
Neurosecretory Protein GL Promotes Normotopic Fat Accumulation in Male ICR Mice


Abstract
:1. Introduction
2. Results
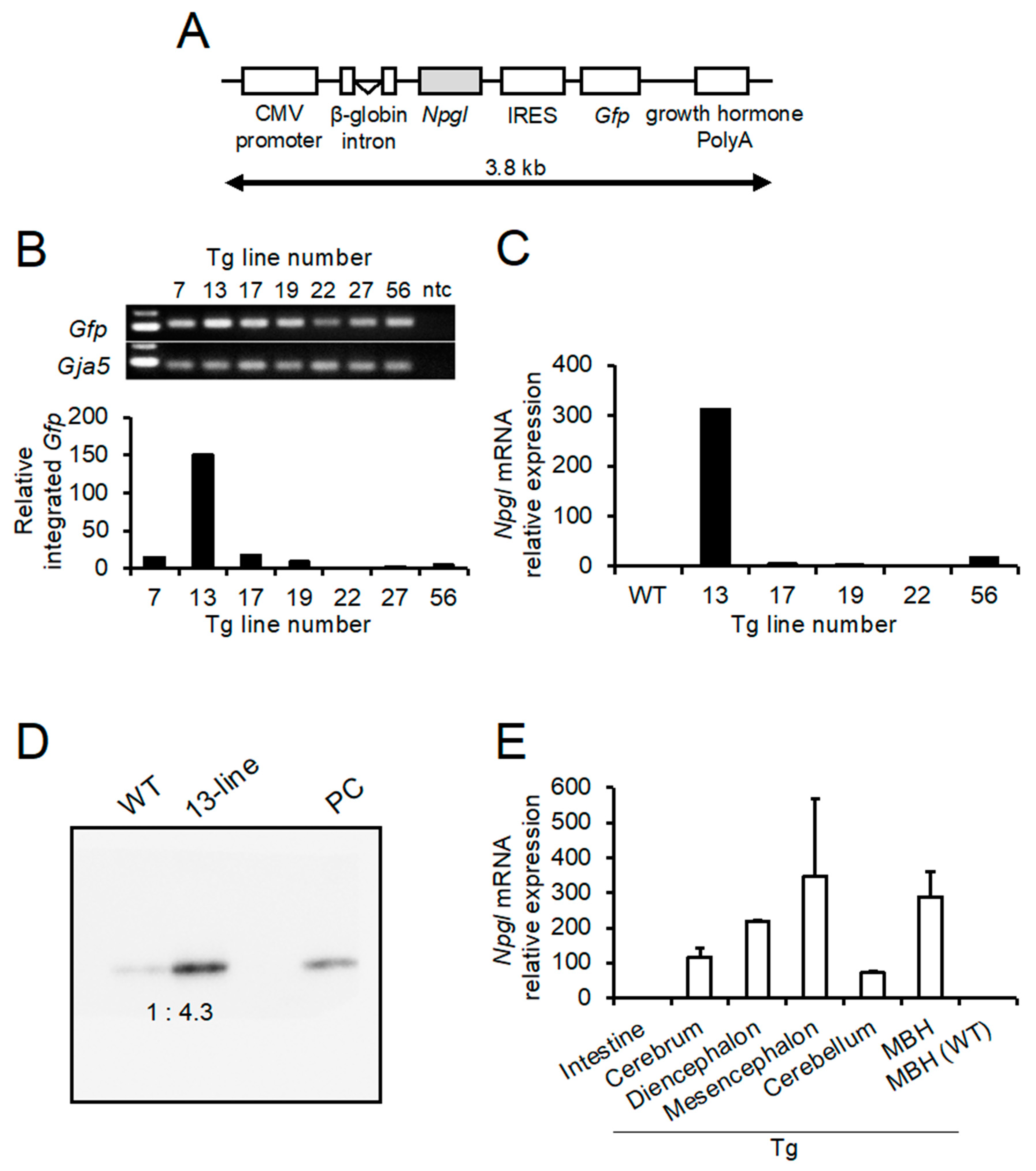
2.1. Production of Tg Mouse
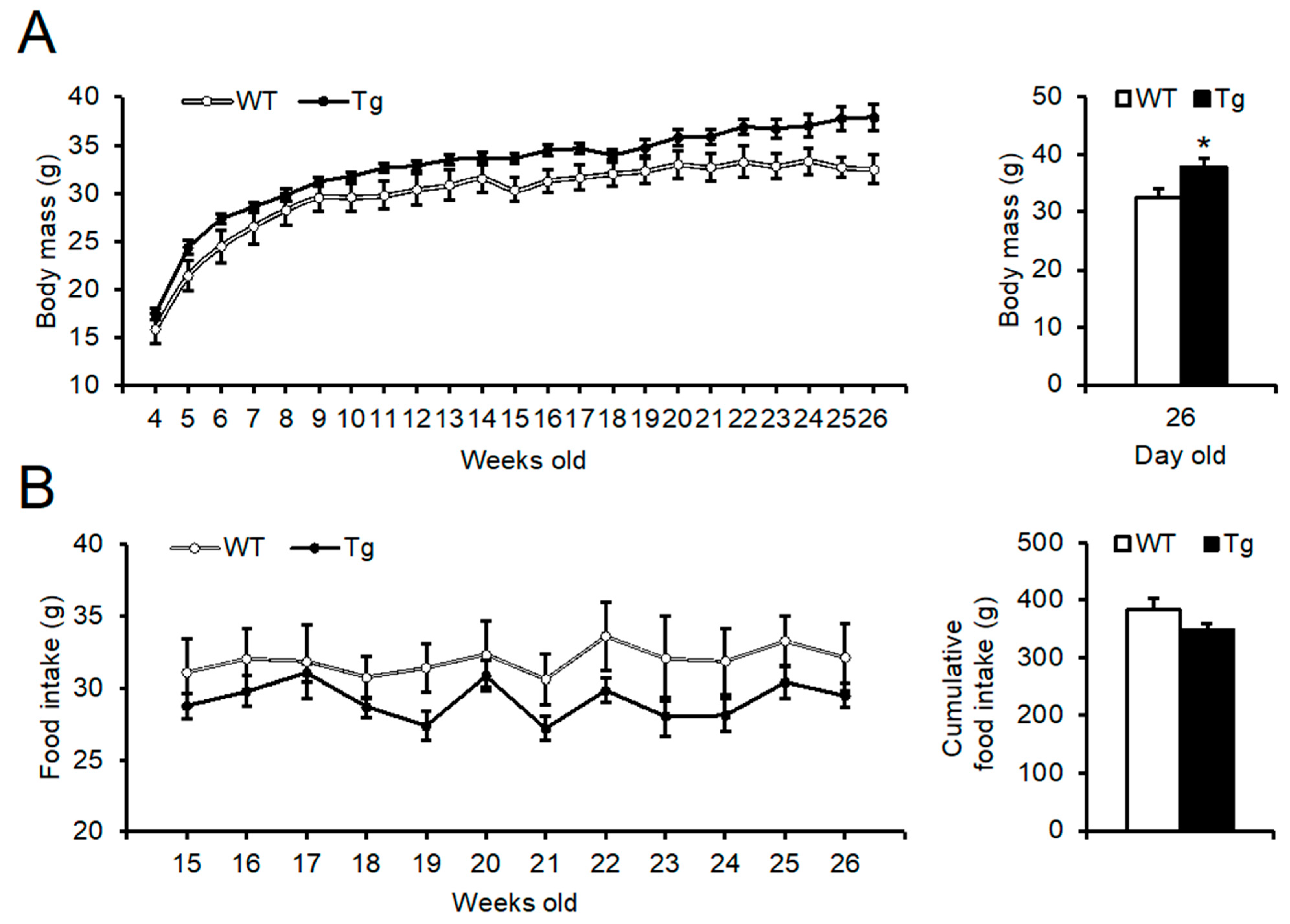
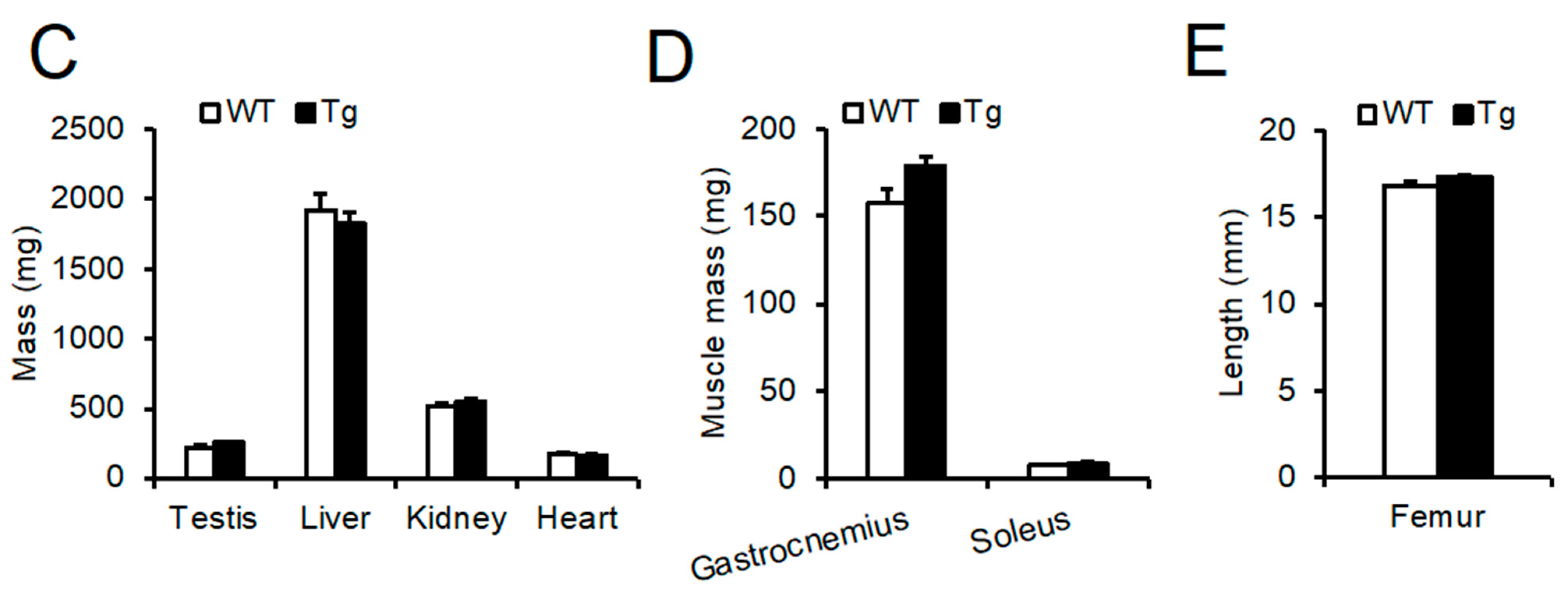
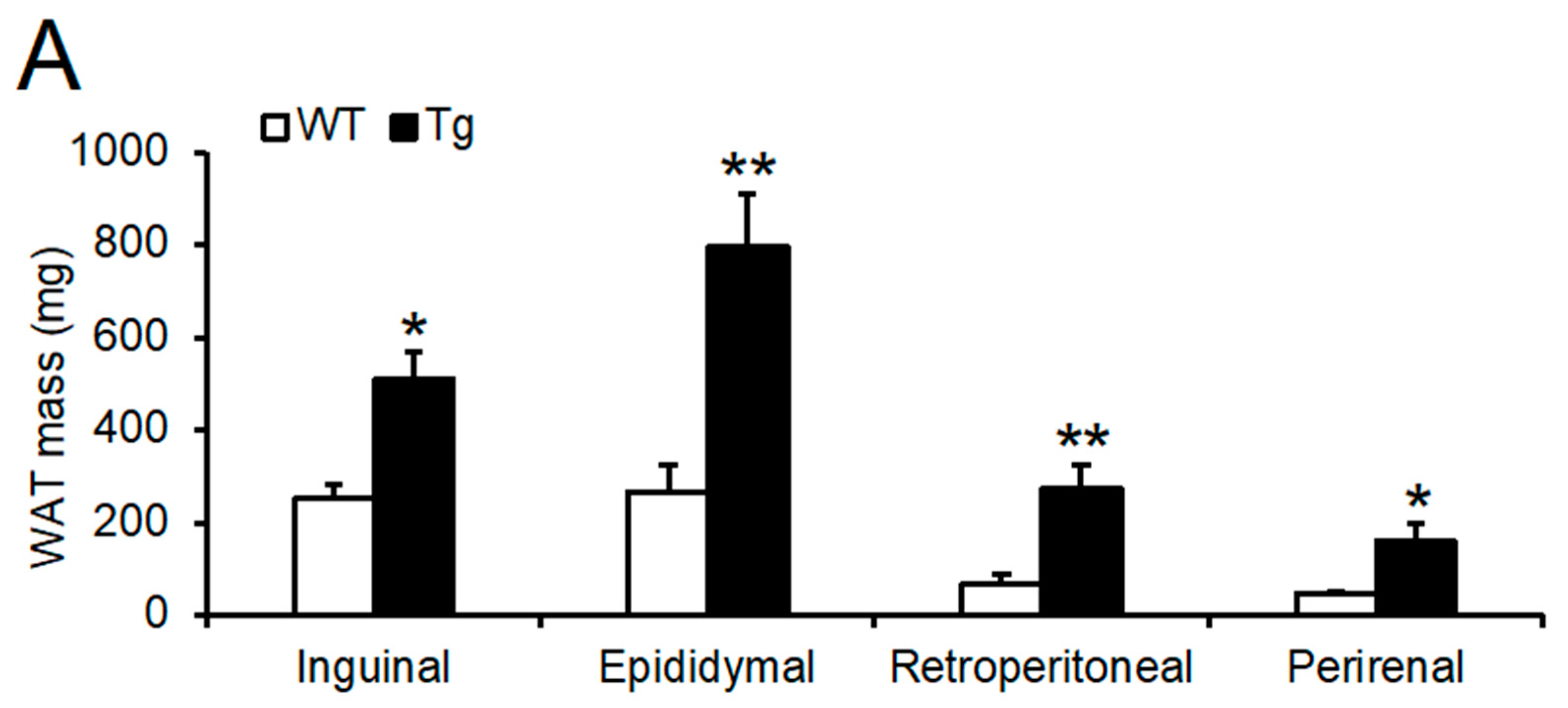
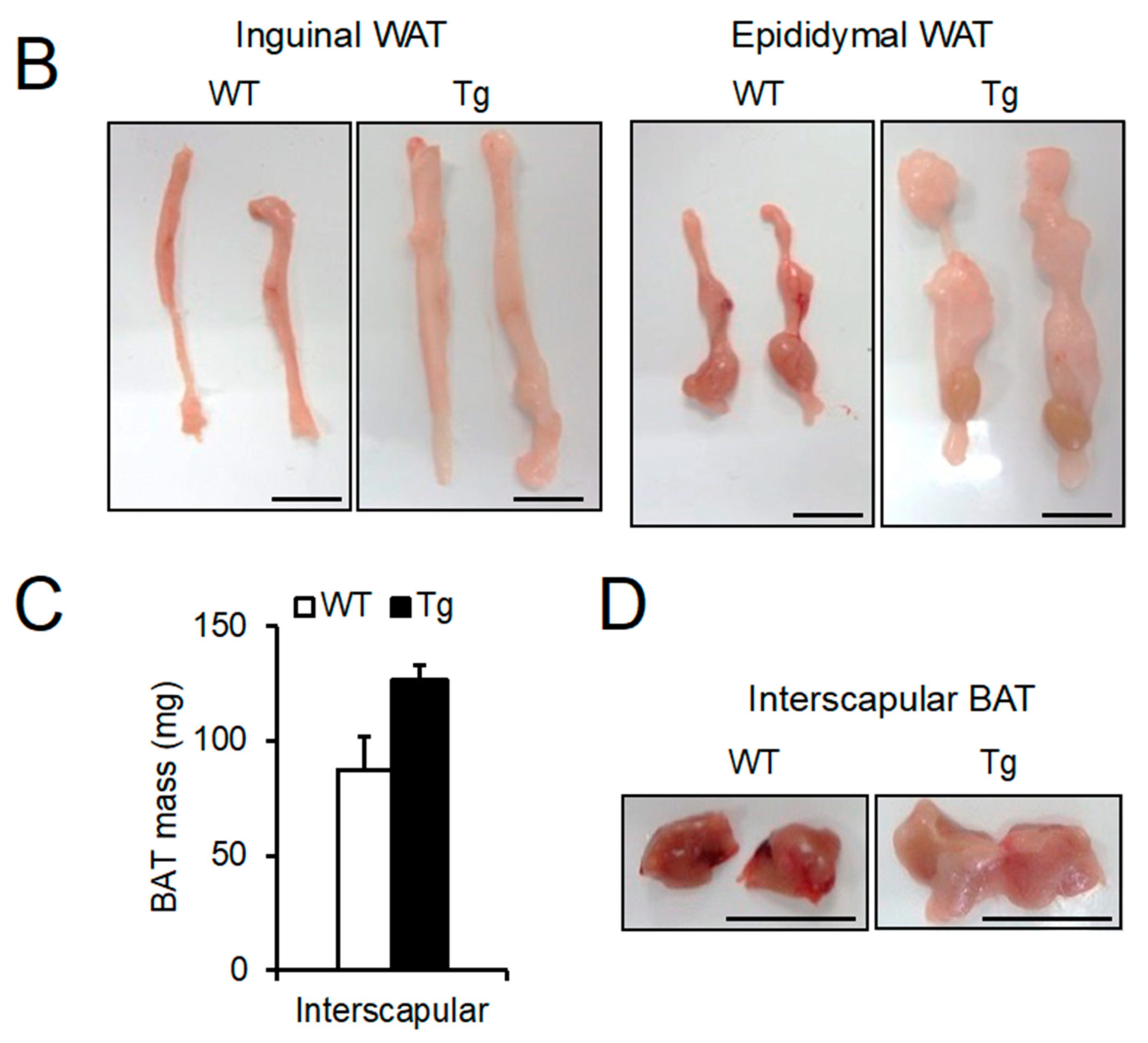
2.2. Phenotypic Analysis of Body Mass, Food Intake, and Peripheral Tissues
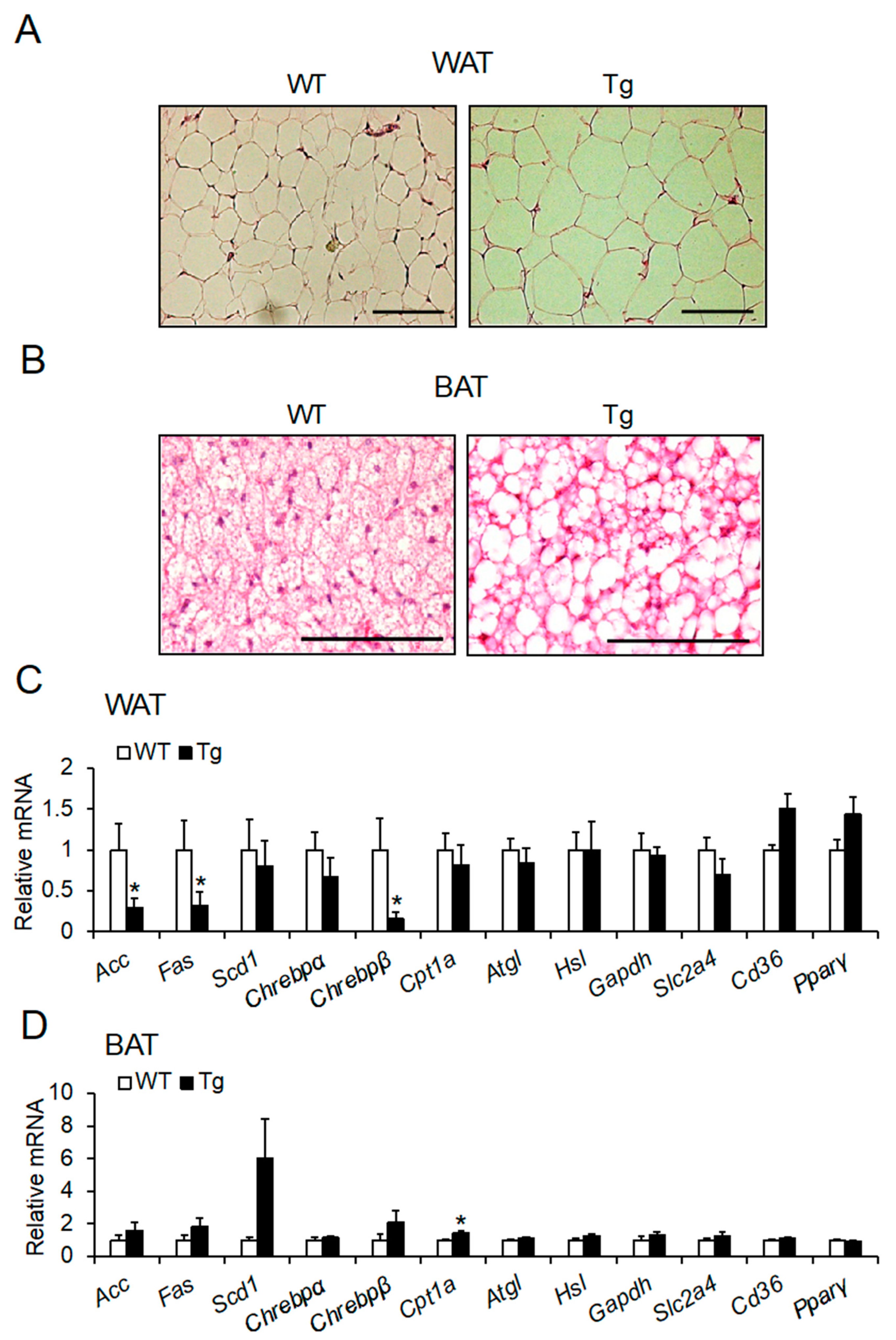
2.3. Lipid Droplet Hypertrophy in Adipose Tissues
2.4. mRNA Expression of Lipid Metabolic Factors in Adipose Tissues
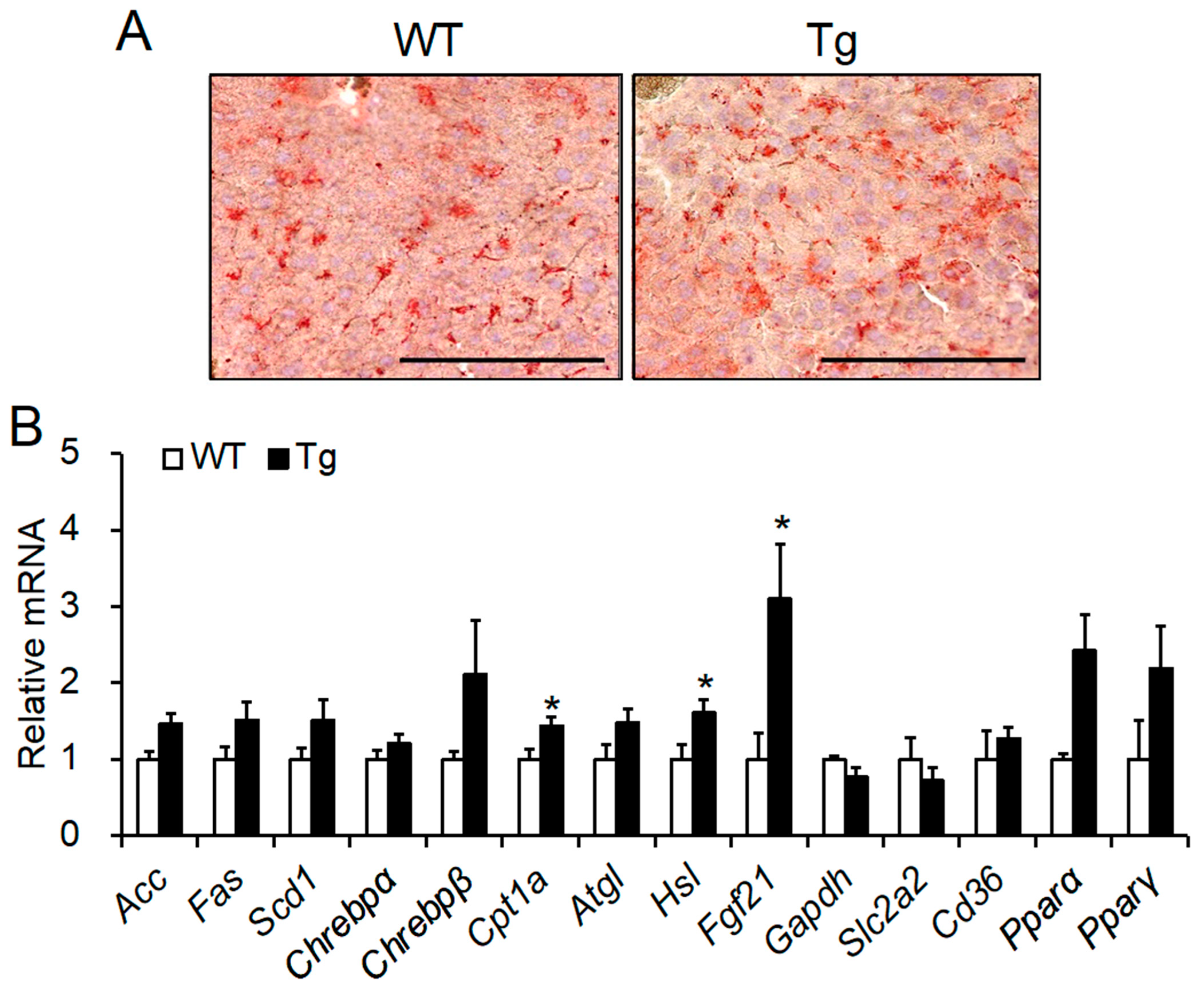
2.5. Histological Analysis and mRNA Expression of Lipid Metabolic Factors in the Liver
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Generation of Transgenic Mouse Overexpressing Npgl
4.3. Selection of the Line of Npgl Tg Mice
4.4. Tissue Collection
4.5. qRT-PCR
4.6. Western Blot Analysis
4.7. Hematoxylin and Eosin Staining
4.8. Oil Red O Staining
4.9. Serum Biochemical Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA carboxylase |
| ACTB | β-Actin |
| AgRP | Agouti-related protein |
| ATGL | Adipose triglyceride lipase |
| α-MSH | α-melanocyte-stimulating hormone |
| BAT | Brown adipose tissue |
| CART | Cocaine- and amphetamine-regulated transcript |
| CD36 | Cluster of differentiation 36 |
| ChREBPα | Carbohydrate-responsive element-binding protein α |
| ChREBPβ | Carbohydrate-responsive element-binding protein β |
| CMV | Cytomegalovirus |
| COVID-19 | Coronavirus disease 2019 |
| CPT1a | Carnitine palmitoyltransferase 1a |
| CRH | Corticotropin-releasing hormone |
| DIO | Diet-induced obesity |
| DIO2 | Type 2 iodothyronine deiodinase |
| FAS | Fatty acid synthase |
| FGF21 | Fibroblast growth factor 21 |
| FSH | Follicle stimulating hormone |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GFP | Green fluorescent protein |
| GH | Growth hormone |
| GHRH | Growth hormone releasing hormone |
| GJA5 | Gap junction alpha-5 protein |
| HFD | High-fat diet |
| HSL | Hormone-sensitive lipase |
| i.c.v. | Intracerebroventricular |
| IRES | Internal ribosome entry sites |
| LH | Luteinizing hormone |
| MBH | Mediobasal hypothalamus |
| MCH | Melanin-concentrating hormone |
| NPGL | Neurosecretory protein GL |
| NPY | Neuropeptide Y |
