
Beneficial Effects of O-GlcNAc Stimulation in a Young Rat Model of Sepsis: Beyond Modulation of Gene Expression

 , , , , , , ,
, , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Impact of Acute O-GlcNAc Stimulation on Young Septic Rats
2.2. NButGT Treatment Increases Cardiac O-GlcNAc Levels without Impacting O-GlcNAc Cycling Enzymes
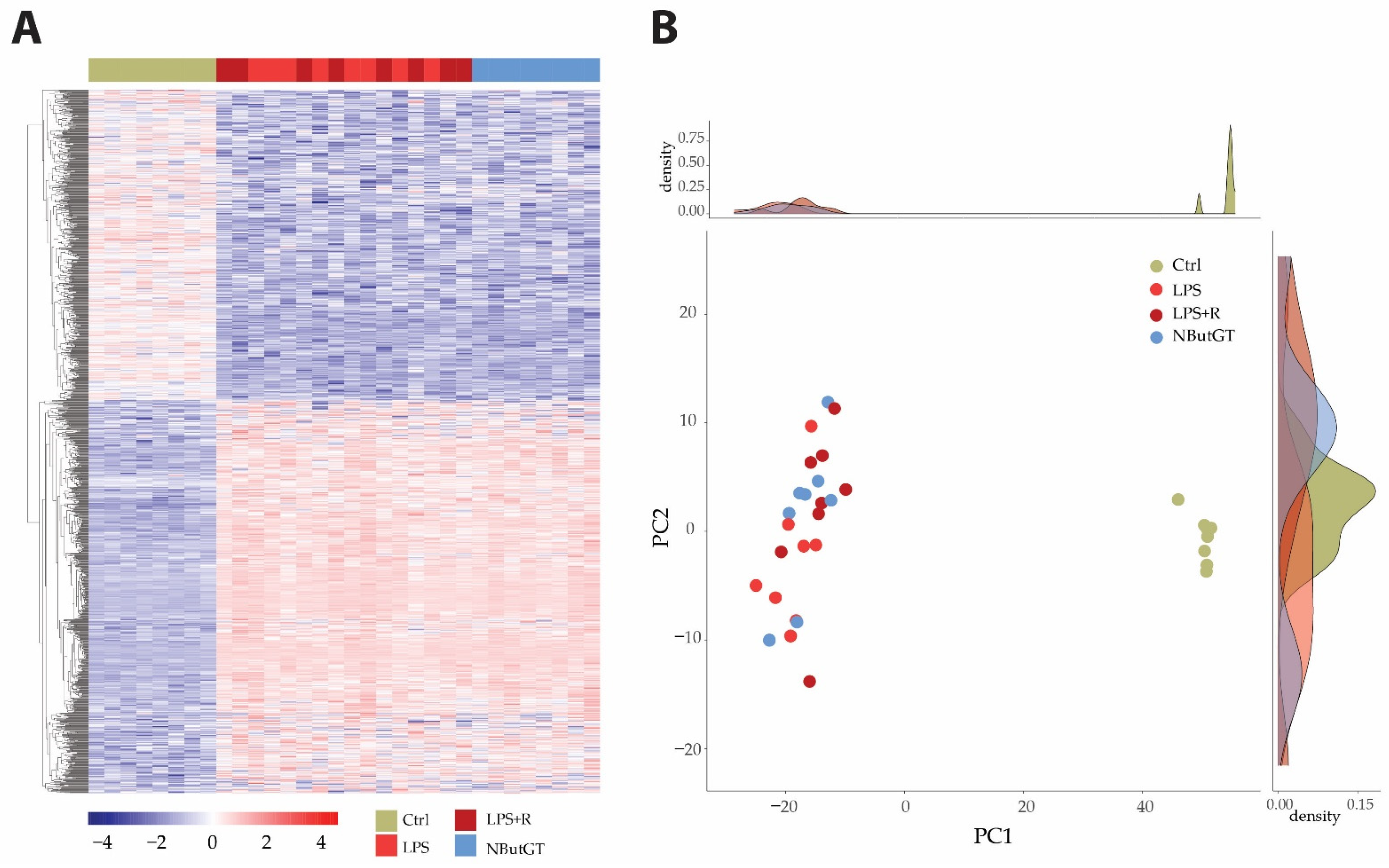
2.3. The LPS Injection Stimulates Pro-Inflammatory Pathways While O-GlcNAc Stimulation Does Not Impact Gene Expression Profiles
3. Discussion
3.1. Lipopolysaccharide Injection Promotes Pro-Inflammatory State
3.2. Gene Expressions Involved in the Integrated Stress Response and Cell Death Are Upregulated with Sepsis
3.3. Transcriptomic Pattern throughout Aging and Species
3.4. Beneficial Effects of O-GlcNAcylation Stimulation in Sepsis: New Insights
4. Methods
4.1. Reagents
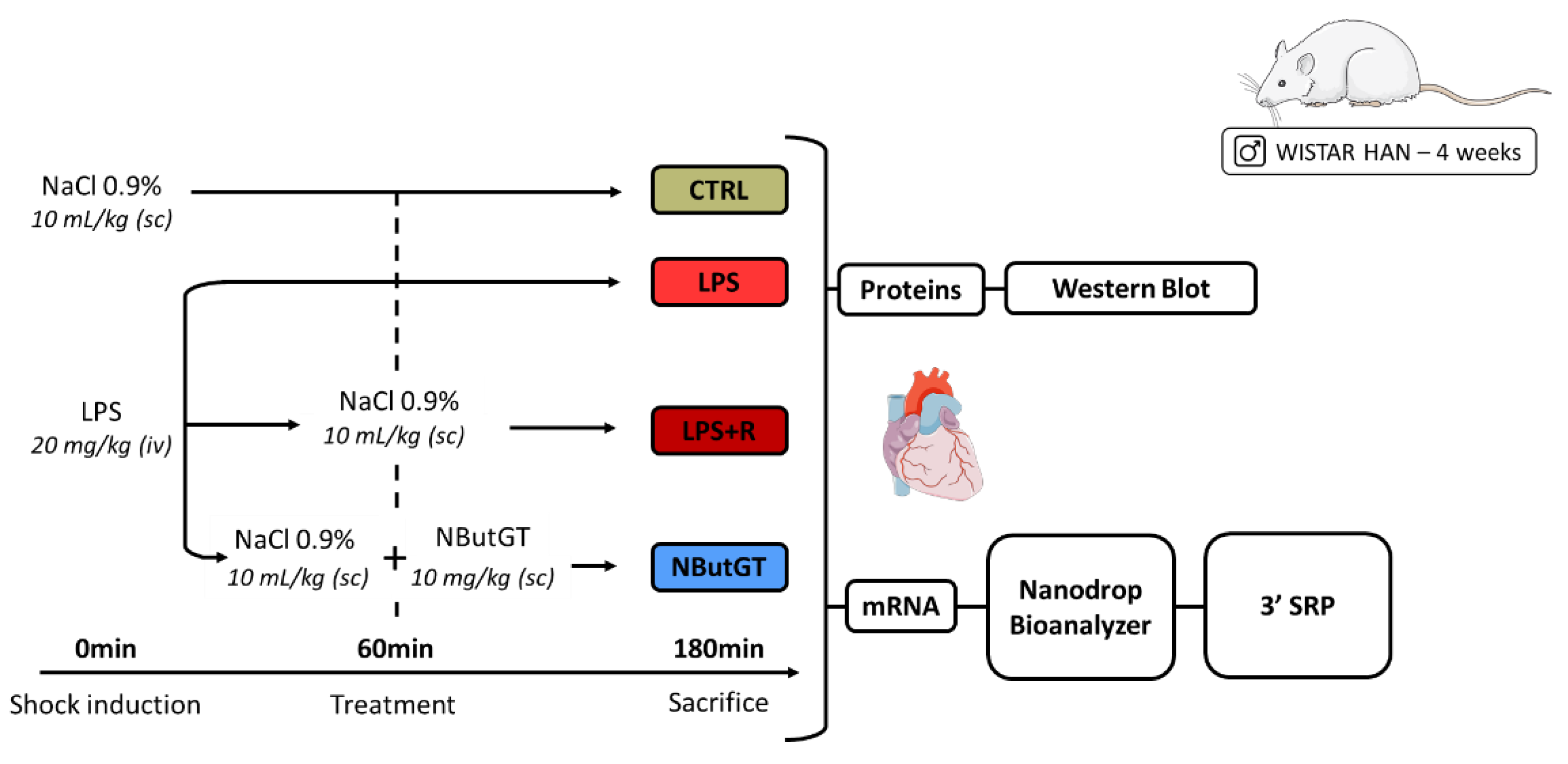
4.2. Animal Models
4.3. Heart Sampling
4.4. Tissue Preparation
4.5. Protein Extraction and Western Blots
4.6. RNA Extraction, Quality Evaluation and DGESeq Processing and Analyses
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, Regional, and National Sepsis Incidence and Mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Schnelzer, A.; Hannemann, N.; Schett, G.; Soulat, D.; Bozec, A. The Transcription Factor FRA-1/AP-1 Controls Lipocalin-2 Expression and Inflammation in Sepsis Model. Front. Immunol. 2021, 12, 701675. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.Y.; Rinchai, D.; Toufiq, M.; Kabeer, B.S.A.; Roelands, J.; Hendrickx, W.; Boughorbel, S.; Bedognetti, D.; Van Panhuys, N.; Chaussabel, D.; et al. Transcriptomic Profile Investigations Highlight a Putative Role for NUDT16 in Sepsis. J. Cell Mol. Med. 2022, 26, 1714–1721. [Google Scholar] [CrossRef]
- Santarsiero, A.; Convertini, P.; Todisco, S.; Pierri, C.L.; De Grassi, A.; Williams, N.C.; Iacobazzi, D.; De Stefano, G.; O’Neill, L.A.J.; Infantino, V. ACLY Nuclear Translocation in Human Macrophages Drives Proinflammatory Gene Expression by NF-ΚB Acetylation. Cells 2021, 10, 2962. [Google Scholar] [CrossRef]
- Lorente-Sorolla, C.; Garcia-Gomez, A.; Català-Moll, F.; Toledano, V.; Ciudad, L.; Avendaño-Ortiz, J.; Maroun-Eid, C.; Martín-Quirós, A.; Martínez-Gallo, M.; Ruiz-Sanmartín, A.; et al. Inflammatory Cytokines and Organ Dysfunction Associate with the Aberrant DNA Methylome of Monocytes in Sepsis. Genome Med. 2019, 11, 66. [Google Scholar] [CrossRef]
