
Facing the Green Threat: A Water Flea’s Defenses against a Carnivorous Plant

 , , , and
, , , and 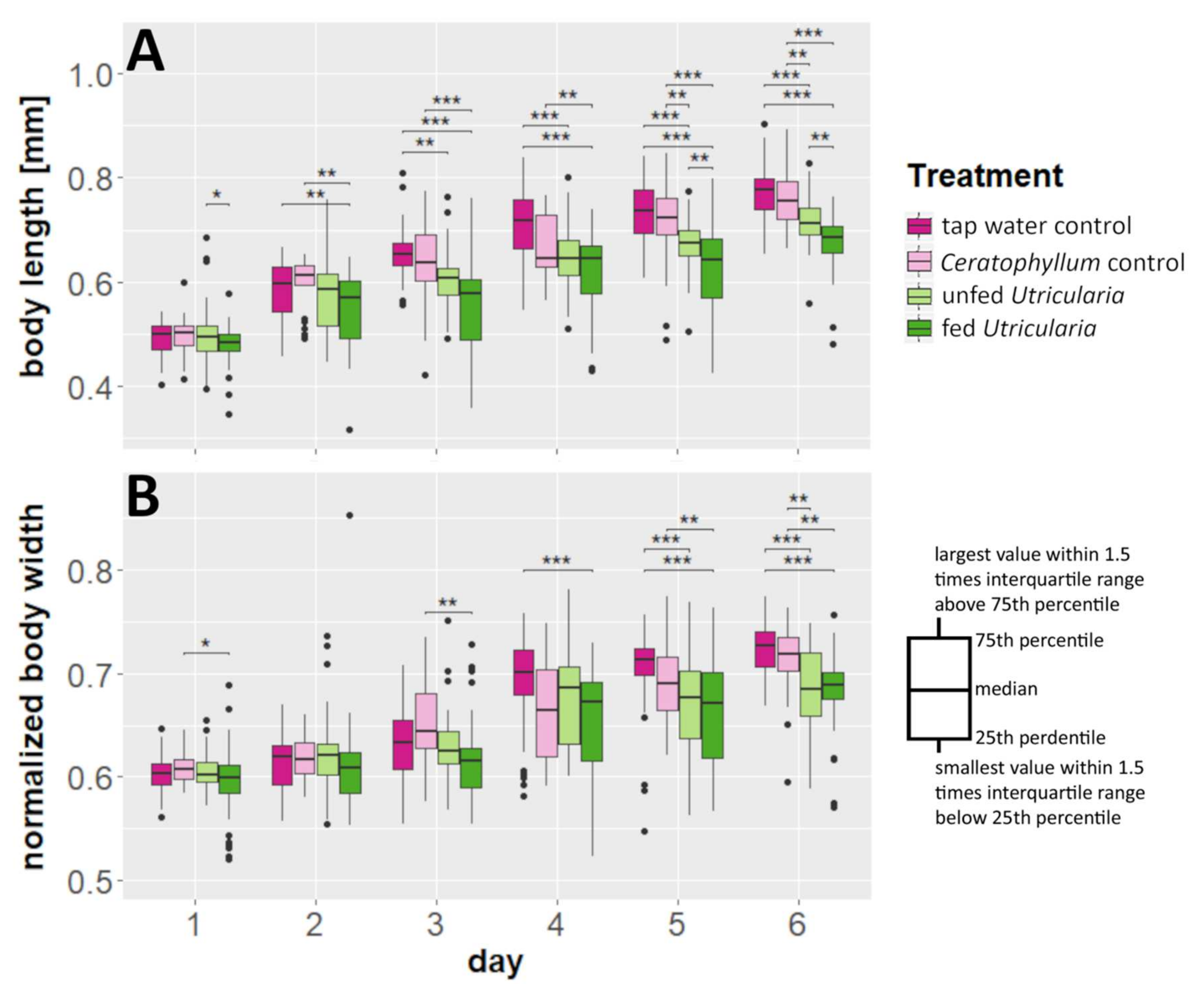{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Trap Entrance Dimensions
2.2. 2D Investigation
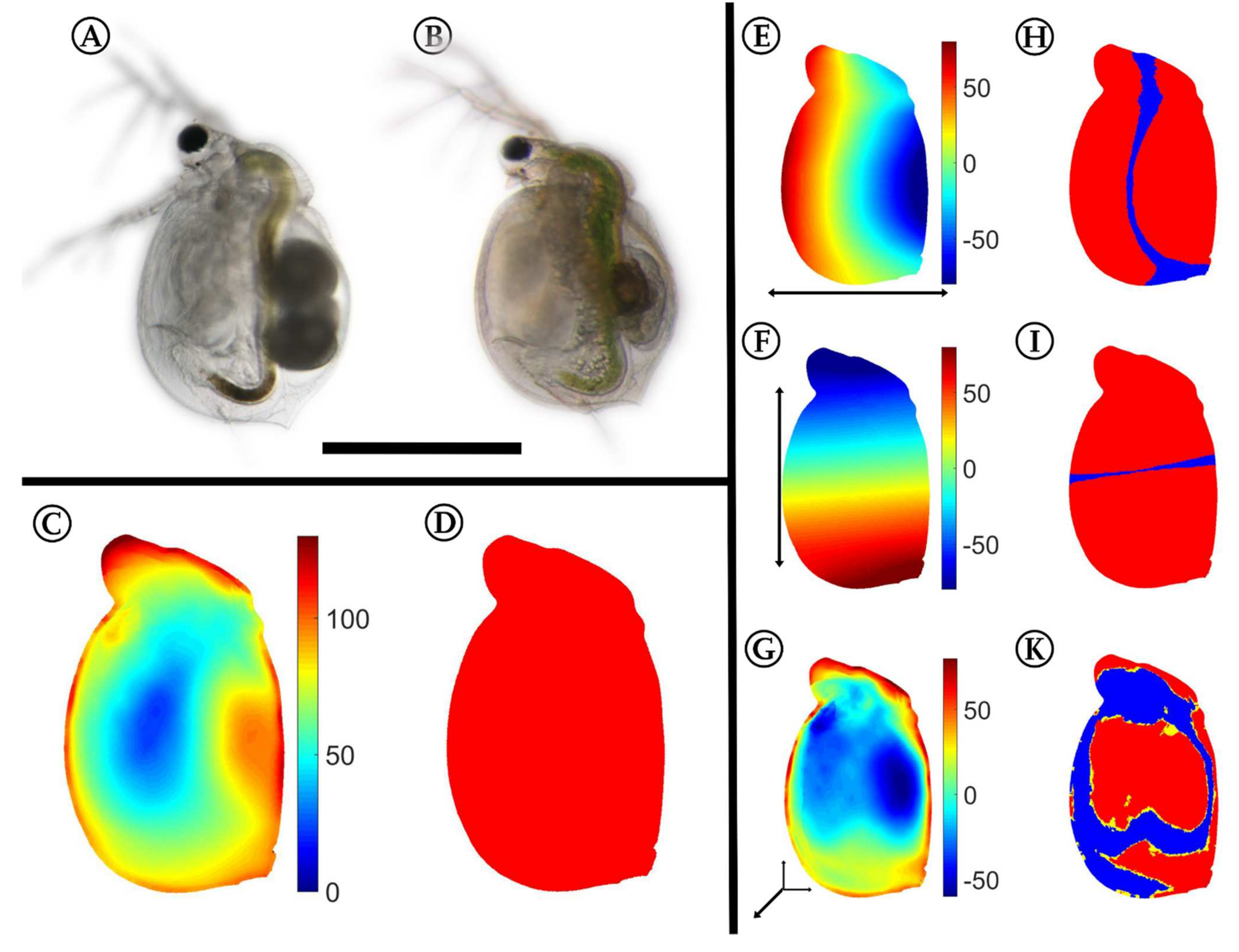
2.3. 3D Analysis
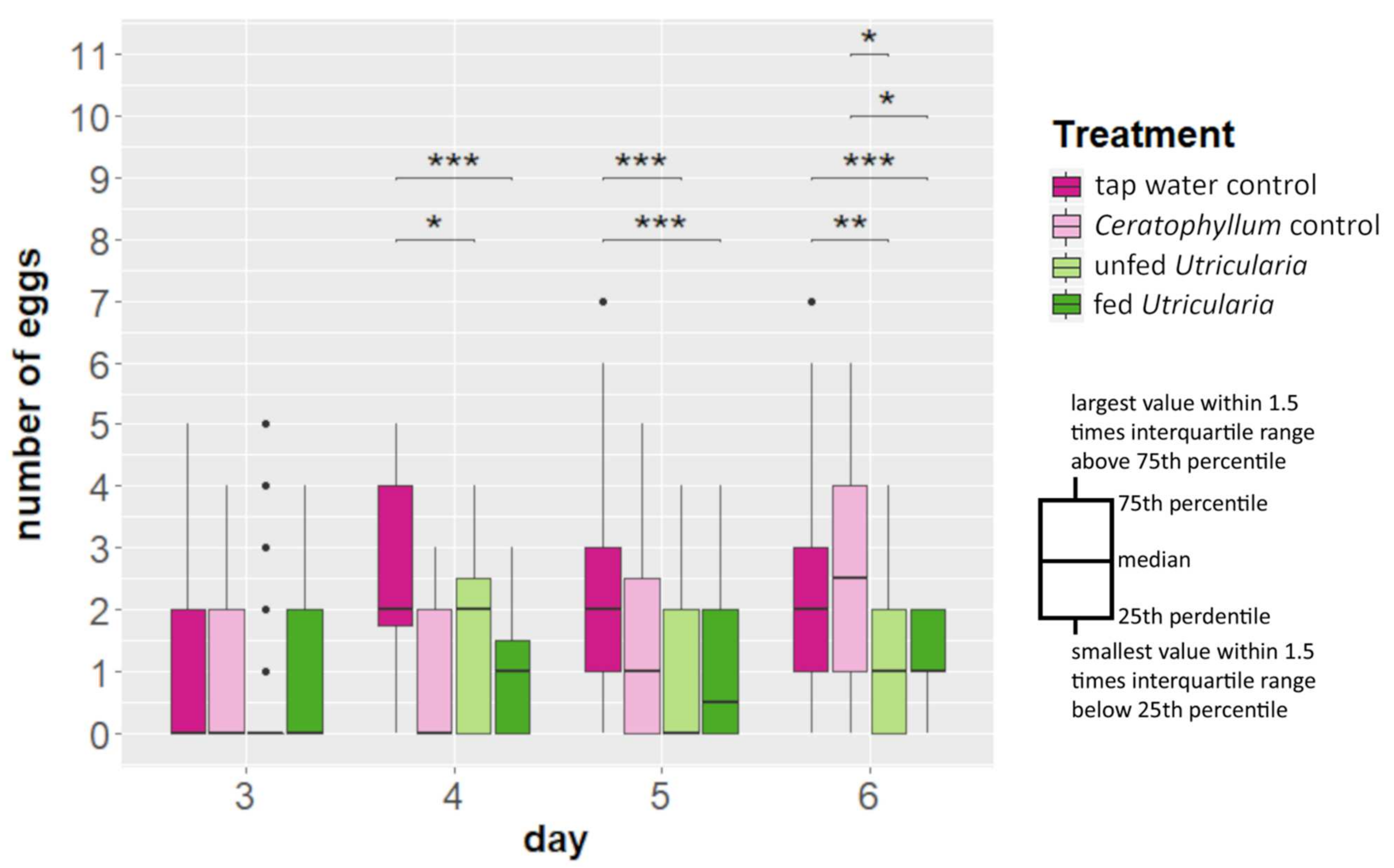
2.4. Life-History Shifts
2.5. Behavioral Alterations
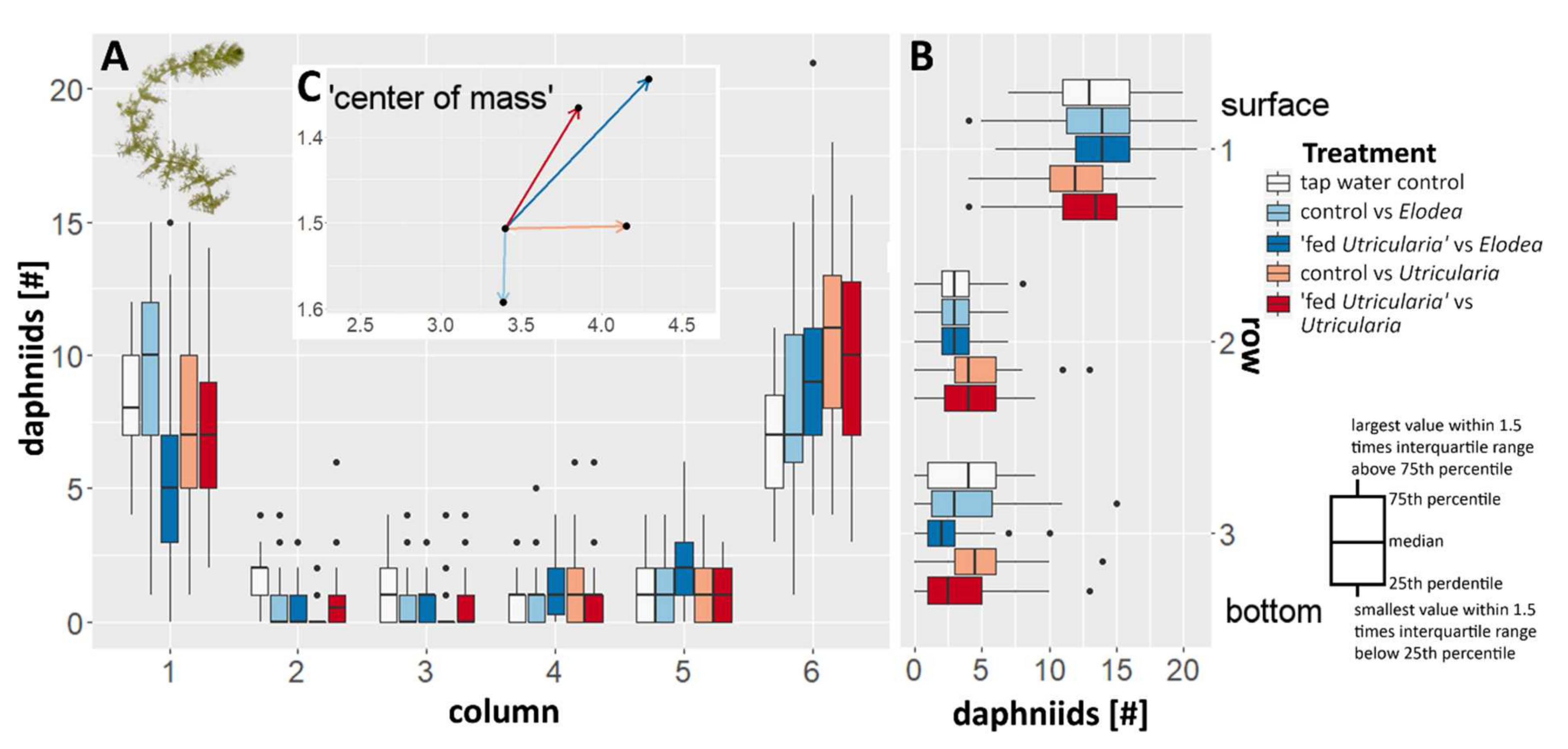
2.5.1. Predator Avoidance
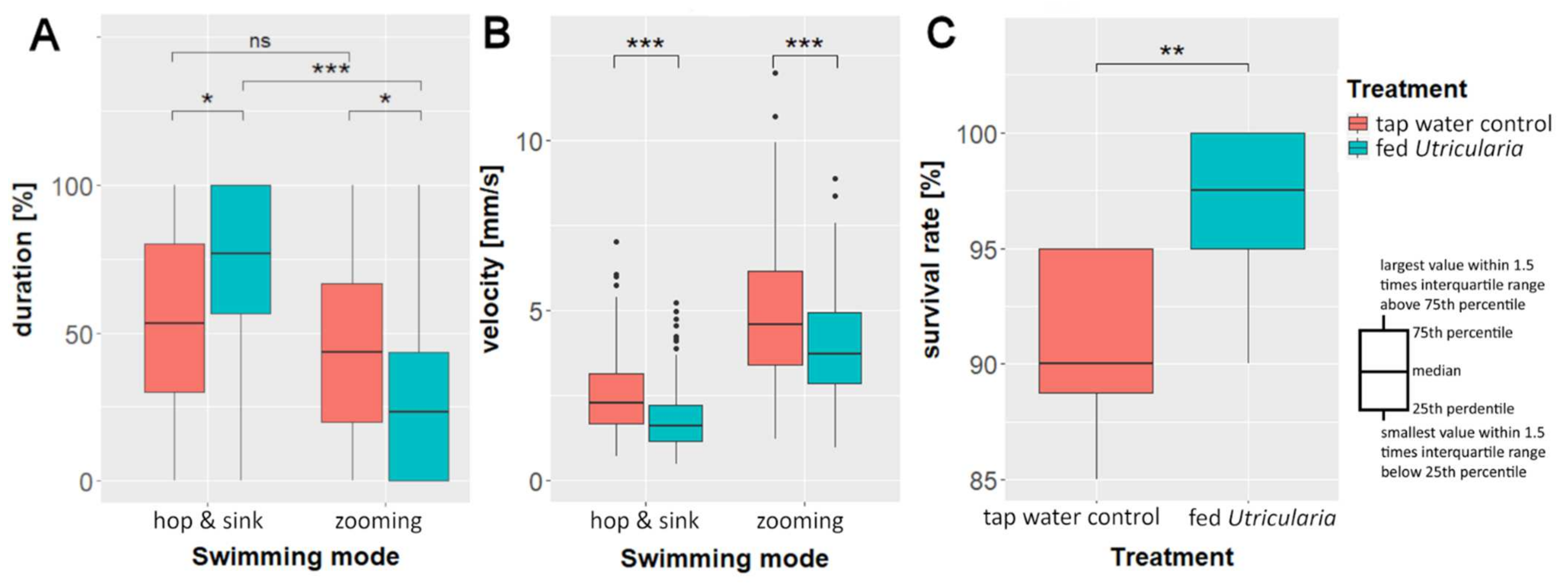
2.5.2. Swimming Modes
2.5.3. Swimming Velocity
2.6. Predation Experiments
3. Discussion
3.1. Morphological Adaptations
3.2. Life-History Adaptations
3.3. Behavioral Adaptations
3.4. Predation Trials
3.5. Origin of the Defense-Inducing Stimulus
4. Materials and Methods
4.1. Study Design
4.2. Cultures
4.2.1. Prey Crustaceans (Ceriodaphnia Dubia)
4.2.2. Predator (Carnivorous Plant Utricularia x Neglecta)
4.3. Trap Entrance Dimensions
4.4. Defense Induction
4.5. Analysis of Morphology and Life-History Alterations
4.6. 3D Analysis of Morphological Alterations
4.7. Analysis of Behavioral Defenses
4.7.1. Predator Avoidance
4.7.2. Swimming Velocity
4.7.3. Swimming Mode
4.8. Predation Experiments
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Černý, M.; Bytel, J. Density and Size Distribution of Daphnia Populations at Different Fish Predation Levels. Hydrobiologia 1991, 225, 199–208. [Google Scholar] [CrossRef]
- Zaret, T.M. Predators, Invisible Prey, and the Nature of Polymorphism in the Cladocera. Limnol. Oceanogr. 1972, 17, 171–184. [Google Scholar] [CrossRef]
- Dodson, S.I. Adaptive Change in Plankton Morphology in Response to Size-Selective Predation: A New Hypothesis of Cyclomorphosis. Limnol. Oceanogr. 1974, 19, 721–729. [Google Scholar] [CrossRef]
- Tollrian, R.; Harvell, C.D. The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar]
- Weiss, L.C.; Laforsch, C.; Tollrian, R. The Taste of Predation and the Defences of Prey. In Chemical Ecology in Aquatic Systems; Hansson, L.A., Brönmark, C., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 111–126. [Google Scholar]
- Weiss, L.C.; Tollrian, R. Predator Induced Defenses in Crustacea. In The Natural History of Crustacean; Thiel, M., Wellborn, G., Eds.; Oxford University Press: New York, NY, USA, 2018; Volume 5, pp. 303–321. [Google Scholar]
