
Cross-Talk between the (Endo)Cannabinoid and Renin-Angiotensin Systems: Basic Evidence and Potential Therapeutic Significance

 , ,
, ,
Abstract
:1. Introduction
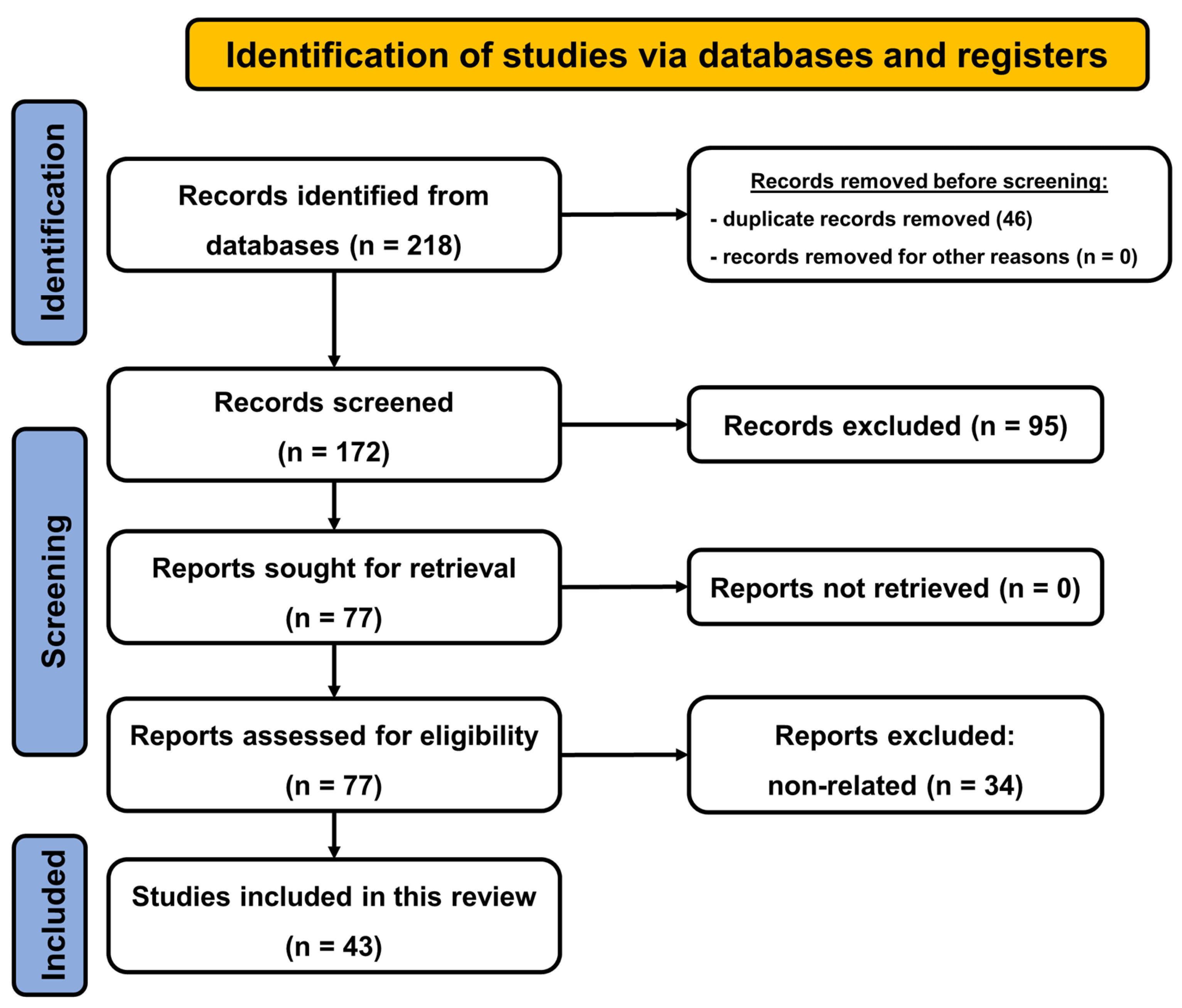
2. Materials and Methods
3. Results and Discussion
3.1. Components and Main Effects of (Endo)Cannabinoid and Renin-Angiotensin Systems
3.1.1. (Endo) Cannabinoid System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Class | Mechanism | Approved Drug | Indications |
|---|---|---|---|---|
| (endo)cannabinoid system | agonists | unselective cannabinoid receptor agonist | dronabinol (Δ9-tetrahydrocannabinol, THC) | anorexia and weight loss in HIV patients, nausea and vomiting in cancer chemotherapy |
| unselective cannabinoid receptor agonist | nabilone | like dronabinol | ||
| antagonists | CB1 receptor antagonist | rimonabant 2 | obesity | |
| other | weak activity towards the cannabinoid system, antioxidant drug | cannabidiol | neuropathic pain; Lennox-Gastaut and Dravet syndrome | |
| (see dronabinol and cannabidiol) | nabiximols (1:1 formulation of dronabinol and cannabidiol) | neuropathic pain in multiple sclerosis, intractable cancer pain | ||
| renin-angiotensin system | agonists | unselective AT receptor agonist | angiotensin II | increase in blood pressure in adults with septic or other distributive shock |
| antagonists | renin inhibitor | e.g., aliskiren | essential hypertension | |
| AT1 receptor antagonist | e.g., candesartan, valsartan | essential hypertension, congestive heart failure | ||
| aldosterone receptor antagonist | e.g., eplerenone | congestive heart failure | ||
| other | angiotensin converting enzyme inhibitor | e.g., perindopril | essential hypertension, congestive heart failure |
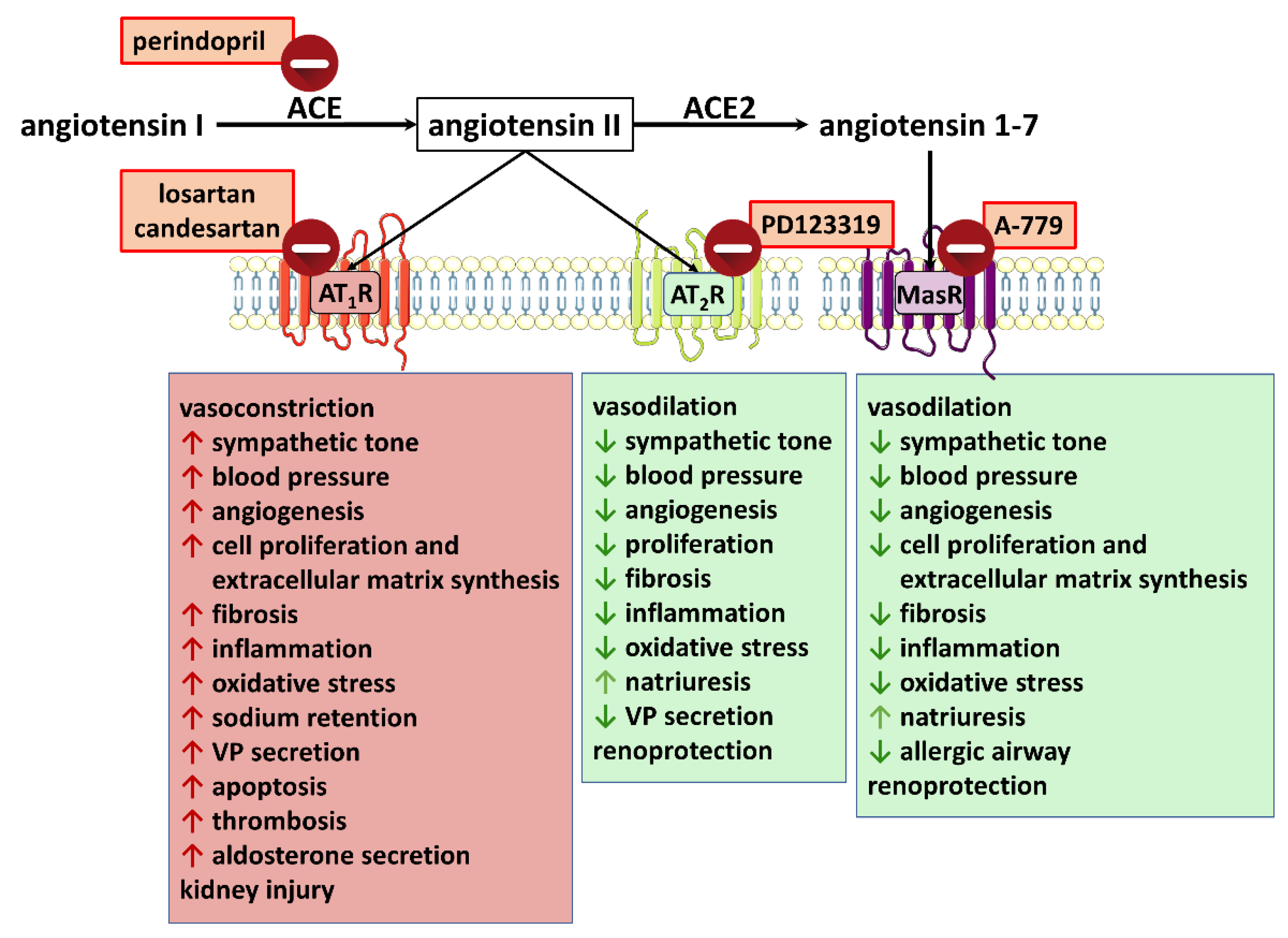
3.1.2. Renin-Angiotensin System
3.2. Examples of Cross-Talk between the (Endo) Cannabinoid and Renin-Angiotensin Systems
| Species | Model | Agonist Concentration (μM) or Dose | Effect | (Functional) Antagonist; Concentration In Vitro (μM) or Dose | Influence on the Agonist Effect | Final Conclusion of the Authors | References |
|---|---|---|---|---|---|---|---|
| cells: Chinese hamster; human; African green monkey | CHO; HEK293; COS7 cells (co-expressing AT1Rs and CB1Rs) from ovaries, kidneys, and fibroblasts, respectively | Ang II (0.1) | ↑2-AG ↔AEA ↑Go protein activation | AM251 (10) THL (1) | ↓Ang II-induced Go protein activation | AT1R stimulation leads to DAGL-mediated transactivation of CB1Rs in an autocrine and paracrine manner | [33,34] |
