
Effect of Passage Number of Conditioned Medium Collected from Equine Amniotic Fluid Mesenchymal Stem Cells: Porcine Oocyte Maturation and Embryo Development

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Effects of eAFMSC-CM Passage Numbers on Cumulus Expansion during IVM (Experiment 1)
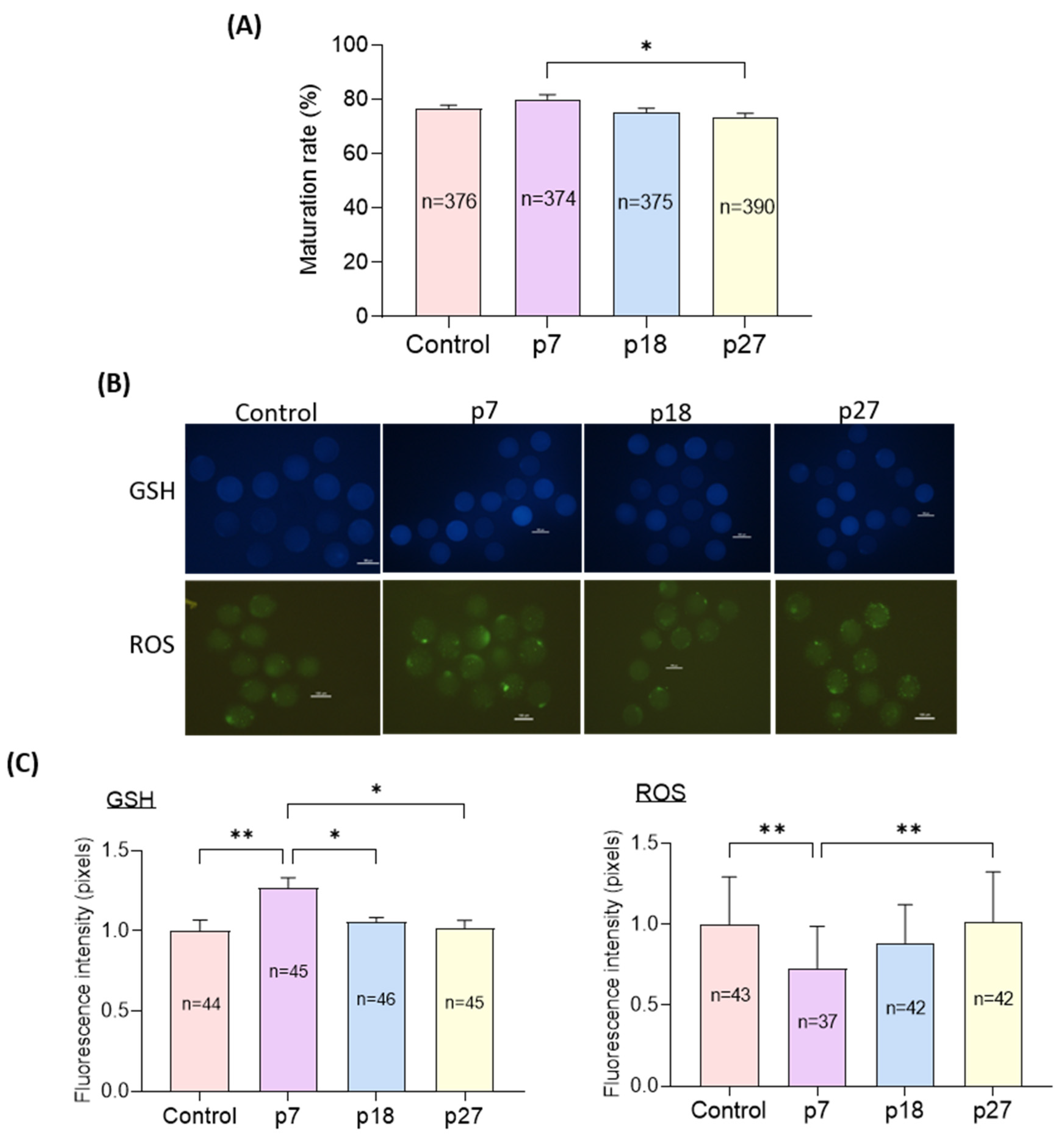
2.2. Effects of eAFMSC-CM Passages during IVM on Oocyte Cytoplasmic Maturation (Experiment 2)
2.3. Levels of Antioxidant Biomarkers in Fresh CM and IVM Medium with eAFMSC-CM (Experiment 3)
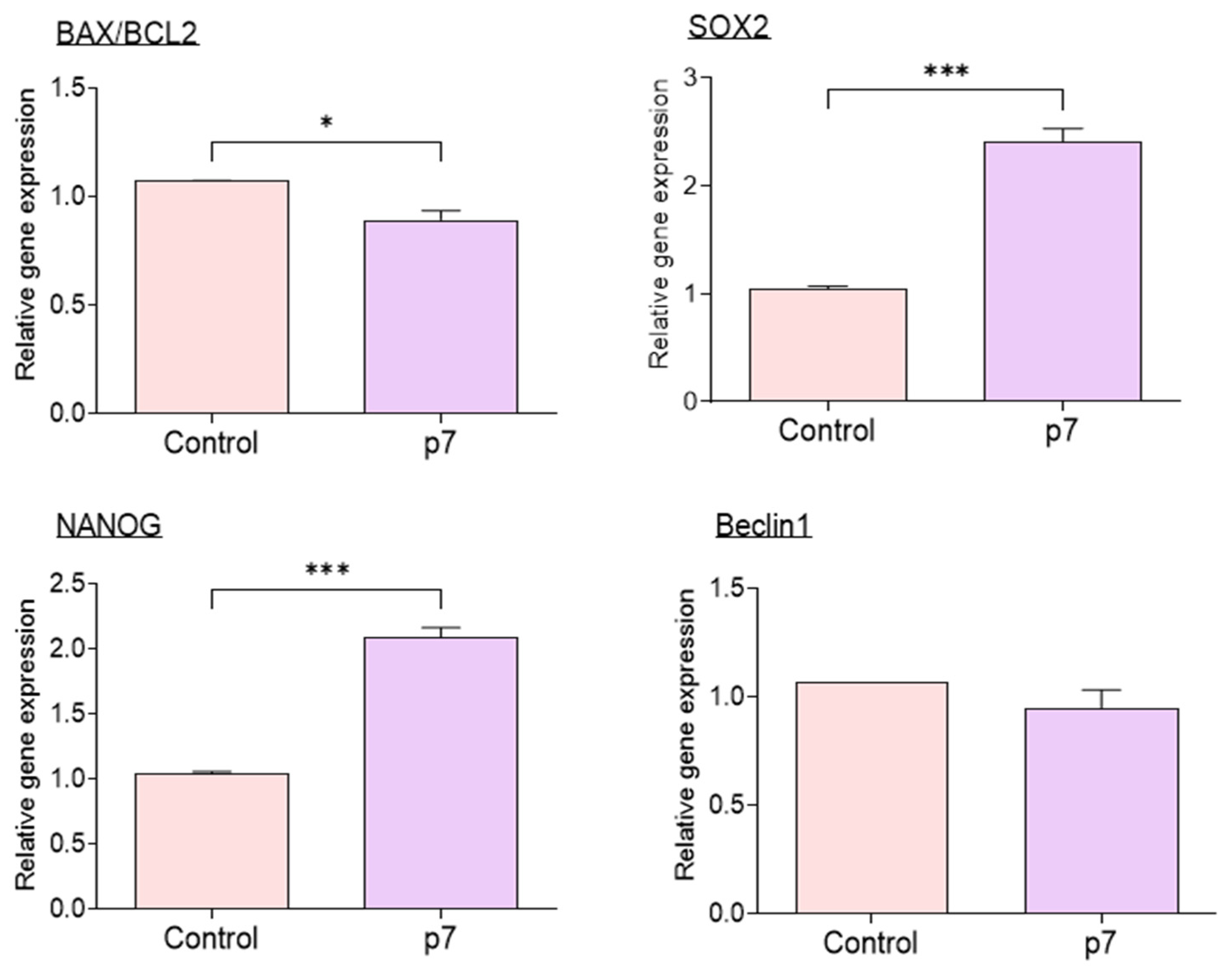
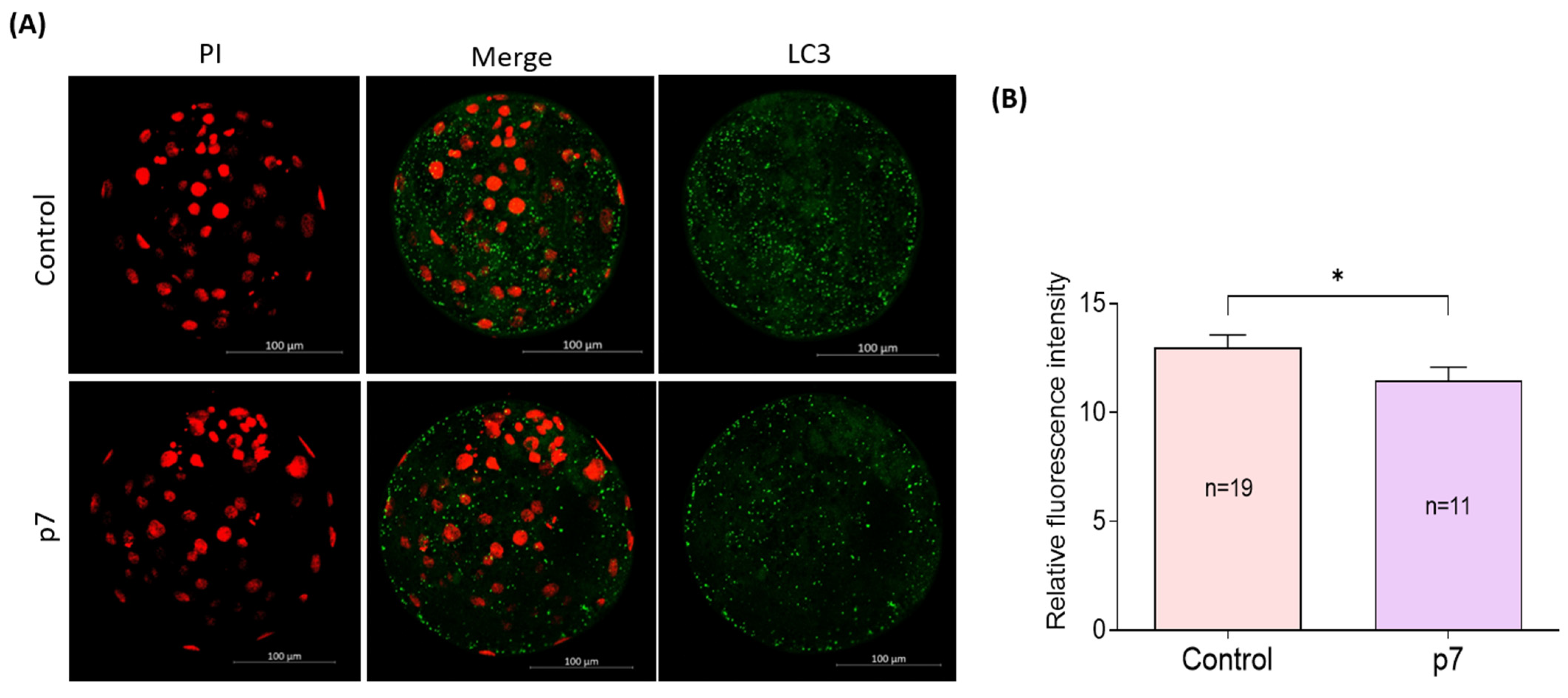
2.4. Effects of eAFMSC-CMp7 during IVM on Porcine Embryo Development after IVF (Experiment 4)
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Reagents and Chemicals
4.3. Preparation of Equine Amniotic Fluid Stem Cell-Conditioned Medium (eAFMSC-CM)
4.4. Maturation In Vitro
4.5. Evaluation of Cumulus Cell Expansion
4.6. Detection of Intracellular Glutathione (GSH) and ROS Levels
4.7. Assessment of Mitochondrial Distribution
4.8. Assessment of Cortical Granule Distribution
4.9. Antioxidant Enzyme Activity Assays
4.10. In Vitro Fertilization
4.11. Embryo Development and Total Blastocyst Counts
4.12. Immunofluorescence Staining
4.13. Analysis of Gene Expression by Quantitative Real-Time PCR
4.14. Experimental Design
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef] [Green Version]
- Prather, R.S. Pig genomics for biomedicine. Nat. Biotechnol. 2013, 31, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Catala, M.G.; Izquierdo, D.; Rodriguez-Prado, M.; Hammami, S.; Paramio, M.T. Effect of oocyte quality on blastocyst development after in vitro fertilization (IVF) and intracytoplasmic sperm injection (ICSI) in a sheep model. Fertil. Steril. 2012, 97, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tian, X.; Zhang, L.; He, C.; Ji, P.; Li, Y.; Tan, D.; Liu, G. Beneficial effect of resveratrol on bovine oocyte maturation and subsequent embryonic development after in vitro fertilization. Fertil. Steril. 