- Wulff-Fuentes, E.; Berendt, R.R.; Massman, L.; Danner, L.; Malard, F.; Vora, J.; Kahsay, R.; Olivier-Van Stichelen, S. The Human O-GlcNAcome Database and Meta-Analysis. Sci. Data 2021, 8, 25. [Google Scholar] [CrossRef]
- Bond, M.R.; Hanover, J.A. A Little Sugar Goes a Long Way: The Cell Biology of O-GlcNAc. J. Cell Biol. 2015, 208, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Hardivillé, S.; Hart, G.W. Nutrient Regulation of Signaling, Transcription, and Cell Physiology by O-GlcNAcylation. Cell Metab. 2014, 20, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Kelly, W.G.; Dahmus, M.E.; Hart, G.W. RNA Polymerase II Is a Glycoprotein. Modification of the COOH-Terminal Domain by O-GlcNAc. J. Biol. Chem. 1993, 268, 10416–10424. [Google Scholar] [CrossRef]
- Yang, Y.R.; Suh, P.-G. O-GlcNAcylation in Cellular Functions and Human Diseases. Adv. Biol. Regul. 2014, 54, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Zachara, N.; Akimoto, Y.; Hart, G.W. The O-GlcNAc Modification. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Ferron, M.; Cadiet, J.; Persello, A.; Prat, V.; Denis, M.; Erraud, A.; Aillerie, V.; Mevel, M.; Bigot, E.; Chatham, J.C.; et al. O-GlcNAc Stimulation: A New Metabolic Approach to Treat Septic Shock. Sci. Rep. 2019, 9, 18751. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.; Dupas, T.; Persello, A.; Dontaine, J.; Bultot, L.; Betus, C.; Pelé, T.; Dhot, J.; Erraud, A.; Maillard, A.; et al. An O-GlcNAcylomic Approach Reveals ACLY as a Potential Target in Sepsis in the Young Rat. Int. J. Mol. Sci. 2021, 22, 9236. [Google Scholar] [CrossRef]
- Macauley, M.S.; He, Y.; Gloster, T.M.; Stubbs, K.A.; Davies, G.J.; Vocadlo, D.J. Inhibition of O-GlcNAcase Using a Potent and Cell-Permeable Inhibitor Does Not Induce Insulin Resistance in 3T3-L1 Adipocytes. Chem. Biol. 2010, 17, 937–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angus, D.C.; van der Poll, T. Severe Sepsis and Septic Shock. N. Engl. J. Med. 2013, 369, 840–851. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Leopold, S.J.; Cranendonk, D.R.; van der Poll, T. Host Innate Immune Responses to Sepsis. Virulence 2014, 5, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutschman, C.S.; Tracey, K.J. Sepsis: Current Dogma and New Perspectives. Immunity 2014, 40, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Singer, M.; De Santis, V.; Vitale, D.; Jeffcoate, W. Multiorgan Failure Is an Adaptive, Endocrine-Mediated, Metabolic Response to Overwhelming Systemic Inflammation. Lancet 2004, 364, 545–548. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Liu, S.F.; Malik, A.B. NF-ΚB Activation as a Pathological Mechanism of Septic Shock and Inflammation. Am. J. Physiol. -Lung Cell. Mol. Physiol. 2006, 290, L622–L645. [Google Scholar] [CrossRef]
- Coldewey, S.M.; Rogazzo, M.; Collino, M.; Patel, N.S.A.; Thiemermann, C. Inhibition of IκB Kinase Reduces the Multiple Organ Dysfunction Caused by Sepsis in the Mouse. Dis. Models Mech. 2013, 6, 1031–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Sig. Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, E. Nuclear Factor–KB and Its Role in Sepsis- Associated Organ Failure. J. Infect. Dis. 2003, 187, S364–S369. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Weng, J.; Fang, D.; Chen, J.; Chen, M. Transcriptomic Study of Lipopolysaccharide-induced Sepsis Damage in a Mouse Heart Model. Exp. Ther. Med. 2020, 20, 3782–3790. [Google Scholar] [CrossRef]
- Wu, T.; Liang, X.; Jiang, Y.; Chen, Q.; Zhang, H.; Zhang, S.; Zhang, C.; Lv, Y.; Xin, J.; Jiang, J.; et al. Comprehensive Transcriptome Profiling of Peripheral Blood Mononuclear Cells from Patients with Sepsis. Int. J. Med. Sci. 2020, 17, 2077–2086. [Google Scholar] [CrossRef]
- Kishore, R.; Verma, S.K. Roles of STATs Signaling in Cardiovascular Diseases. JAK-STAT 2012, 1, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Harhous, Z.; Booz, G.W.; Ovize, M.; Bidaux, G.; Kurdi, M. An Update on the Multifaceted Roles of STAT3 in the Heart. Front. Cardiovasc. Med. 2019, 6, 150. [Google Scholar] [CrossRef] [Green Version]
- Mascareno, E.; El-Shafei, M.; Maulik, N.; Sato, M.; Guo, Y.; Das, D.K.; Siddiqui, M.A.Q. JAK/STAT Signaling Is Associated With Cardiac Dysfunction During Ischemia and Reperfusion. Circulation 2001, 104, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Peng, D.; Shi, W.; Guo, J.; Huo, S.; Men, L.; Zhang, C.; Li, S.; Lv, J.; Lin, L. IL-6/STAT3 Signaling Promotes Cardiac Dysfunction by Upregulating FUNDC1-Dependent Mitochondria-Associated Endoplasmic Reticulum Membranes Formation in Sepsis Mice. Front. Cardiovasc. Med. 2022, 8, 790612. [Google Scholar] [CrossRef]