- Riessen, H.P.; Gilbert, J.J. Divergent Developmental Patterns of Induced Morphological Defenses in Rotifers and Daphnia: Ecological and Evolutionary Context. Limnol. Oceanogr. 2019, 64, 541–557. [Google Scholar] [CrossRef]
- Krueger, D.A.; Dodson, S.I. Embryological Induction and Predation Ecology in Daphnia pulex. Limnol. Oceanogr. 1981, 26, 219–223. [Google Scholar] [CrossRef]
- Spitze, K.; Sadler, T.D. Evolution of a Generalist Genotype: Multivariate Analysis of the Adaptiveness of Phenotypic Plasticity. Am. Nat. 1996, 148, 108–123. [Google Scholar] [CrossRef]
- Weiss, L.C.; Heilgenberg, E.; Deussen, L.; Becker, S.M.; Kruppert, S.; Tollrian, R. Onset of Kairomone Sensitivity and the Development of Inducible Morphological Defenses in Daphnia pulex. Hydrobiologia 2016, 779, 135–145. [Google Scholar] [CrossRef]
- Petrusek, A.; Tollrian, R.; Schwenk, K.; Haas, A.; Laforsch, C. A “Crown of Thorns” Is an Inducible Defense That Protects Daphnia against an Ancient Predator. Proc. Natl. Acad. Sci. USA 2009, 106, 2248–2252. [Google Scholar] [CrossRef]
- Tollrian, R. Predator-Induced Helmet Formation in Daphnia cucullata (Sars). Arch. für Hydrobiol. 1990, 119, 191–196. [Google Scholar] [CrossRef]
- Tollrian, R. Fish-Kairomone Induced Morphological Changes in Daphnia lumholtzi (Sars). Arch. für Hydrobiol. 1994, 130, 69–75. [Google Scholar] [CrossRef]
- Barry, M.J.; Bayly, I.A. Further Studies on Predator Induction of Crests in Australian Daphnia and the Effects of Crests on Predation. Mar. Freshw. Res. 1985, 36, 519–535. [Google Scholar] [CrossRef]
- Weiss, L.C.; Leimann, J.; Tollrian, R. Predator-Induced Defences in Daphnia longicephala: Location of Kairomone Receptors and Timeline of Sensitive Phases to Trait Formation. J. Exp. Biol. 2015, 218, 2918–2926. [Google Scholar] [CrossRef]
- Grant, J.W.G.; Bayly, I.A. Predator Induction of Crests in Morphs of the Daphnia carinata King Complex. Limnol. Oceanogr. 1981, 26, 201–218. [Google Scholar] [CrossRef]
- Laforsch, C.; Ngwa, W.; Grill, W.; Tollrian, R. An Acoustic Microscopy Technique Reveals Hidden Morphological Defenses in Daphnia. Proc. Natl. Acad. Sci. USA 2004, 101, 15911–15914. [Google Scholar] [CrossRef]
- Rabus, M.; Söllradl, T.; Clausen-Schaumann, H.; Laforsch, C. Uncovering Ultrastructural Defences in Daphnia magna—An Interdisciplinary Approach to Assess the Predator-Induced Fortification of the Carapace. PLoS ONE 2013, 8, e67856. [Google Scholar] [CrossRef]