2014, 101, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; Schultz, R.M.; O’Brien, M.; Chesnel, F. Relationship between the developmental programs controlling nuclear and cytoplasmic maturation of mouse oocytes. Dev. Biol. 1994, 164, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.E.; Thompson, J.G.; Armstrong, D.T.; Gilchrist, R.B. Effect of specific phosphodiesterase isoenzyme inhibitors during in vitro maturation of bovine oocytes on meiotic and developmental capacity. Biol. Reprod. 2004, 71, 1142–1149. [Google Scholar] [CrossRef] [Green Version]
- Reader, K.L.; Stanton, J.L.; Juengel, J.L. The Role of Oocyte Organelles in Determining Developmental Competence. Biology 2017, 6, 35. [Google Scholar] [CrossRef]
- Mao, L.; Lou, H.; Lou, Y.; Wang, N.; Jin, F. Behaviour of cytoplasmic organelles and cytoskeleton during oocyte maturation. Reprod. Biomed. Online 2014, 28, 284–299. [Google Scholar] [CrossRef] [Green Version]
- Cotterill, M.; Harris, S.E.; Collado Fernandez, E.; Lu, J.; Huntriss, J.D.; Campbell, B.K.; Picton, H.M. The activity and copy number of mitochondrial DNA in ovine oocytes throughout oogenesis in vivo and during oocyte maturation in vitro. Mol. Hum. Reprod. 2013, 19, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Harvey, A.J. Mitochondria in early development: Linking the microenvironment, metabolism and the epigenome. Reproduction 2019, 157, R159–R179. [Google Scholar] [CrossRef] [Green Version]
- Marchal, R.; Tomanek, M.; Terqui, M.; Mermillod, P. Effects of cell cycle dependent kinases inhibitor on nuclear and cytoplasmic maturation of porcine oocytes. Mol. Reprod. Dev. 2001, 60, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H. Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development. Int. J. Mol. Sci. 2021, 22, 579. [Google Scholar] [CrossRef]
- Menck, M.C.; Guyader-Joly, C.; Peynot, N.; Le Bourhis, D.; Lobo, R.B.; Renard, J.P.; Heyman, Y. Beneficial effects of Vero cells for developing IVF bovine eggs in two different coculture systems. Reprod. Nutr. Dev. 1997, 37, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adib, M.; Seifati, S.M.; Ashkezari, M.D.; Khoradmehr, A.; Rezaee-Ranjbar-Sardari, R.; Tahajjodi, S.S.; Aflatoonian, B. The effect of the human cumulus cells-conditioned medium on in vitro maturation of mouse oocyte: An experimental study. Int. J. Reprod. Biomed. 