- Zhuang, L.; Jia, K.; Chen, C.; Li, Z.; Zhao, J.; Hu, J.; Zhang, H.; Fan, Q.; Huang, C.; Xie, H.; et al. DYRK1B-STAT3 Drives Cardiac Hypertrophy and Heart Failure by Impairi.ing Mitochondrial Bioenergetics. Circulation 2022, 145, 829–846. [Google Scholar] [CrossRef]
- Lei, W.; Liu, D.; Sun, M.; Lu, C.; Yang, W.; Wang, C.; Cheng, Y.; Zhang, M.; Shen, M.; Yang, Z.; et al. Targeting STAT3: A Crucial Modulator of Sepsis. J. Cell Physiol. 2021, 236, 7814–7831. [Google Scholar] [CrossRef] [PubMed]
- Rudiger, A.; Dyson, A.; Felsmann, K.; Carré, J.E.; Taylor, V.; Hughes, S.; Clatworthy, I.; Protti, A.; Pellerin, D.; Lemm, J.; et al. Early Functional and Transcriptomic Changes in the Myocardium Predict Outcome in a Long-Term Rat Model of Sepsis. Clin. Sci. 2013, 124, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbaby, S.; Matsuda, N.; Tomita, K.; Hattori, K.; Palikhe, S.; Yokoo, H.; Hattori, Y. Beneficial Effect of STAT3 Decoy Oligodeoxynucleotide Transfection on Organ Injury and Mortality in Mice with Cecal Ligation and Puncture-Induced Sepsis. Sci. Rep. 2020, 10, 15316. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pan, X.; Mao, L.; Pan, H.; Xu, W.; Hu, Y.; Yu, X.; Chen, Z.; Qian, S.; Ye, Y.; et al. Phospho-Tyr705 of STAT3 Is a Therapeutic Target for Sepsis through Regulating Inflammation and Coagulation. Cell Commun. Signal. 2020, 18, 104. [Google Scholar] [CrossRef]
- Hu, Y.; Zhong, W.; Chen, M.; Zhang, Q. Identifying Crucial Genes for Prognosis in Septic Patients. Medicine 2019, 98, e16807. [Google Scholar] [CrossRef]
- Whalen, M.J.; Doughty, L.A.; Carlos, T.M.; Wisniewski, S.R.; Kochanek, P.M.; Carcillo, J.A. Intercellular Adhesion Molecule-1 and Vascular Cell Adhesion Molecule-1 Are Increased in the Plasma of Children with Sepsis-Induced Multiple Organ Failure. Crit. Care Med. 2000, 28, 2600–2607. [Google Scholar] [CrossRef]
- Sosa-Bustamante, G.P.; Amador-Licona, N.; Barbosa-Sabanero, G.; Guízar-Mendoza, J.M.; López-Briones, S.; Mulgado-Águas, C.I.; Torres-Pedroza, A. Intercellular Adhesion Molecules and Mortality for Sepsis in Infants Younger than 1 Year of Life. Rev. Investig. Clin. 2011, 63, 601–606. [Google Scholar]
