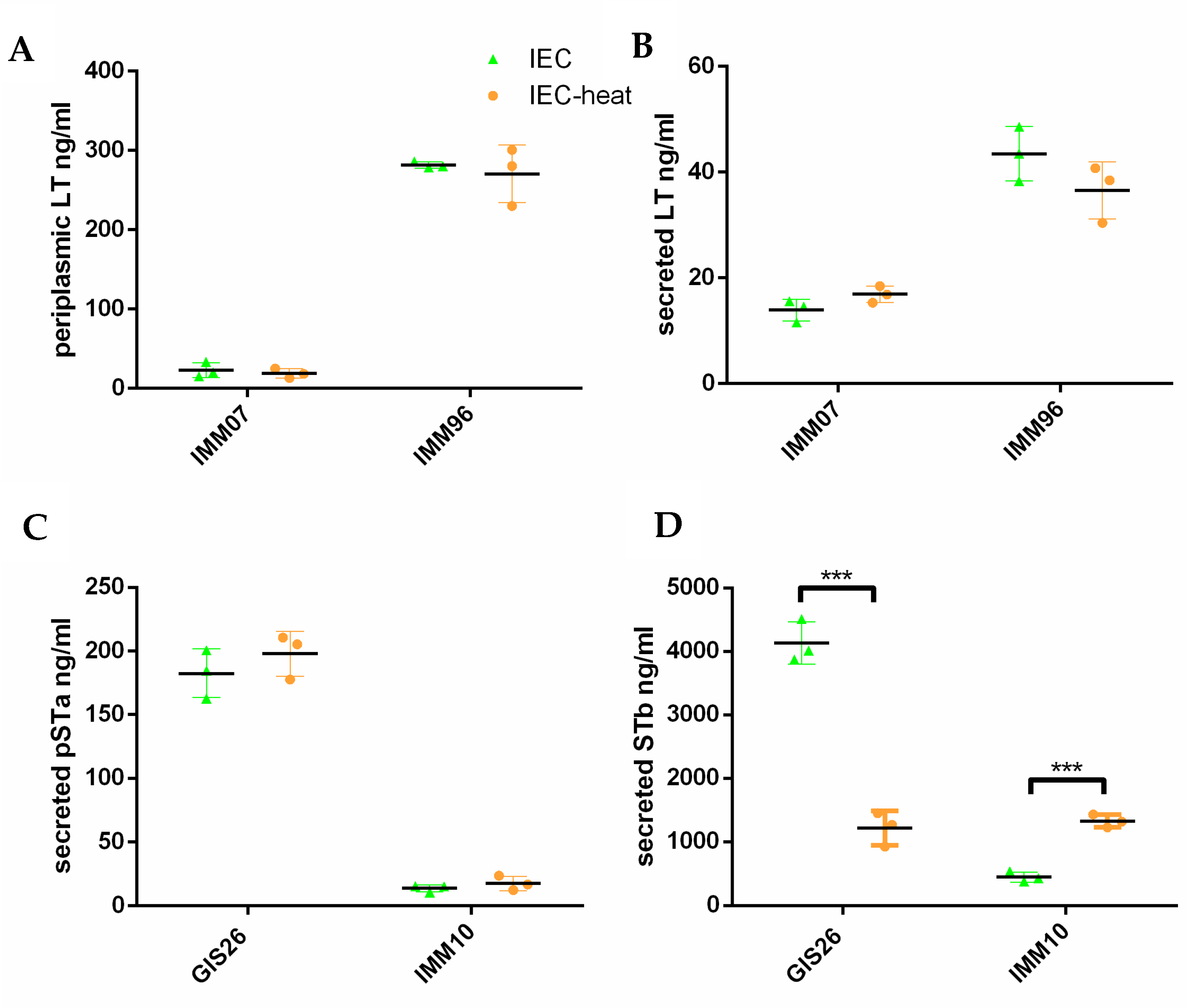
2.1. Epithelial Factors Modulate Expression of Virulence Factors in ETEC Isolates Differing in Their Enterotoxin Secretion
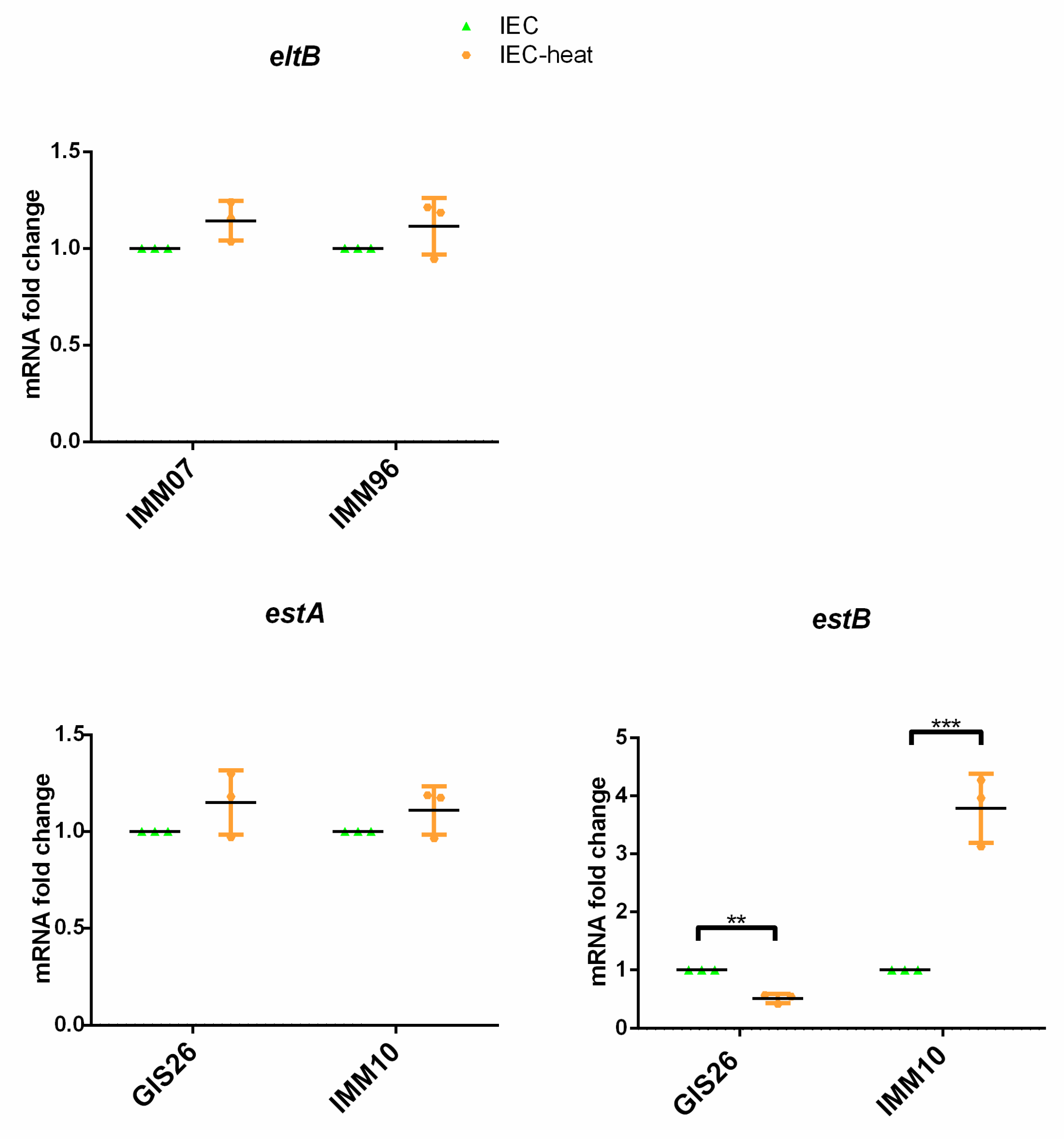
2.2. Heat-Treatment of Epithelial Factors Resulted in Opposite Transcriptional Levels of Virulence Factors in ETEC
3. Discussion
4. Materials and Methods
4.1. Preparation of Intestinal Epithelial Cell-Conditioned Medium
4.2. Bacterial Strains and Growth Conditions
4.3. Isolation of Total Bacterial RNA
4.4. qPCR
4.5. Detection of Enterotoxins Secreted by ETEC Strains Grown in Different Media
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Melkebeek, V.; Goddeeris, B.; Cox, E. ETEC vaccination in pigs. Vet. Immunol. Immunopathol. 2013, 152, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kuhlmann, F.M.; Chakraborty, S.; Bourgeois, A.L.; Foulke-Abel, J.; Tumala, B.; Vickers, T.J.; Sack, D.A.; DeNearing, B.; Harro, C.D.; et al. Enterotoxigenic Escherichia coli–blood group A interactions intensify diarrheal severity. J. Clin. Investig. 2018, 128, 3298–3311. [Google Scholar] [CrossRef] [Green Version]
- von Mentzer, A.; Tobias, J.; Wiklund, G.; Nordqvist, S.; Aslett, M.; Dougan, G.; Sjöling, A.; Svennerholm, A.-M. Identification and characterization of the novel colonization factor CS30 based on whole genome sequencing in enterotoxigenic Escherichia coli (ETEC). Sci. Rep. 2017, 7, 12514. [Google Scholar] [CrossRef] [Green Version]
- Kharat, V.B.; Ahmed, M.; Jiang, Z.-D.; Riddle, M.S.; DuPont, H.L. Colonization Factors in Enterotoxigenic Escherichia coli Strains in Travelers to Mexico, Guatemala, and India Compared with Children in Houston, Texas. Am. J. Trop. Med. Hyg. 2017, 96, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, A.; Luo, Q.; Roy, K.; Shabaan, S.; Kumar, P.; Qadri, F.; Fleckenstein, J.M. Contribution of the Highly Conserved EaeH Surface Protein to Enterotoxigenic Escherichia coli Pathogenesis. Infect. Immun. 2014, 82, 3657–3666. [Google Scholar] [CrossRef] [Green Version]
- Roy, K.; Hilliard, G.M.; Hamilton, D.J.; Luo, J.; Ostmann, M.M.; Fleckenstein, J. Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 2009, 457, 594–598. [Google Scholar] [CrossRef]
- Nagy, B.; Fekete, P.Z. Enterotoxigenic Escherichia coli in veterinary medicine. Int. J. Med. Microbiol. 2005, 295, 443–454. [Google Scholar] [CrossRef]