2020, 18, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Akbari, H.; Eftekhar Vaghefi, S.H.; Shahedi, A.; Habibzadeh, V.; Mirshekari, T.R.; Ganjizadegan, A.; Mollaei, H.; Ahmadi, M.; Nematollahi-Mahani, S.N. Mesenchymal Stem Cell-Conditioned Medium Modulates Apoptotic and Stress-Related Gene Expression, Ameliorates Maturation and Allows for the Development of Immature Human Oocytes after Artificial Activation. Genes 2017, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H. Effects of Human Endothelial Progenitor Cell and Its Conditioned Medium on Oocyte Development and Subsequent Embryo Development. Int. J. Mol. Sci. 2020, 21, 7983. [Google Scholar] [CrossRef]
- Cantinieaux, D.; Quertainmont, R.; Blacher, S.; Rossi, L.; Wanet, T.; Noel, A.; Brook, G.; Schoenen, J.; Franzen, R. Conditioned medium from bone marrow-derived mesenchymal stem cells improves recovery after spinal cord injury in rats: An original strategy to avoid cell transplantation. PLoS ONE 2013, 8, e69515. [Google Scholar] [CrossRef] [PubMed]
- Saleem, R.; Mohamed-Ahmed, S.; Elnour, R.; Berggreen, E.; Mustafa, K.; Al-Sharabi, N. Conditioned Medium from Bone Marrow Mesenchymal Stem Cells Restored Oxidative Stress-Related Impaired Osteogenic Differentiation. Int. J. Mol. Sci. 2021, 22, 13458. [Google Scholar] [CrossRef]
- De Coppi, P.; Bartsch, G., Jr.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of amniotic stem cell lines with potential for therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef]
- Tsai, M.S.; Lee, J.L.; Chang, Y.J.; Hwang, S.M. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef]
- Prusa, A.R.; Marton, E.; Rosner, M.; Bettelheim, D.; Lubec, G.; Pollack, A.; Bernaschek, G.; Hengstschlager, M. Neurogenic cells in human amniotic fluid. Am. J. Obstet. Gynecol. 2004, 191, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Siegel, N.; Rosner, M.; Hanneder, M.; Valli, A.; Hengstschläger, M. Stem cells in amniotic fluid as new tools to study human genetic diseases. Stem Cell Rev. Rep. 2007, 3, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.A.; Choi, H.S.; Kim, K.J.; Lee, D.S.; Lee, J.H.; Park, J.Y.; Kim, E.Y.; Li, X.; Oh, H.Y.; Lee, D.S.; et al. Isolation of canine mesenchymal