- Figueras-Aloy, J.; Gómez-López, L.; Rodríguez-Miguélez, J.-M.; Salvia-Roiges, M.D.; Jordán-García, I.; Ferrer-Codina, I.; Carbonell-Estrany, X.; Jiménez-González, R. Serum Soluble ICAM-1, VCAM-1, L-Selectin, and P-Selectin Levels as Markers of Infection and Their Relation to Clinical Severity in Neonatal Sepsis. Am. J. Perinatol. 2007, 24, 331–338. [Google Scholar] [CrossRef]
- Zonneveld, R.; Martinelli, R.; Shapiro, N.I.; Kuijpers, T.W.; Plötz, F.B.; Carman, C.V. Soluble Adhesion Molecules as Markers for Sepsis and the Potential Pathophysiological Discrepancy in Neonates, Children and Adults. Crit. Care 2014, 18, 204. [Google Scholar] [CrossRef] [Green Version]
- Pakos-Zebrucka, K.; Koryga, I.; Mnich, K.; Ljujic, M.; Samali, A.; Gorman, A.M. The Integrated Stress Response. EMBO Rep. 2016, 17, 1374–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.M.; Yang, W.-L.; Wang, P. Endoplasmic Reticulum Stress in Sepsis. Shock 2015, 44, 294–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzing, U.B.; von Loeffelholz, C.; Steidl, R.; Romeike, B.; Winkler, R.; Rauchfuß, F.; Settmacher, U.; Stoppe, C.; Coldewey, S.M.; Weinmann, C.; et al. Endoplasmic Reticulum Stress and the Unfolded Protein Response in Skeletal Muscle of Subjects Suffering from Peritoneal Sepsis. Sci. Rep. 2022, 12, 504. [Google Scholar] [CrossRef]
- Jiao, G.; Hao, L.; Wang, M.; Zhong, B.; Yu, M.; Zhao, S.; Wang, P.; Feng, R.; Tan, S.; Chen, L. Upregulation of Endoplasmic Reticulum Stress Is Associated with Diaphragm Contractile Dysfunction in a Rat Model of Sepsis. Mol. Med. Rep. 2017, 15, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Massaud-Ribeiro, L.; Silami, P.H.N.C.; Lima-Setta, F.; Prata-Barbosa, A. Pediatric Sepsis Research: Where Are We and Where Are We Going? Front. Pediatrics 2022, 10, 829119. [Google Scholar] [CrossRef] [PubMed]
- Randolph, A.G.; McCulloh, R.J. Pediatric Sepsis: Important Considerations for Diagnosing and Managing Severe Infections in Infants, Children, and Adolescents. Virulence 2014, 5, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, J.L.; Cvijanovich, N.Z.; Allen, G.L.; Thomas, N.J.; Freishtat, R.J.; Anas, N.; Meyer, K.; Checchia, P.A.; Lin, R.; Shanley, T.P.; et al. The Influence of Developmental Age on the Early