| cells: mouse | neuro2A cells, a neuroblastoma cell line co-expressing CB1Rs and AT1Rs | Ang II (0.01–10) | ↑pERK levels via Gαi instead of Gαq the expression of AT1R shifts CB1Rs from an intracellular compartment to the plasma membrane | losartan CB1R-targeting siRNA RIM (1) THL (1) HU210 (0.0001) | Ang II-induced ↑pERK ↓ by losartan, CB1R-targeting siRNA, RIM, and THL; ↑ by HU210 (occurring in the presence of a very low non-signaling concentration of Ang II only) | AT1Rs and CB1Rs form receptor heteromers; blocking CB1R activity prevented the Ang II-mediated pathologic effect | [35] |
| cells: rats | hepatic stellate cells from control rats (cHSCs) and rats treated with ethanol for 8 months (eHSCs) | Ang II (1) | CB1R, AT1R and AT1R-CB1 heteromer levels in eHSCs > cHSCs; ↑pERK levels, ↑mitogenic and ↑profibrogenic markers in eHSCs > cHSCs | RIM (1) | ↓Ang II-induced changes | ||
| Blood Vessels | |||||||
| rats Wistar | aortic VSMCs | Ang II (0.1) | ↑2-AG level ↑Ca2+ signal | THL (1) JZL184 (1) | ↓ and ↑ of Ang II-induced 2-AG formation and Ca2+ signal by THL and JZL184, respectively | Ang II stimulates eCB (2-AG) release from the vascular wall that reduces the vasoconstrictor effects of Ang II via CB1R activation (eCBs act as protective negative feedback in response to Ang II) | [36] |
| rats and/or mice | aortic rings from rats aortic rings from CB1−/− and WT mice | Ang II (0.001–0.1) | concentration-dependent contraction | WIN-2 (10) O2050 (1) THL (1) JZL184 (1) | vasodilation to WIN-2; not detected in CB1−/− O2050, THL↑ , and JZL184↓ vasoconstrictor effect of Ang II; amplificatory effect of O2050 in WT only | ||
| rats and/or mice | skeletal muscle arterioles, saphenous arteries | Ang II (0.001–0.1) | concentration-dependent contraction | WIN-2 (1) O2050 (1) RIM (1) AM251 (1) THL (1) | vasodilation to WIN-2; not detected in CB1−/− ↑vasoconstrictor effect of Ang II in WT but not in CB1−/− | Ang II stimulates eCB release from the vascular wall that reduces the vasoconstrictor effects of Ang II via CB1R activation (eCBs act as protective negative feedback in response to Ang II) | [37] |
| rats Wistar | intramural coronary resistance arterioles | Ang II (0.0001–10) | concentration-dependent contraction | WIN-2 (0.0001–1) O2050 (1) THL (1) | vasodilatation to WIN-2 reduced by O2050 and AM251 ↑vasoconstrictor effect of Ang II | Ang II stimulates eCB release from the vascular wall that reduces the vasoconstrictor effects of Ang II via CB1R activation (eCBs act as protective negative feedback in response to Ang II) | [38] |