stem cells from amniotic fluid and differentiation into hepatocyte-like cells. Vitr. Cell. Dev. Biology. Anim. 2013, 49, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Gatti, M.; Zavatti, M.; Beretti, F.; Giuliani, D.; Vandini, E.; Ottani, A.; Bertucci, E.; Maraldi, T. Oxidative Stress in Alzheimer’s Disease: In Vitro Therapeutic Effect of Amniotic Fluid Stem Cells Extracellular Vesicles. Oxidative Med. Cell. Longev. 2020, 2020, 2785343. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhao, L.; Deng, H.; Wei, M.; Li, J.; Xu, K. Immune tolerance of amniotic fluid stem cell-induced rat kidney graft and influences on oxidative stress. Transplant. Proc. 2013, 45, 3394–3401. [Google Scholar] [CrossRef] [PubMed]
- Al-Husseiny, F.; Sobh, M.A.; Ashour, R.H.; Foud, S.; Medhat, T.; El-Gilany, A.H.; Elghannam, D.; Abdel-Ghaffar, H.; Saad, M.A.; Sobh, M. Amniotic Fluid-Derived Mesenchymal Stem Cells Cut Short the Acuteness of Cisplatin-Induced Nephrotoxicity in Sprague-Dawley Rats. Int. J. Stem Cells 2016, 9, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Seo, M.S.; Kang, J.G.; Chae, J.S.; Kang, K.S. Isolation and characterization of equine amniotic fluid-derived multipotent stem cells. Cytotherapy 2011, 13, 341–349. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Leng, Y.; Panaitescu, B.; Silva, P.; Faro, J.; Alhousseini, A.; Gill, N.; et al. The immunophenotype of amniotic fluid leukocytes in normal and complicated pregnancies. Am. J. Reprod. Immunol. 2018, 79, e12827. [Google Scholar] [CrossRef] [Green Version]
- Fehrer, C.; Lepperdinger, G. Mesenchymal stem cell aging. Exp. Gerontol. 2005, 40, 926–930. [Google Scholar] [CrossRef]
- Kretlow, J.D.; Jin, Y.Q.; Liu, W.; Zhang, W.J.; Hong, T.H.; Zhou, G.; Baggett, L.S.; Mikos, A.G.; Cao, Y. Donor age and cell passage affects differentiation potential of murine bone marrow-derived stem cells. BMC Cell Biol. 2008, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Bonab, M.M.; Alimoghaddam, K.; Talebian, F.; Ghaffari, S.H.; Ghavamzadeh, A.; Nikbin, B. Aging of mesenchymal stem cell in vitro. BMC Cell Biol. 2006, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sareen, N.; Sequiera, G.L.; Chaudhary, R.; Abu-El-Rub, E.; Chowdhury, S.R.; Sharma, V.; Surendran, A.; Moudgil, M.; Fernyhough, P.; Ravandi, A.; et