Transcriptomic Response of Children with Septic Shock. Mol. Med. 2011, 17, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Raymond, S.L.; López, M.C.; Baker, H.V.; Larson, S.D.; Efron, P.A.; Sweeney, T.E.; Khatri, P.; Moldawer, L.L.; Wynn, J.L. Unique Transcriptomic Response to Sepsis Is Observed among Patients of Different Age Groups. PLoS ONE 2017, 12, e0184159. [Google Scholar] [CrossRef]
- Mohammed, A.; Cui, Y.; Mas, V.R.; Kamaleswaran, R. Differential Gene Expression Analysis Reveals Novel Genes and Pathways in Pediatric Septic Shock Patients. Sci. Rep. 2019, 9, 11270. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; EL-Manzalawy, Y. Machine Learning Based Refined Differential Gene Expression Analysis of Pediatric Sepsis. BMC Med. Genom. 2020, 13, 122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, J.; Chen, X.; Wang, J.; Fang, F. Transcriptomic Markers in Pediatric Septic Shock Prognosis: An Integrative Analysis of Gene Expression Profiles. Braz. J. Med. Biol. Res. 2021, 54, e10152. [Google Scholar] [CrossRef]
- Rosier, F.; Nuñez, N.F.; Torres, M.; Loriod, B.; Rihet, P.; Pradel, L.C. Transcriptional Response in a Sepsis Mouse Model Reflects Transcriptional Response in Sepsis Patients. Int. J. Mol. Sci. 2022, 23, 821. [Google Scholar] [CrossRef] [PubMed]
- Lambeck, S.; Weber, M.; Gonnert, F.; Mrowka, R.; Bauer, M. Comparison of Sepsis-Induced Transcriptomic Changes in a Murine Model to Clinical Blood Samples Identifies Common Response Patterns. Front. Microbiol. 2012, 3, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efron, P.A.; Darden, D.B.; Wang, Z.; Nacionales, D.C.; Lopez, M.-C.; Hawkins, R.B.; Cox, M.C.; Rincon, J.C.; Ungaro, R.; Dirain, M.L.; et al. Transcriptomic Responses from Improved Murine Sepsis Models Can Better Mimic Human Surgical Sepsis. FASEB J. 2021, 35, e21156. [Google Scholar] [CrossRef] [PubMed]
- Fink, M.P. Animal Models of Sepsis. Virulence 2014, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Stortz, J.A.; Raymond, S.L.; Mira, J.C.; Moldawer, L.L.; Mohr, A.M.; Efron, P.A. Murine Models of Sepsis and Trauma: Can We Bridge the Gap? ILAR J. 2017, 58, 90–105. [Google Scholar] [CrossRef] [Green Version]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic Responses in Mouse Models Poorly Mimic Human Inflammatory Diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [Green Version]