| ODS | Octadecylsilane |
| PFA | Paraformaldehyde |
| PC | Positive control |
| PGC1α | Peroxisome proliferator-activated receptor-γ coactivator 1α |
| POMC | Proopiomelanocortin |
| PPARα | Peroxisome proliferator-activated receptor α |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| PRL | Prolactin |
| qRT-PCR | Quantitative reverse transcriptase PCR |
| RPS18 | Ribosomal protein S18 |
| SCD1 | Stearoyl-CoA desaturase 1 |
| SGLT2 | Sodium-glucose cotransporter 2 |
| SLC2A2 | Glucose transporter 2 |
| SLC2A4 | Glucose transporter 4 |
| SST | Somatostatin |
| Tg | Transgenic |
| TRH | Thyrotropin-releasing hormone |
| TSH | Thyroid stimulating hormone |
| UCP1 | Uncoupling protein 1 |
| WAT | White adipose tissue |
| WT | Wild-type |
References
- Stefan, N.; Birkenfeld, A.L.; Schulze, M.B. Global pandemics interconnected—Obesity, impaired metabolic health and COVID-19. Nat. Rev. Endocrinol. 2021, 17, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.; Choi, M.-S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simha, V. Management of hypertriglyceridemia. BMJ 2020, 371, m3109. [Google Scholar] [CrossRef] [PubMed]
- Seravalle, G.; Grassi, G. Obesity and hypertension. Pharmacol. Res. 2017, 122, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Waterson, M.J.; Horvath, T.L. Neuronal regulation of energy homeostasis: Beyond the hypothalamus and feeding. Cell Metab. 2015, 22, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, B.G. Neuropeptide Y in multiple hypothalamic sites controls eating behavior, endocrine, and autonomic systems for body energy balance. In The Biology of Neuropeptide Y and Related Peptides. Contemporary Neuroscience; Colmers, W.F., Wahlestedt, C., Eds.; Humana Press: Totowa, NJ, USA, 1993; pp. 457–509. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Crawley, J.N. The role of galanin in feeding behavior. Neuropeptides 1999, 33, 369–375. [Google Scholar] [CrossRef]
- Tritos, N.A.; Maratos-Flier, E. Two important systems in energy homeostasis: Melanocortins and melanin-concentrating hormone. Neuropeptides 1999, 33, 339–349. [Google Scholar] [CrossRef]
- Yaswen, L.; Diehl, N.; Brennan, M.B.; Hochgeschwender, U. Obesity in the mouse model of pro-opiomelanocortin deficiency responds to peripheral melanocortin. Nat. Med. 1999, 5, 1066–1070. [Google Scholar] [CrossRef]