al. Early passaging of mesenchymal stem cells does not instigate significant modifications in their immunological behavior. Stem Cell Res. Ther. 2018, 9, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Amano, T.; Shimizu, H. Roles of gap junctional communication of cumulus cells in cytoplasmic maturation of porcine oocytes cultured in vitro. Biol. Reprod. 2000, 62, 913–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chian, R.C.; Niwa, K.; Sirard, M.A. Effects of cumulus cells on male pronuclear formation and subsequent early development of bovine oocytes in vitro. Theriogenology 1994, 41, 1499–1508. [Google Scholar] [CrossRef]
- Palta, P.; Chauhan, M.S. Laboratory production of buffalo (Bubalus bubalis) embryos. Reprod. Fertil. Dev. 1998, 10, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Filali, M.; Frydman, N.; Belot, M.P.; Hesters, L.; Gaudin, F.; Tachdjian, G.; Emilie, D.; Frydman, R.; Machelon, V. Oocyte in-vitro maturation: BCL2 mRNA content in cumulus cells reflects oocyte competency. Reprod. Biomed. Online 2009, 19 (Suppl. 4), 4309. [Google Scholar]
- Kim, E.H.; Ridlo, M.R.; Lee, B.C.; Kim, G.A. Melatonin-Nrf2 Signaling Activates Peroxisomal Activities in Porcine Cumulus Cell-Oocyte Complexes. Antioxidants 2020, 9, 1080. [Google Scholar] [CrossRef]
- Gwarzo, M.Y. Nrf2 transcription factor gene regulates basal transcription of mitochondrial superoxide dismutase enzyme in mouse brain. Afr. J. Biotechnol. 2009, 8, 5169–5172. [Google Scholar]
- Hansen, J.M.; Harris, C. Glutathione during embryonic development. Biochim. Biophys. Acta 2015, 1850, 1527–1542. [Google Scholar] [CrossRef]
- Luberda, Z. The role of glutathione in mammalian gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar] [PubMed]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol. Cells 2011, 32, 491–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatemoto, H.; Ootaki, K.; Shigeta, K.; Muto, N. Enhancement of Developmental Competence after In Vitro Fertilization of Porcine Oocytes by Treatment with Ascorbic Acid 2-O-α-Glucoside During In Vitro Maturation1. Biol. Reprod. 