| rats Wistar | pulmonary arteries | Ang II (0.0001–0.03) | concentration-dependent contraction | AM251 (1) RHC80267 (40) JZL184 (1) URB597 (1) | AM251 and RHC80267 ↑ but JZL184 ↓ vasoconstrictor effect of Ang II; URB597 ↔ | Ang II stimulates eCB (2-AG) release from the vascular wall that reduces the vasoconstrictor effects of Ang II via CB1R activation (eCBs act as protective negative feedback in response to Ang II) | [39] |
| rats | uterine artery from hypertensive TgA and normotensive SD rats | Ang II (0.00001–0.01) | concentration-dependent contraction, stronger in TgA | URB597 (1) JZL184 (1) RIM (1) | ↓responses to Ang II in SD and TgA ↓responses to Ang II in TgA ↔responses to Ang II in SD and TgA | eCBs reduce the Ang II-induced contraction in a CB1R-independent manner in the early stages of hypertensive pregnancy (eCBs act as protective negative feedback in response to Ang II) | [40] |
| rats SD mice | VSMCs from rat and mouse thoracic aortas with CB1R expression | Ang II (1) | ↑ROS production ↑NADPH oxidase activity | RIM (0.1–1) or AM251 (1) CP55940 (1) | ↓AT1Rs and decrease in the Ang II-induced ↑ROS production and ↑NADPH oxidase activity ↑AT1Rs | CB1R inhibition (in vitro and in vivo) has atheroprotective effects by down-regulation of AT1Rs, decreased vascular ROS, and thus improved endothelial function in hypercholesterolemic ApoE−/− mice | [41] |
| mice | ApoE−/− treated with a cholesterol-rich diet | development of atherosclerotic plaques, ↓aorta relaxation, ↔aortic AT1R level | RIM (10 mg/kg/day; p.o.) for 7 weeks | ↓aortic AT1Rs and improvement of endothelial function, no effect on atherosclerotic plaques | |||
| Heart | |||||||
| rats SD | isolated Langendorff-perfused hearts | Ang II (0.001–0.1) 2-AG (1) WIN-2 (1) | ↓CF and moderate negative inotropic effect 2-AG and WIN-2: ↑CF WIN-2: negative inotropic effect | O2050 (1) + Ang II orlistat (10) + Ang II | ↓cardiac effects of Ang II | besides direct cardiac responses, Ang II induces indirect ones via eCBs (probably 2-AG) activating CB1Rs:
| [42] |