- Kruppert, S.; Horstmann, M.; Weiss, L.C.; Witzel, U.; Schaber, C.F.; Gorb, S.N.; Tollrian, R. Biomechanical Properties of Predator-Induced Body Armour in the Freshwater Crustacean Daphnia. Sci. Rep. 2017, 7, 9750. [Google Scholar] [CrossRef]
- Machacek, J. Indirect Effect of Planktivorous Fish and the Growth and Reproduction of Daphnia galeata. Hydrobiologia 1991, 225, 193–197. [Google Scholar] [CrossRef]
- Weider, L.J.; Pijanowska, J. Plasticity of Daphnia Life Histories in Response to Chemical Cues from Predators. Oikos 1993, 67, 385–392. [Google Scholar] [CrossRef]
- Dodson, S.I.; Havel, J.E. Indirect Prey Effects: Some Morphological and Life History Responses of Daphnia pulex Exposed to Notonecta undulata. Limnol. Oceanogr. 1988, 33, 1274–1285. [Google Scholar] [CrossRef]
- Hanazato, T.; Yasuno, M. Zooplankton Community Structure Driven by Vertebrate and Invertebrate Predators. Oecologia 1989, 81, 450–458. [Google Scholar] [CrossRef]
- Stibor, H. Predator Induced Life-History Shifts in a Freshwater Cladoceran. Oecologia 1992, 92, 162–165. [Google Scholar] [CrossRef]
- Stibor, H.; Lüning, J. Predator-Induced Phenotypic Variation in the Pattern of Growth and Reproduction in Daphnia hyalina (Crustacea: Cladocera). Funct. Ecol. 1994, 8, 97–101. [Google Scholar] [CrossRef]
- Lampert, W. The Adaptive Significance of Diel Vertical Migration of Zooplankton. Funct. Ecol. 1989, 3, 21–27. [Google Scholar] [CrossRef]
- Langer, S.M.; Tollrian, R.; Bianco, G.; Hansson, L.-A.; Weiss, L.C.; Ekvall, M.T. A Three-Dimensional Perspective of Daphnia ’s Swimming Behavior with and without Predator Cues. Limnol. Oceanogr. 2019, 64, 1515–1525. [Google Scholar] [CrossRef]
- Havel, J.E.; Dodson, S.I. Environmental Cues for Cyclomorphosis in Daphnia retrocurva Forbes. Freshw. Biol. 1985, 15, 469–478. [Google Scholar] [CrossRef]
- Mette, N.; Wilbert, N.; Barthlott, W. Food Composition of Aquatic Bladderworts (Utricularia, Lentibulariaceae) in Various Habiats. Beiträge zur Biol. der Pflanz. 2000, 72, 1–14. [Google Scholar]
- Kurbatova, S.A.; Yershov, I.Y. Crustaceans and Rotifers in the Predatory Feeding of Utricularia. Inland Water Biol. 2009, 2, 271–275. [Google Scholar] [CrossRef]
- Harms, S. The Effect of Bladderwort (Utricularia) Predation on Microcrustacean Prey. Freshw. Biol. 2002, 47, 1608–1617. [Google Scholar] [CrossRef]
- Poppinga, S.; Daber, L.E.; Westermeier, A.S.; Kruppert, S.; Horstmann, M.; Tollrian, R.; Speck, T. Biomechanical Analysis of Prey Capture in the Carnivorous Southern Bladderwort (Utricularia australis). Sci. Rep. 2017, 7, 1776. [Google Scholar] [CrossRef]