- Arase, K.; Shargill, N.S.; Bray, G.A. Effects of corticotropin releasing factor on genetically obese (fatty) rats. Physiol. Behav. 1989, 45, 565–570. [Google Scholar] [CrossRef]
- Kristensen, P.; Judge, M.E.; Thim, L.; Ribel, U.; Christjansen, K.N.; Wulff, B.S.; Clausen, J.T.; Jensen, P.B.; Madsen, O.D.; Vrang, N.; et al. Hypothalamic CART is a new anorectic peptide regulated by leptin. Nature 1998, 393, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.B.S. Direct and indirect effects of leptin on adipocyte metabolism. Biochim. Biophys. Acta 2014, 1842, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, T.; OHare, J.; Diggs-Andrews, K.; Schweiger, M.; Cheng, B.; Lindtner, C.; Zielinski, E.; Vempati, P.; Su, K.; Dighe, S.; et al. Brain insulin controls adipose tissue lipolysis and lipogenesis. Cell Metab. 2011, 13, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkotou, E.; Jeon, J.Y.; Wang, X.; Marino, F.E.; Carlson, M.; Trombly, D.J.; Maratos-Flier, E. Mice with MCH ablation resist diet-induced obesity through strain-specific mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R117–R124. [Google Scholar] [CrossRef]
- Hui, S.T.; Kurt, Z.; Tuominen, I.; Norheim, F.; Davis, R.C.; Pan, C.; Dirks, D.L.; Magyar, C.E.; French, S.W.; Chella Krishnan, K.; et al. The genetic architecture of diet-induced hepatic fibrosis in mice. Hepatology 2018, 68, 2182–2196. [Google Scholar] [CrossRef] [PubMed]
- Nehrenberg, D.L.; Hua, K.; Estrada-Smith, D.; Garland, T.; Pomp, D. Voluntary exercise and its effects on body composition depend on genetic selection history. Obesity 2009, 17, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Draisma, H.H.M.; Pool, R.; Kobl, M.; Jansen, R.; Petersen, A.K.; Vaarhorst, A.A.M.; Yet, I.; Haller, T.; Demirkan, A.; Esko, T.; et al. Genome-wide association study identifies novel genetic variants contributing to variation in blood metabolite levels. Nat. Commun. 2015, 6, 7208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukena, K.; Iwakoshi-Ukena, E.; Taniuchi, S.; Bessho, Y.; Maejima, S.; Masuda, K.; Shikano, K.; Kondo, K.; Furumitsu, M.; Tachibana, T. Identification of a cDNA encoding a novel small secretory protein, neurosecretory protein GL, in the chicken hypothalamic infundibulum. Biochem. Biophys. Res. Commun. 