- Wang, H.; Zhong, Z.; Luo, Y.; Cox, E.; Devriendt, B. Heat-Stable Enterotoxins of Enterotoxigenic Escherichia coli and Their Impact on Host Immunity. Toxins 2019, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Garcia, R.S.; Cox, E.; Devriendt, B. Porcine Enterotoxigenic Escherichia coli Strains Differ in Their Capacity to Secrete Enterotoxins through Varying YghG Levels. Appl. Environ. Microbiol. 2020, 86, e00523-20. [Google Scholar] [CrossRef] [PubMed]
- Wijemanne, P.; Xing, J.; Berberov, E.M.; Marx, D.B.; Francis, D.H.; Moxley, R.A. Relationship between Heat-Labile Enterotoxin Secretion Capacity and Virulence in Wild Type Porcine-Origin Enterotoxigenic Escherichia coli Strains. PLoS ONE 2015, 10, e0117663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devriendt, B.; Stuyven, E.; Verdonck, F.; Goddeeris, B.; Cox, E. Enterotoxigenic Escherichia coli (K88) induce proinflammatory responses in porcine intestinal epithelial cells. Dev. Comp. Immunol. 2010, 34, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Motyka, N.I.; Stewart, S.R.; Hollifield, I.E.; Kyllo, T.R.; Mansfield, J.A.; Norton, E.B.; Clements, J.D.; Bitoun, J.P. Elevated Extracellular cGMP Produced after Exposure to Enterotoxigenic Escherichia coli Heat-Stable Toxin Induces Epithelial IL-33 Release and Alters Intestinal Immunity. Infect. Immun. 2021, 89, e00707-20. [Google Scholar] [CrossRef]
- Johnson, A.M.; Kaushik, R.S.; Francis, D.H.; Fleckenstein, J.M.; Hardwidge, P.R. Heat-Labile Enterotoxin Promotes Escherichia coli Adherence to Intestinal Epithelial Cells. J. Bacteriol. 2009, 191, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gao, X.; Hardwidge, P.R. Heat-labile enterotoxin-induced activation of NF-κB and MAPK pathways in intestinal epithelial cells impacts enterotoxigenic Escherichia coli (ETEC) adherence. Cell. Microbiol. 2012, 14, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, A.; Tumula, B.; Vickers, T.J.; Alvarado, D.; Ciorba, M.A.; Bhuiyan, T.R.; Qadri, F.; Singer, B.B.; Fleckenstein, J.M. CEACAMs serve as toxin-stimulated receptors for enterotoxigenic Escherichia coli. Proc. Natl. Acad. Sci. USA 2020, 117, 29055–29062. [Google Scholar] [CrossRef]