| mice | streptozotocin-induced diabetes | diabetic cardiomyopathy | ↑myocardial CB1 and AT1R expression and AEA level connected with cardiac dysfunction, inflammation, oxidative/nitrative stress | RIM or AM281 (10 mg/kg; i.p. daily for 11 weeks) or CB1R deletion (CB1−/− mice) | pharmacological inhibition or genetic deletion of CB1Rs—improvement of diabetic cardiac dysfunction connected with ↓AT1Rs and CB1Rs in LV | overactivation of the eCB system and CB1Rs may play an important role in the pathogenesis of diabetic cardiomyopathy by facilitating AT1R expression and signaling | [43] |
| rats Wistar | isolated Langendorff-perfused hearts underwent ischemia + reperfusion | ischemia and reperfusion | ↑stroke size, ↓ventricular function; ↑ cardiac AT1R level and ↔ cardiac AT2R level | CBD (5 mg/kg; i.p. daily for 10 days) | ↓stroke size and ↑ventricular function; ↓ cardiac AT1R level and ↑ cardiac AT2R level | cardioprotective effect of CBD might result from an increase in cardioprotective AT2Rs stimulating counter-regulatory effects on the AT1Rs | [44] |
| mice | Ang II-induced fibrosis and inflammation | Ang II infusion (1 µg/kg/min [preventive] or 500 [therapeutic] for 4 weeks) | fibrosis and inflammation in the heart, aorta, lung, kidney, and skin | EHP-101 (2, 5 or 20 mg/kg for 4 or 2 weeks) | ↓cardiac, aortic, lung, kidney, and skin fibrosis and inflammation in the preventive or therapeutic model | EHP-101 (dual agonist of CB2Rs and PPARγ) can alleviate cardiac, aortic, lung, kidney, and skin inflammation induced by Ang II | [45] |
| Blood Pressure | |||||||
| mice CB1−/− CB1+/+ | anesthetized | Ang II (1 μg/kg/min) | ↑BP in WT and CB1−/− | O2050 (10 mg/kg; p.o.) | ↑ pressor effect of Ang II in WT, not in CB1−/− | confirmation of in vitro experiments on isolated arteries that Ang II stimulates release of eCBs (2-AG) from the vascular wall that reduce vasoconstrictor effects of Ang II via CB1R activation | [36] |
| rats SD | conscious | Ang II-induced hypertension (60 ng/min; s.c. for 10–12 days) | ↑BP | AM251 (3 mg/kg; i.v.) URB597 (10 mg/kg; i.v.) in pentobarbital- anaesthetized rats | AM251 ↑BP and URB597 ↓BP in Ang II-induced hypertension but not in normotension | the Ang II-induced hypertension is diminished by eCBs acting at CB1Rs; effect of URB597 reduced by AM251 | [19] |
| rats Wistar | conscious | Ang II (500 ng/kg/h) + VP (50 ng/kg/h for 4 days)- induced hypertension | ↑BP | AEA (3 mg/kg) URB597 (3 mg/kg) WIN-2 (150 μg/kg) AM251 (3 mg/kg) | AEA, WIN-2 ↓BP in Ang II-VP-induced hypertension; URB597 enhanced the effect of AEA; AM251 blocked the effect of WIN-2 | the Ang II-VP induced hypertension might be diminished by eCBs acting at CB1Rs | [46] |