- Adamec, L. Functional Characteristics of Traps of Aquatic Carnivorous Utricularia Species. Aquat. Bot. 2011, 95, 226–233. [Google Scholar] [CrossRef]
- Sasago, A.; Sibaoka, T. Water Extrusion in the Trap Bladders of Utricularia vulgaris II. A Possible Mechanism of Water Outflow. Bot. Mag. Tokyo 1985, 98, 113–124. [Google Scholar] [CrossRef]
- Sasago, A.; Sibaoka, T. Water Extrusion in the Trap Bladders of Utricularia vulgaris I. A Possible Pathway of Water across the Bladder Wall. Bot. Mag. Tokyo 1985, 98, 55–66. [Google Scholar] [CrossRef]
- Vincent, O.; Weisskopf, C.; Poppinga, S.; Masselter, T.; Speck, T.; Joyeux, M.; Quilliet, C.; Marmottant, P. Ultra-Fast Underwater Suction Traps. Proc. Biol. Sci. 2011, 278, 2909–2914. [Google Scholar] [CrossRef] [PubMed]
- Poppinga, S.; Weisskopf, C.; Westermeier, A.S.; Masselter, T.; Speck, T. Fastest Predators in Plant Kingdom: Functional Morphology and Biomechanics of Suction Traps Found in the Largest Genus of Carnivorous Plants. AoB Plants 2016, 8, plv140. [Google Scholar] [CrossRef]
- Juniper, B.E.; Robins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989; ISBN 0-12-392170-8. [Google Scholar]
- Gordon, E.; Pacheco, S. Prey Composition in the Carnivorous Plants Utricularia inflata and U. gibba (Lentibulariaceae) from Paria Peninsula, Venezuela. Rev. Biol. Trop. 2007, 55, 795–803. [Google Scholar] [CrossRef]
- Horstmann, M.; Fleischmann, A.; Tollrian, R.; Poppinga, S. Snapshot Prey Spectrum Analysis of the Phylogenetically Early-Diverging Carnivorous Utricularia multifida from U. Section Polypompholyx (Lentibulariaceae). PLoS ONE 2021, 16, e0249976. [Google Scholar] [CrossRef]
- Horstmann, M.; Topham, A.T.; Stamm, P.; Kruppert, S.; Colbourne, J.K.; Tollrian, R.; Weiss, L.C. Scan, Extract, Wrap, Compute—a 3D Method to Analyse Morphological Shape Differences. PeerJ 2018, 6, e4861. [Google Scholar] [CrossRef]
- Chen, M.S. Inducible Direct Plant Defense against Insect Herbivores: A Review. Insect Sci. 2008, 15, 101–114. [Google Scholar] [CrossRef]
- Keathley, C.P.; Potter, D.A. Behavioral Plasticity of a Grass-Feeding Caterpillar in Response to Spiny- or Smooth-Edged Leaf Blades. Arthropod. Plant. Interact. 2011, 5, 339–349. [Google Scholar] [CrossRef]