- Sturbelle, R.T.; Avila, L.F.D.C.D.; Roos, T.B.; Borchardt, J.L.; Conceição, R.D.C.D.S.D.; Dellagostin, O.A.; Leite, F.P.L. The role of quorum sensing in Escherichia coli (ETEC) virulence factors. Vet. Microbiol. 2015, 180, 245–252. [Google Scholar] [CrossRef]
- Crofts, A.A.; Giovanetti, S.M.; Rubin, E.J.; Poly, F.M.; Gutiérrez, R.L.; Talaat, K.R.; Porter, C.K.; Riddle, M.S.; DeNearing, B.; Brubaker, J.; et al. Enterotoxigenic E. coli virulence gene regulation in human infections. Proc. Natl. Acad. Sci. USA 2018, 115, E8968–E8976. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.A.; Schifferli, D.M. Differential regulation of fasA and fasH expression of Escherichia coli 987P fimbriae by environmental cues. Mol. Microbiol. 1997, 25, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.M.; Åberg, A.; Straseviçiene, J.; Emődy, L.; Uhlin, B.E.; Balsalobre, C. Type 1 Fimbriae, a Colonization Factor of Uropathogenic Escherichia coli, Are Controlled by the Metabolic Sensor CRP-cAMP. PLoS Pathog. 2009, 5, e1000303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haycocks, J.R.J.; Sharma, P.; Stringer, A.M.; Wade, J.; Grainger, D.C. The Molecular Basis for Control of ETEC Enterotoxin Expression in Response to Environment and Host. PLoS Pathog. 2015, 11, e1004605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijemanne, P.; Moxley, R.A. Glucose Significantly Enhances Enterotoxigenic Escherichia coli Adherence to Intestinal Epithelial Cells through Its Effects on Heat-Labile Enterotoxin Production. PLoS ONE 2014, 9, e113230. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, L.; Ali, Z.B.; Nygren, E.; Wang, Z.; Karlsson, S.; Zhu, B.; Quiding-Järbrink, M.; Sjöling, Å. Alkaline pH Is a Signal for Optimal Production and Secretion of the Heat Labile Toxin, LT in Enterotoxigenic Escherichia coli (ETEC). PLoS ONE 2013, 8, e74069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G. Diarrheagenic Escherichia coli Signaling and Interactions with Host Innate Immunity and Intestinal Microbiota. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 2017. [Google Scholar]
- Busque, P.; Letellier, A.; Harel, J.; Dubreuil, J.D. Production of Escherichia coli STb enterotoxin is subject to catabolite repression. Microbiology 1995, 141, 1621–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorsey, F.C.; Fischer, J.F.; Fleckenstein, J.M. Directed delivery of heat-labile enterotoxin by enterotoxigenic Escherichia coli. Cell. Microbiol. 2006, 8, 1516–1527. [Google Scholar] [CrossRef]