2014, 446, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, D.; Shikano, K.; Saito, T.; Iwakoshi-Ukena, E.; Furumitsu, M.; Ochi, Y.; Sato, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Neurosecretory protein GL, a hypothalamic small secretory protein, participates in energy homeostasis in male mice. Endocrinology 2017, 158, 1120–1129. [Google Scholar] [CrossRef]
- Iwakoshi-Ukena, E.; Shikano, K.; Kondo, K.; Taniuchi, S.; Furumitsu, M.; Ochi, Y.; Sasaki, T.; Okamoto, S.; Bentley, G.E.; Kriegsfeld, L.J.; et al. Neurosecretory protein GL stimulates food intake, de novo lipogenesis, and onset of obesity. eLife 2017, 6, e28527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukena, K. Avian and murine neurosecretory protein GL participates in the regulation of feeding and energy metabolism. Gen. Comp. Endocrinol. 2018, 260, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Shikano, K.; Iwakoshi-Ukena, E.; Kato, M.; Furumitsu, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Neurosecretory protein GL induces fat accumulation in chicks. Front. Endocrinol. 2019, 10, 392. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Iwakoshi-Ukena, E.; Fukumura, K.; Shikano, K.; Furumitsu, M.; Morishita, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Hypothalamic overexpression of neurosecretory protein GL leads to obesity in male C57BL/6J mice. Neuroendocrinology 2022, 112, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Al-Shawi, R.; Kinnaird, J.; Burke, J.; Bishop, J.O. Expression of a foreign gene in a line of transgenic mice is modulated by a chromosomal position effect. Mol. Cell. Biol. 1990, 10, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Bestor, T.H. Gene silencing as a threat to the success of gene therapy. J. Clin. Investig. 2000, 105, 409–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, A.R.; Harkins, R.N.; Wang, P.; Qian, H.S.; Liu, P.; Rubanyi, G.M. Transcriptional silencing is associated with extensive methylation of the CMV promoter following adenoviral gene delivery to muscle. J. Gene Med. 2004, 6, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Li, H.P.; Hung, Y.H.; Leu, Y.W.; Wu, W.H.; Wang, F.S.; Lee, K.D.; Chang, P.J.; Wu, C.S.; Lu, Y.J.; et al. Targeted methylation of CMV and E1A viral promoters. Biochem. Biophys. Res. Commun. 