- Thompson, D.B. Consumption Rates and the Evolution of Diet-Induced Plasticity in the Head Morphology of Melanoplus femurrubrum (Orthoptera: Acrididae). Oecologia 1992, 89, 204–213. [Google Scholar] [CrossRef]
- Guiral, D.; Rougier, C. Trap Size and Prey Selection of Two Coexisting Bladderwort (Utricularia) Species in a Pristine Tropical Pond (French Guiana) at Different Trophic Levels. Ann. Limnol. 2007, 43, 147–159. [Google Scholar] [CrossRef]
- Dodson, S.I. The Ecological Role of Chemical Stimuli for the Zooplankton: Predator-Induced Morphology in Daphnia. Oecologia 1989, 78, 361–367. [Google Scholar] [CrossRef]
- Tucker, R.P.; Woolpy, S.P. The Effect of Parthenogenetic Eggs in Daphnia magna on Prey Location by the Bluegill Sunfish (Lepomis macrochirus). Hydrobiologia 1984, 109, 215–217. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation, Body Size and Composition of Plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Aránguiz-Acuña, A.; Ramos-Jiliberto, R.; Sarma, N.; Sarma, S.S.S.; Bustamante, R.O.; Toledo, V. Benefits, Costs and Reactivity of Inducible Defences: An Experimental Test with Rotifers. Freshw. Biol. 2010, 55, 2114–2122. [Google Scholar] [CrossRef]
- Dodson, S.I.; Hanazato, T.; Gorski, P.R. Behavioral Responses of Daphnia pulex Exposed to Carbaryl and Chaoborus Kairomone. Environ. Toxicol. Chem. 1995, 14, 43–50. [Google Scholar] [CrossRef]
- Freeman, A.S. The Maintenance, Evolution, and Impacts of Inducible Morphological Defenses in Mytilus edulis: Responses to Multiple and Invasive Predators. Ph.D. Dissertation, University of New Hampshire, Durham, NH, USA, 2007. [Google Scholar]
- De Meester, L.; Cousyn, C. The Change in Phototactic Behaviour of a Daphnia magna Clone in the Presence of Fish Kairomones: The Effect of Exposure Time. Hydrobiologia 1997, 360, 169–175. [Google Scholar] [CrossRef]
- Lauridsen, T.L.; Lodge, D.M. Avoidance by Daphnia magna of Fish and Macrophytes: Chemical Cues and Predator-Mediated Use of Macrophyte Habitat. Limnol. Oceanogr. 1996, 41, 794–798. [Google Scholar] [CrossRef]
- Gerritsen, J.; Strickler, J.R. Encounter Probabilities and Community Structure in Zooplankton: A Mathematical Model. J. Fish. Board Canada 1977, 34, 73–82. [Google Scholar] [CrossRef]
- Pijanowska, J.; Kowalczewski, A. Cues from Injured Daphnia and from Cyclopoids Feeding on Daphnia Can Modify Life Histories of Conspecifics. Hydrobiologia 1997, 350, 99–103. [Google Scholar] [CrossRef]