- Kishigami, S.; Ito, K. Roles of cysteine residues of DsbB in its activity to reoxidize DsbA, the protein disulphide bond catalyst of Escherichia coli. Genes Cells 1996, 1, 201–208. [Google Scholar] [CrossRef]
- Yamanaka, H.; Kameyama, M.; Baba, T.; Fujii, Y.; Okamoto, K. Maturation pathway of Escherichia coli heat-stable enterotoxin I: Requirement of DsbA for disulfide bond formation. J. Bacteriol. 1994, 176, 2906–2913. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, E.; Fairbrother, J.M.; Zentek, J.; Belanger, L.; Tremblay, D.; Tremblay, C.L.; Rohe, I.; Vahjen, W.; Brunelle, M.; Hellmann, K.; et al. Efficacy of a single oral dose of a live bivalent E. coli vaccine against post-weaning diarrhea due to F4 and F18-positive enterotoxigenic E. coli. Vet. J. 2017, 226, 32–39. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, J.; Zhang, C.; Jiang, C.; Ma, Y.; He, H.; Wu, Y.; Devriendt, B.; Cox, E.; Zhang, H. Toll-like receptor 5-mediated IL-17C expression in intestinal epithelial cells enhances epithelial host defense against F4+ ETEC infection. Vet. Res. 2019, 50, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Van Nguyen, U.; Rodriguez, P.Y.D.L.F.; Devriendt, B.; Cox, E. F4+ ETEC infection and oral immunization with F4 fimbriae elicits an IL-17-dominated immune response. Vet. Res. 2015, 46, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, M.; Geens, M.; Schauvliege, S.; Gasthuys, F.; van der Meulen, J.; Dubreuil, J.D.; Goddeeris, B.M.; Niewold, T.; Cox, E. Role of Heat-Stable Enterotoxins in the Induction of Early Immune Responses in Piglets after Infection with Enterotoxigenic Escherichia coli. PLoS ONE 2012, 7, e41041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Luo, Q.; Vickers, T.J.; Sheikh, A.; Lewis, W.G.; Fleckenstein, J. EatA, an Immunogenic Protective Antigen of Enterotoxigenic Escherichia coli, Degrades Intestinal Mucin. Infect. Immun. 2014, 82, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, A.; Wangdi, T.; Vickers, T.J.; Aaron, B.; Palmer, M.; Miller, M.J.; Kim, S.; Herring, C.; Simoes, R.; Crainic, J.A.; et al. Enterotoxigenic Escherichia coli Degrades the Host MUC2 Mucin Barrier to Facilitate Critical Pathogen-Enterocyte Interactions in Human Small Intestine. Infect. Immun. 2021, 90, 00572-21. [Google Scholar] [CrossRef]