2001, 65, 1800–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, F.; Romero, S.; De Vos, M.; Verheyen, G.; Smitz, J. Human cumulus-enclosed germinal vesicle oocytes from early antral follicles reveal heterogeneous cellular and molecular features associated with in vitro maturation capacity. Hum. Reprod. 2015, 30, 1396–1409. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, E.M.; Vireque, A.A.; Adona, P.R.; Meirelles, F.V.; Ferriani, R.A.; Navarro, P.A. Cytoplasmic maturation of bovine oocytes: Structural and biochemical modifications and acquisition of developmental competence. Theriogenology 2009, 71, 836–848. [Google Scholar] [CrossRef]
- Zeng, J.; Jiang, M.; Wu, X.; Diao, F.; Qiu, D.; Hou, X.; Wang, H.; Li, L.; Li, C.; Ge, J.; et al. SIRT4 is essential for metabolic control and meiotic structure during mouse oocyte maturation. Aging Cell 2018, 17, e12789. [Google Scholar] [CrossRef]
- Burkart, A.D.; Xiong, B.; Baibakov, B.; Jiménez-Movilla, M.; Dean, J. Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy. J. Cell Biol. 2012, 197, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sun, Q.Y. Evaluation of oocyte quality: Morphological, cellular and molecular predictors. Reprod. Fertil. Dev. 2007, 19, 1–12. [Google Scholar] [CrossRef]
- Wang, Q.; Frolova, A.I.; Purcell, S.; Adastra, K.; Schoeller, E.; Chi, M.M.; Schedl, T.; Moley, K.H. Mitochondrial dysfunction and apoptosis in cumulus cells of type I diabetic mice. PLoS ONE 2010, 5, e15901. [Google Scholar] [CrossRef] [Green Version]
- Galeati, G.; Modina, S.; Lauria, A.; Mattioli, M. Follicle somatic cells influence pig oocyte penetrability and cortical granule distribution. Mol. Reprod. Dev. 1991, 29, 40–46. [Google Scholar] [CrossRef]
- Gu, Y.; Li, T.; Ding, Y.; Sun, L.; Tu, T.; Zhu, W.; Hu, J.; Sun, X. Changes in mesenchymal stem cells following long-term culture in vitro. Mol. Med. Rep. 2016, 13, 5207–5215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedi, M.; Parham, A.; Dehghani, H.; Mehrjerdi, H.K. Stemness Signature of Equine Marrow-derived Mesenchymal Stem Cells. Int. J. Stem Cells 2017, 10, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraniak, P.R.; McDevitt, T.C. Stem cell paracrine actions and tissue regeneration. Regen. Med. 2010, 5, 121–143. [Google Scholar] [CrossRef] [Green Version]