- Weiss, L.C. Sensory Ecology of Predator-Induced Phenotypic Plasticity. Front. Behav. Neurosci. 2019, 12, 330. [Google Scholar] [CrossRef]
- Tollrian, R.; Dodson, S.I. Inducible Defenses in Cladocera: Constraints, Costs, and Multipredator Environments. In The Ecology and Evolution of Inducible Defenses; Tollrian, R., Harvell, C.D., Eds.; Princeton University Press: Princeton, NJ, USA, 1999; pp. 177–202. [Google Scholar]
- Laforsch, C.; Beccara, L.; Tollrian, R. Inducible Defenses: The Relevance of Chemical Alarm Cues in Daphnia. Limnol. Oceanogr. 2006, 51, 1466–1472. [Google Scholar] [CrossRef]
- Brown, W.L.; Eisner, T.; Whittaker, R.H. Allomones and Kairomones: Transspecific Chemical Messengers. Bioscience 1970, 20, 21–22. [Google Scholar] [CrossRef]
- Weiss, L.C.; Albada, B.; Becker, S.M.; Meckelmann, S.W.; Klein, J.; Meyer, M.; Schmitz, O.J.; Sommer, U.; Leo, M.; Zagermann, J.; et al. Identification of Chaoborus Kairomone Chemicals That Induce Defences in Daphnia. Nat. Chem. Biol. 2018, 14, 1133–1139. [Google Scholar] [CrossRef]
- Tollrian, R. Neckteeth Formation in Daphnia pulex as an Example of Continuous Phenotypic Plasticity: Morphological Effects of Chaoborus Kairomone Concentration and Their Quantification. J. Plankton Res. 1993, 15, 1309–1318. [Google Scholar] [CrossRef]
- Adamec, L.; Poppinga, S. Measurement of the Critical Negative Pressure inside Traps of Aquatic Carnivorous Utricularia Species. Aquat. Bot. 2016, 133, 10–16. [Google Scholar] [CrossRef]
- Adamec, L. Photosynthetic CO2 Affinity of the Aquatic Carnivorous Plant Utricularia australis (Lentibulariaceae) and Its Investment in Carnivory. Ecol. Res. 2009, 24, 327–333. [Google Scholar] [CrossRef]
- Hall, M.R. Assessing the Costs and Benefits of Active Traps in the Carnivorous Plant Utricularia Australis: Effects of Trap Size and Age; University of California: Fresno, CA, USA, 2018. [Google Scholar]
- Englund, G.; Harms, S. Effects of Light and Microcrustacean Prey on Growth and Investment in Carnivory in Utricularia vulgaris. Freshw. Biol. 2003, 48, 786–794. [Google Scholar] [CrossRef]
- Adamec, L. Respiration and Photosynthesis of Bladders and Leaves of Aquatic Utricularia Species. Plant Biol. 2006, 8, 765–769. [Google Scholar] [CrossRef]
- Bobrov, A.A.; Volkova, P.A.; Kopylov-Guskov, Y.O.; Mochalova, O.A.; Kravchuk, A.E.; Nekrasova, D.M. Unknown Sides of Utricularia (Lentibulariaceae) Diversity in East Europe and North Asia or How Hybridization Explained Old Taxonomical Puzzles. Perspect. Plant Ecol. Evol. Syst. 2022, 54, 125649. [Google Scholar] [CrossRef]