- Sahl, J.W.; Rasko, D.A. Analysis of Global Transcriptional Profiles of Enterotoxigenic Escherichia coli Isolate E24377A. Infect. Immun. 2012, 80, 1232–1242. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Yin, X.; Yu, H.; Zhao, L.; Sabour, P.; Gong, J. Involvement of Quorum Sensing and Heat-Stable Enterotoxin a in Cell Damage Caused by a Porcine Enterotoxigenic Escherichia coli Strain. Infect. Immun. 2011, 79, 1688–1695. [Google Scholar] [CrossRef] [Green Version]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Bodero, M.D.; Munson, G.P. Cyclic AMP Receptor Protein-Dependent Repression of Heat-Labile Enterotoxin. Infect. Immun. 2009, 77, 791–798. [Google Scholar] [CrossRef] [Green Version]
- Kansal, R.; Rasko, D.A.; Sahl, J.W.; Munson, G.P.; Roy, K.; Luo, Q.; Sheikh, A.; Kuhne, K.J.; Fleckenstein, J.M. Transcriptional Modulation of Enterotoxigenic Escherichia coli Virulence Genes in Response to Epithelial Cell Interactions. Infect. Immun. 2013, 81, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Joffré, E.; von Mentzer, A.; Svennerholm, A.-M.; Sjöling, A. Identification of new heat-stable (STa) enterotoxin allele variants produced by human enterotoxigenic Escherichia coli (ETEC). Int. J. Med. Microbiol. 2016, 306, 586–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Enterotoxins | Infection |
|---|---|---|
| GIS26 | LT+pSTa+STb+ | PW |
| IMM07 | LT+pSTa−STb+ | PW |
| IMM10 | LT+pSTa+STb+ | N |
| IMM96 | LT+pSTa−STb+ | PW |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Cox, E.; Devriendt, B. Intestinal Epithelial Cells Modulate the Production of Enterotoxins by Porcine Enterotoxigenic E. coli Strains. Int. J. Mol. Sci. 2022, 23, 6589. https://doi.org/10.3390/ijms23126589
Wang H, Cox E, Devriendt B. Intestinal Epithelial Cells Modulate the Production of Enterotoxins by Porcine Enterotoxigenic E. coli Strains. International Journal of Molecular Sciences. 2022; 23(12):6589. https://doi.org/10.3390/ijms23126589
Chicago/Turabian StyleWang, Haixiu, Eric Cox, and Bert Devriendt. 2022. "Intestinal Epithelial Cells Modulate the Production of Enterotoxins by Porcine Enterotoxigenic E. coli Strains" International Journal of Molecular Sciences 23, no. 12: 6589. https://doi.org/10.3390/ijms23126589