- Maguire, G.; Biology, I. Stem cell therapy without the cells. Commun. Integr. Biol. 2013, 6, e26631. [Google Scholar] [CrossRef] [PubMed]
- Kongsuphol, P.; Mukda, S.; Nopparat, C.; Villarroel, A.; Govitrapong, P. Melatonin attenuates methamphetamine-induced deactivation of the mammalian target of rapamycin signaling to induce autophagy in SK-N-SH cells. J. Pineal Res. 2009, 46, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Yeh, P.Y.; Lu, Y.S.; Hsu, C.H.; Chen, K.F.; Lee, W.C.; Feng, W.C.; Chen, C.S.; Kuo, M.L.; Cheng, A.L. OSU-03012, a novel celecoxib derivative, induces reactive oxygen species-related autophagy in hepatocellular carcinoma. Cancer Res. 2008, 68, 9348–9357. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Xu, J.; Lin, Y.; Zhao, X.; Liu, X.; Chi, Z. Autophagy is upregulated in rats with status epilepticus and partly inhibited by Vitamin E. Biochem. Biophys. Res. Commun. 2009, 379, 949–953. [Google Scholar] [CrossRef]
- Zhu, C.C.; Zhang, Y.; Duan, X.; Han, J.; Sun, S.C. Toxic effects of HT-2 toxin on mouse oocytes and its possible mechanisms. Arch. Toxicol. 2016, 90, 1495–1505. [Google Scholar] [CrossRef]
- Jia, Z.Z.; Zhang, J.W.; Zhou, D.; Xu, D.Q.; Feng, X.Z. Deltamethrin exposure induces oxidative stress and affects meiotic maturation in mouse oocyte. Chemosphere 2019, 223, 704–713. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, H.R.; Kim, M.G.; Lee, J.S.; Jin, S.J.; Lee, H.T. The effect of poly(ADP-ribosyl)ation inhibition on the porcine cumulus-oocyte complex during in vitro maturation. Biochem. Biophys. Res. Commun. 2017, 483, 752–758. [Google Scholar] [CrossRef]
- Park, M.R.; Gupta, M.K.; Lee, H.R.; Das, Z.C.; Uhm, S.J.; Lee, H.T. Possible involvement of Class III phosphatidylinositol-3-kinase in meiotic progression of porcine oocytes beyond germinal vesicle stage. Theriogenology 2011, 75, 940–950. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Product Size (bp) | NCBI Accession No. |
|---|---|---|---|
| GAPDH | F: gtc ggt tgt gga tct gac ct | 207 | NM_001206359.1 |
| R: ttg acg aag tgg tcg ttg ag | |||