- Kruppert, S.; Horstmann, M.; Weiss, L.C.; Schaber, C.F.; Gorb, S.N.; Tollrian, R. Push or Pull? The Light-Weight Architecture of the Daphnia pulex Carapace Is Adapted to Withstand Tension, Not Compression. J. Morphol. 2016, 277, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Westermeier, A.S.; Fleischmann, A.; Müller, K.; Schäferhoff, B.; Rubach, C.; Speck, T.; Poppinga, S. Trap Diversity and Character Evolution in Carnivorous Bladderworts (Utricularia, Lentibulariaceae). Sci. Rep. 2017, 7, 12052. [Google Scholar] [CrossRef] [PubMed]
- Adamec, L. Why Do Aquatic Carnivorous Plants Prefer Growing in Dystrophic Waters? ACTA Biol. Slov. 2012, 55, 3–8. [Google Scholar]
- Mitchell, M.D.; Bairos-Novak, K.R.; Ferrari, M.C.O. Mechanisms Underlying the Control of Responses to Predator Odours in Aquatic Prey. J. Exp. Biol. 2017, 220, 1937–1946. [Google Scholar] [CrossRef] [PubMed]
- Ekvall, M.T.; Bianco, G.; Linse, S.; Linke, H.; Bäckman, J.; Hansson, L.A. Three-Dimensional Tracking of Small Aquatic Organisms Using Fluorescent Nanoparticles. PLoS ONE 2013, 8, e78498. [Google Scholar] [CrossRef]
- Wickramarathna, L.N.; Noss, C.; Lorke, A. Hydrodynamic Trails Produced by Daphnia: Size and Energetics. PLoS ONE 2014, 9, e92383. [Google Scholar] [CrossRef]
- O’Keefe, T.C.; Brewer, M.C.; Dodson, S.I. Swimming Behavior of Daphnia: Its Role in Determining Predation Risk. J. Plankton Res. 1998, 20, 973–984. [Google Scholar] [CrossRef]
- Dodson, S.I.; Ramcharan, C. Size-Specific Swimming Behavior of Daphnia pulex. J. Plankton Res. 1991, 13, 1367–1379. [Google Scholar] [CrossRef]
- Mort, M.A. Chaoborus Predation and the Function of Phenotypic Variation in Daphnia. Hydrobiologia 1986, 133, 39–44. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Gorjanc, G.; Grothendieck, G.; Korosec, A.; Lumley, T.; MacQueen, D.; Magnusson, A.; Rogers, J.; Others, A. Gdata: Various R Programming Tools for Data Manipulation. R Package, version 2.18.0. 2017. [Google Scholar]
- Kassambara, A. “Ggpubr”: “ggplot2” Based Publication Ready Plots. R Package, version 0.2.5. 2020. [Google Scholar]
- Ahlmann-Eltze, C. Ggsignif: Significance Brackets for “Ggplot2”. R Package, version 0.6.0. 2019. [Google Scholar]
- Kassambra, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests; R Package, version 0.6.0; 2020.