2010, 402, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; O’Callaghan, P.M.; Droms, K.A.; James, D.C. A mechanistic understanding of production instability in CHO cell lines expressing recombinant monoclonal antibodies. Biotechnol. Bioeng. 2011, 108, 2434–2446. [Google Scholar] [CrossRef]
- Osterlehner, A.; Simmeth, S.; Göpfert, U. Promoter methylation and transgene copy numbers predict unstable protein production in recombinant chinese hamster ovary cell lines. Biotechnol. Bioeng. 2011, 108, 2670–2681. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mariati; Chusainow, J.; Yap, M.G.S. DNA methylation contributes to loss in productivity of monoclonal antibody-producing CHO cell lines. J. Biotechnol. 2010, 147, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Aprahamian, T.; Kikuchi, R.; Shimizu, A.; Papanicolaou, K.N.; MacLauchlan, S.; Maruyama, S.; Walsh, K. Vascular rarefaction mediates whitening of brown fat in obesity. J. Clin. Investig. 2014, 124, 2099–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissing, L.; Scherer, T.; Tödter, K.; Knippschild, U.; Greve, J.W.; Buurman, W.A.; Pinnschmidt, H.O.; Rensen, S.S.; Wolf, A.M.; Bartelt, A.; et al. De novo lipogenesis in human fat and liver is linked to ChREBP-β and metabolic health. Nat. Commun. 2013, 4, 1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Ko, S.K.; Chung, S.H. Euonymus alatus prevents the hyperglycemia and hyperlipidemia induced by high-fat diet in ICR mice. J. Ethnopharmacol. 2005, 102, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Ko, S.K.; Choi, J.G.; Chung, S.H. Salicornia herbacea prevents high fat diet-induced hyperglycemia and hyperlipidemia in ICR mice. Arch. Pharm. Res. 2006, 29, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Zhuhua, Z.; Zhiquan, W.; Zhen, Y.; Yixin, N.; WeiWei, Z.; XiaoYong, L.; Yueming, L.; Hongmei, Z.; Li, Q.; Qing, S. A novel mice model of metabolic syndrome: The high-fat-high-fructose diet-fed ICR mice. Exp. Anim. 2015, 64, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahara, K.; Bannai, M.; Maruyama, K.; Suzuki, Y.; Okame, R.; Murakami, N. Characterization of a novel genetically obese mouse model demonstrating early onset hyperphagia and hyperleptinemia. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E451–E463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, R.; Sakazaki, F.; Okuno, T.; Nakamuro, K.; Ueno, H. Difference in glucose intolerance between C57BL/6J and ICR strain mice with streptozotocin/nicotinamide-induced diabetes. Biomed. Res. 2012, 33, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Shikano, K.; Iwakoshi-Ukena, E.; Saito, T.; Narimatsu, Y.; Kadota, A.; Furumitsu, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Neurosecretory protein GL induces fat accumulation in mice. J. Endocrinol. 2020, 244, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stanley, B.G.; Leibowitz, S.F. Neuropeptide Y injected in the paraventricular hypothalamus: A powerful stimulant of feeding behavior. Proc. Natl. Acad. Sci. USA 1985, 82, 3940–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, C.; Clegg, K.E.; Palmiter, R.D. Sensitivity to leptin and susceptibility to seizures of mice lacking neuropeptide Y. Nature 1996, 381, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Thorsell, A.; Heilig, M. Diverse functions of neuropeptide Y revealed using genetically modified animals. Neuropeptides 2002, 36, 182–193. [Google Scholar] [CrossRef]
- Qian, S.; Chen, H.; Weingarth, D.; Trumbauer, M.E.; Novi, D.E.; Guan, X.; Yu, H.; Shen, Z.; Feng, Y.; Frazier, E.; et al. Neither agouti-related protein nor neuropeptide Y is critically required for the regulation of energy homeostasis in mice. Mol. Cell. Biol. 2002, 22, 5027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krashes, M.J.; Koda, S.; Ye, C.; Rogan, S.C.; Adams, A.C.; Cusher, D.S.; Maratos-Flier, E.; Roth, B.L.; Lowell, B.B. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Investig. 2011, 121, 1424–1428. [Google Scholar] [CrossRef] [Green Version]
- Zhan, C.; Zhou, J.; Feng, Q.; Zhang, J.; Lin, S.; Bao, J.; Wu, P.; Luo, M. Acute and long-term suppression of feeding behavior by POMC neurons in the brainstem and hypothalamus, respectively. J. Neurosci. 2013, 33, 3624–3632. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Sato, T.; Tateyama, M.; Kageyama, H.; Maejima, Y.; Nakata, M.; Hirako, S.; Matsuo, T.; Kyaw, S.; Shiuchi, T.; et al. Activation of AMPK-regulated CRH neurons in the PVH is sufficient and necessary to Induce dietary preference for carbohydrate over fat. Cell Rep. 2018, 22, 706–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.M.; Sunagawa, G.A.; Soya, S.; Abe, M.; Sakurai, K.; Ishikawa, K.; Yanagisawa, M.; Hama, H.; Hasegawa, E.; Miyawaki, A.; et al. A discrete neuronal circuit induces a hibernation-like state in rodents. Nature 2020, 583, 109–114. [Google Scholar] [CrossRef]
- Iizuka, K.; Takeda, J.; Horikawa, Y. Glucose induces FGF21 mRNA expression through ChREBP activation in rat hepatocytes. FEBS Lett. 2009, 583, 2882–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine regulation of the fasting response by PPARα-mediated induction of fibroblast growth factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, C.; Xiao, J.; Liu, L.; Zhang, M.; Wang, C.; Wu, G.; Zheng, M.H.; Xu, L.M.; Chen, Y.P.; et al. Fibroblast growth factor 21 deficiency exacerbates chronic alcohol-induced hepatic steatosis and injury. Sci. Rep. 2016, 6, 31026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komiya, C.; Tsuchiya, K.; Shiba, K.; Miyachi, Y.; Furuke, S.; Shimazu, N.; Yamaguchi, S.; Kanno, K.; Ogawa, Y. Ipragliflozin improves hepatic steatosis in obese mice and liver dysfunction in type 2 diabetic patients irrespective of body weight reduction. PLoS ONE 2016, 11, e0151511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, R.J.F.; Yeo, G.S.H. The genetics of obesity: From discovery to biology. Nat. Rev. Genet. 2021, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | Tg | |
|---|---|---|
| Leptin (ng/mL) | 0.676 ± 0.134 | 1.96 ± 0.438 * |
| Insulin (ng/mL) | 1.06 ± 0.270 | 0.973 ± 0.149 |
| Cholesterol (mg/dL) | 123 ± 7.19 | 113 ± 4.64 |
| Triglycerides (mg/dL) | 162 ± 12.27 | 139 ± 16.0 |
| Free fatty acids (mEq/L) | 1.70 ± 0.256 | 1.84 ± 0.182 |
| Glucose (mg/dL) | 129 ± 6.11 | 136 ± 7.88 |
| Gene | Sense Primer (5′ to 3′) | Antisense Primer (5′ to 3′) |
|---|---|---|
| Gfp | ACCACTACCTGAGCACCCAGTC | GTCCATGCCGAGAGTGATCC |
| Gja5 | ACCATGGAGGTGGCCTTCA | CATGCAGGGTATCCAGGAAGA |
| Npgl | GGAACCATGGCTTAGGAAGG | TCTAAGGAGCTGAGAATATGCA |
| Acc | TCCGCACTGACTGTAACCACAT | TGCTCCGCACAGATTCTTCA |
| Fas | AGGGGTCGACCTGGTCCTCA | GCCATGCCCAGAGGGTGGTT |
| Scd1 | CTGTACGGGATCATACTGGTTC | GCCGTGCCTTGTAAGTTCTG |
| Chrebpα | CGACACTCACCCACCTCTTC | TTGTTCAGCCGGATCTTGTC |
| Chrebpβ | TCTGCAGATCGCGTGGAG | CTTGTCCCGGCATAGCAAC |