- Van der Laarse, S.A.M.; Leney, A.C.; Heck, A.J.R. Crosstalk between Phosphorylation and O-GlcNAcylation: Friend or Foe. The FEBS J. 2018, 285, 3152–3167. [Google Scholar] [CrossRef] [Green Version]
- Macauley, M.S.; Whitworth, G.E.; Debowski, A.W.; Chin, D.; Vocadlo, D.J. O-GlcNAcase Uses Substrate-Assisted Catalysis: Kinetic Analysis and Development of Highly Selective Mechanism-Inspired Inhibitors. J. Biol. Chem. 2005, 280, 25313–25322. [Google Scholar] [CrossRef] [Green Version]
- Dupas, T.; Denis, M.; Dontaine, J.; Persello, A.; Bultot, L.; Erraud, A.; Vertommen, D.; Bouchard, B.; Tessier, A.; Rivière, M.; et al. Protein O -GlcNAcylation Levels Are Regulated Independently of Dietary Intake in a Tissue and Time-specific Manner during Rat Postnatal Development. Acta Physiol. 2021, 231, e13566. [Google Scholar] [CrossRef]
- Charpentier, E.; Cornec, M.; Dumont, S.; Meistermann, D.; Bordron, P.; David, L.; Redon, R.; Bonnaud, S.; Bihouée, A. 3’ RNA Sequencing for Robust and Low-Cost Gene Expression Profiling; Protocol Exchange; Research Square: Durham, NC, USA, 2021. [Google Scholar]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dupas, T.; Persello, A.; Blangy-Letheule, A.; Denis, M.; Erraud, A.; Aillerie, V.; Leroux, A.A.; Rivière, M.; Lebreton, J.; Tessier, A.; et al. Beneficial Effects of O-GlcNAc Stimulation in a Young Rat Model of Sepsis: Beyond Modulation of Gene Expression. Int. J. Mol. Sci. 2022, 23, 6430. https://doi.org/10.3390/ijms23126430
Dupas T, Persello A, Blangy-Letheule A, Denis M, Erraud A, Aillerie V, Leroux AA, Rivière M, Lebreton J, Tessier A, et al. Beneficial Effects of O-GlcNAc Stimulation in a Young Rat Model of Sepsis: Beyond Modulation of Gene Expression. International Journal of Molecular Sciences. 2022; 23(12):6430. https://doi.org/10.3390/ijms23126430
Chicago/Turabian StyleDupas, Thomas, Antoine Persello, Angélique Blangy-Letheule, Manon Denis, Angélique Erraud, Virginie Aillerie, Aurélia A. Leroux, Matthieu Rivière, Jacques Lebreton, Arnaud Tessier, and et al. 2022. "Beneficial Effects of O-GlcNAc Stimulation in a Young Rat Model of Sepsis: Beyond Modulation of Gene Expression" International Journal of Molecular Sciences 23, no. 12: 6430. https://doi.org/10.3390/ijms23126430