- Storey, J.D.; Tibshirani, R. Statistical Significance for Genomewide Studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef]
- Adamec, L.; Kučerová, A. Overwintering Temperatures Affect Freezing Temperatures of Turions of Aquatic Plants. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 497–501. [Google Scholar] [CrossRef]
- Adamec, L. Mineral Nutrient Relations in the Aquatic Carnivorous Plant Utricularia australis and Its Investment in Carnivory. Fundam. Appl. Limnol./Arch. für Hydrobiol. 2008, 171, 175–183. [Google Scholar] [CrossRef]
- Horstmann, M.; Heier, L.; Kruppert, S.; Weiss, L.C.; Tollrian, R.; Adamec, L.; Westermeier, A.S.; Speck, T.; Poppinga, S. Comparative Prey Spectra Analyses on the Endangered Aquatic Carnivorous Waterwheel Plant (Aldrovanda vesiculosa, Droseraceae) at Several Naturalized Microsites in the Czech Republic and Germany. Integr. Org. Biol. 2019, 1, oby012. [Google Scholar] [CrossRef]
- Poppinga, S.; Smaij, J.; Westermeier, A.S.; Horstmann, M.; Kruppert, S.; Tollrian, R.; Speck, T. Prey Capture Analyses in the Carnivorous Aquatic Waterwheel Plant (Aldrovanda vesiculosa L., Droseraceae). Sci. Rep. 2019, 9, 18590. [Google Scholar] [CrossRef]
- Darnowski, D.; Bauer, U.; Mendez, M.; Horner, J.D.; Plancho, B.J. Prey Selection and Specialization by Carnivorous Plants. In Carnivorous Plants: Physiology, Ecology and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 285–293. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruppert, S.; Horstmann, M.; Weiss, L.C.; Konopka, E.; Kubitza, N.; Poppinga, S.; Westermeier, A.S.; Speck, T.; Tollrian, R. Facing the Green Threat: A Water Flea’s Defenses against a Carnivorous Plant. Int. J. Mol. Sci. 2022, 23, 6474. https://doi.org/10.3390/ijms23126474
Kruppert S, Horstmann M, Weiss LC, Konopka E, Kubitza N, Poppinga S, Westermeier AS, Speck T, Tollrian R. Facing the Green Threat: A Water Flea’s Defenses against a Carnivorous Plant. International Journal of Molecular Sciences. 2022; 23(12):6474. https://doi.org/10.3390/ijms23126474
Chicago/Turabian StyleKruppert, Sebastian, Martin Horstmann, Linda C. Weiss, Elena Konopka, Nadja Kubitza, Simon Poppinga, Anna S. Westermeier, Thomas Speck, and Ralph Tollrian. 2022. "Facing the Green Threat: A Water Flea’s Defenses against a Carnivorous Plant" International Journal of Molecular Sciences 23, no. 12: 6474. https://doi.org/10.3390/ijms23126474