| Cpt1a | CCTGGGCATGATTGCAAAG | GGACGCCACTCACGATGTT |
| Atgl | AACACCAGCATCCAGTTCAA | GGTTCAGTAGGCCATTCCTC |
| Hsl | GCTGGGCTGTCAAGCACTGT | GTAACTGGGTAGGCTGCCAT |
| Gapdh | AAGGTCATCCCAGAGCTGAA | CTGCTTCACCACCTTCTTGA |
| Slc2a4 | GTAACTTCATTGTCGGCATGG | AGCTGAGATCTGGTCAAACG |
| Cd36 | TCCTCTGACATTTGCAGGTCTATC | AAAGGCATTGGCTGGAAGAA |
| Pparγ | GCCCTTTGGTGACTTTATGGA | GCAGCAGGTTGTCTTGGATG |
| Fgf21 | CCTCTAGGTTTCTTTGCCAACAG | AAGCTGCAGGCCTCAGGAT |
| Slc2a2 | GGCTAATTTCAGGACTGGTT | TTTCTTTGCCCTGACTTCCT |
| Pparα | TCGAATATGTGGGGACAAGG | GACAGGCACTTGTGAAAACG |
| Npy | TATCTCTGCTCGTGTGTTTG | GATTGATGTAGTGTCGCAGA |
| Agrp | TGTTCCCAGAGTTCCCAGGTC | GCATTGAAGAAGCGGCAGTAGCAC |
| Pomc | AGCTGCCTTTCCGCGACA | ATCTATGGAGGTCTGAAGCA |
| Ghrh | TGCCATCTTCACCACCAAC | TCATCTGCTTGTCCTCTGTCC |
| Sst | GAGGACCTGCGACTAGACTGAC | CAGCAGCTCTGCCAAGAAGTA |
| Trh | TCGTGCTAACTGGTATCCCC | CCCAAATCTCCCCTCTCTTC |
| Tsh | CACCATCTGTGCTGGGTATTG | CATCCTGGTATTTCCACCGTTC |
| Lh | TGGCCGCAGAGAATGAGTTC | ACTCGGACCATGCTAGGACA |
| Fsh | GGAGAGCAATCTGCTGCCAT | GCCGAGCTGGGTCCTTATAC |
| Prl | GGCTACACCTGAAGACAAGGAACAA | TGTTCCTCAATCTCTTTGGCTCTTG |
| Gh | GGAGGCTAGTGCTTTTCCCG | AGGCACGCTCGAACTCTTTG |
| Ucp1 | CAAAAACAGAAGGATTGCCGAAA | TCTTGGACTGAGTCGTAGAGG |
| Dio2 | CCACCTTCTTGACTTTGCCA | GGTGAGCCTCATCAATGTATAC |
| Pgc1α | GCAACATGCTCAAGCCAAAC | TGCAGTTCCAGAGAGTTCCA |
| Actb | GGCACCACACCTTATACAAT | AGGTCTCAAACATGATCTGG |
| Rps18 | CCTGAGAAGTTCCAGCACAT | TTCTCCAGCCCTCTTGGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narimatsu, Y.; Matsuura, D.; Iwakoshi-Ukena, E.; Furumitsu, M.; Ukena, K. Neurosecretory Protein GL Promotes Normotopic Fat Accumulation in Male ICR Mice. Int. J. Mol. Sci. 2022, 23, 6488. https://doi.org/10.3390/ijms23126488
Narimatsu Y, Matsuura D, Iwakoshi-Ukena E, Furumitsu M, Ukena K. Neurosecretory Protein GL Promotes Normotopic Fat Accumulation in Male ICR Mice. International Journal of Molecular Sciences. 2022; 23(12):6488. https://doi.org/10.3390/ijms23126488
Chicago/Turabian StyleNarimatsu, Yuki, Daichi Matsuura, Eiko Iwakoshi-Ukena, Megumi Furumitsu, and Kazuyoshi Ukena. 2022. "Neurosecretory Protein GL Promotes Normotopic Fat Accumulation in Male ICR Mice" International Journal of Molecular Sciences 23, no. 12: 6488. https://doi.org/10.3390/ijms23126488