| BAX | F: ggt cgc gct ttt cta ctt tg | 111 | XM_003127290 |
| R: cga tct cga agg aag tcc ag | |||
| BCL2 | F: aaa caa tgc agc agc tga ga | 85 | NM_214285 |
| R: aac cac ccc agc tag agt ca | |||
| HAS2 | F: cag gga caa ttc agc cac tt | 100 | NM_214053.1 |
| R: ggt gac atg ttg gga gct tt | |||
| TNFAIP6 | F: gaa gca cgg tcg ggc aag | 141 | NM_001159607.1 |
| R: cat cca ccc agc agc aca g | |||
| PTGS2 | F: tcc tga aca cct ccg ctt tg | 147 | NM_214321.1 |
| R: agc cgt tca tcg tcc cat tc | |||
| Cx37 | F: cac cct gtc cct acc tcg ta | 101 | NM_001244224.1 |
| R: gag cac cag gga gat gag tc | |||
| Cx43 | F: gct ggt cgt atc ctt ggt gt | 83 | NM_001244212.1 |
| R: tct ttc cct tca cac gat cc | |||
| SOD2 | F: tgg agg cca cat caa tca ta | 136 | NM_214127.2 |
| R: agc ggt caa ctt ctc ctt ga | |||
| NRF-2 | F: gtg cct ata agt ccc ggt ca | 108 | MH101365.1 |
| R: atg cag agc ttt tgc cct ta | |||
| BECN1 | F: gat agt ggc gga aaa tct cg | 160 | NM_001044530.1 |
| R: cat ctg ggc ata acg cat ct | |||
| SOX2 | F: cgc aga cct aca tga acg | 103 | NM_001123197.1 |
| R: tcg gac ttg acc act gag | |||
| NANOG | F: ccc gaa gca tcc att tcc ag | 171 | NM_001129971.1 |
| R: tgt gga aga atc agg gct gt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, A.; Oh, H.J.; Ji, K.; Choi, E.M.; Kim, D.; Kim, E.; Kim, M.K. Effect of Passage Number of Conditioned Medium Collected from Equine Amniotic Fluid Mesenchymal Stem Cells: Porcine Oocyte Maturation and Embryo Development. Int. J. Mol. Sci. 2022, 23, 6569. https://doi.org/10.3390/ijms23126569
Park A, Oh HJ, Ji K, Choi EM, Kim D, Kim E, Kim MK. Effect of Passage Number of Conditioned Medium Collected from Equine Amniotic Fluid Mesenchymal Stem Cells: Porcine Oocyte Maturation and Embryo Development. International Journal of Molecular Sciences. 2022; 23(12):6569. https://doi.org/10.3390/ijms23126569
Chicago/Turabian StylePark, Ahyoung, Hyun Ju Oh, Kukbin Ji, Eunha Miri Choi, Dongern Kim, Eunyoung Kim, and Min Kyu Kim. 2022. "Effect of Passage Number of Conditioned Medium Collected from Equine Amniotic Fluid Mesenchymal Stem Cells: Porcine Oocyte Maturation and Embryo Development" International Journal of Molecular Sciences 23, no. 12: 6569. https://doi.org/10.3390/ijms23126569