| rats SHR WKY | conscious | BP was higher in SHR than in WKY | RIM 3 mg/kg i.v. URB597 1.7 mg/kg i.v. | RIM ↑BP and URB597 ↓BP in SHR but not in normotensive WKY | in SHR in which RAS is overactivated eCBs acting at CB1Rs reduce BP | [19] | |
| rats | conscious (mRen2)27 hypertensive rats or normotensive SD | (mRen2)27: higher RAS activity | RIM (10 mg/kg; p.o. acutely or daily for 28 days) | acutely: ↓BP and ↓HR in hypertensive but not in SD chronically: ↓BP and ↓HR; ↔plasma Ang II, ↔Ang 1-7; ↔ACE; improvement of sympathetic and parasympathetic BRS | upregulated ECS contributes to hypertension and impaired autonomic function in this Ang II-dependent model; systemic CB1R blockade may be an effective therapy for Ang II-dependent hypertension and the associated metabolic syndrome | [47] | |
| rats | anaesthetized (mRen2)27 hypertensive, ASrAOGEN and SD rats | (mRen2)27: higher RAS activity; AsrAO-GEN: low glial angiotensinogen | levels in NTS: 2-AG: (mRen2)27 > SD > ASrAOGEN; AEA: (mRen2)27 ≈ SD ≈ ASrAOGEN dorsal medulla: CB1: ASrAOGEN < (mRen2)27 ≈ SD; CB2: no differences | RIM (0.36 and 36 pmol/rat; NTS) | ↑BRS in (mRen2)27; ↓BRS in ASrAOGEN; ↔BRS in SD | upregulated brain ECS in Ang II-dependent hypertension may contribute to the impaired baroreceptor sensitivity in this model of hypertension | [48] |
| rats | obese fa/fa Zucker rats and control lean fa/+ Zucker rats; isoflurane-anaesthetized | acute Ang II (30 and 100 ng/kg, i.v.) | stronger pressor response in obese than in lean rats | RIM (3 or 10 mg/kg, p.o.) for 12 months | normalized the acute pressor response to Ang II in obese rats to the level of lean rats | authors suggest that chronic CB1R blockade by RIM might reduce vascular AT1R expression; an indirect mechanism related to the decrease in the cholesterol level should also be taken under consideration | [49] |
| rats SHR WKY | conscious | SHR in comparison to WKY: higher BP, carotid, mesenteric artery: ↑AT1Rs, ↑ACE kidney: ↔AT1Rs, ↔AT2Rs, ↑ACE | PEA (30 mg/kg; s.c. for 5 weeks) | BP in SHR↓ SHRarteries: ↓AT1Rs, ↓ACE level SHR kidney: ↓AT1Rs, ↑AT2Rs, and ↓ACE level associated with ↓oxidative and nitrosative stress | PEA lowers BP and protects against hypertensive renal injury partially via reduction in vascular AT1Rs and Ang II-mediated effects and via modulation of the RAS, leading the AT1/AT2 balance towards an anti-hypertensive status | [50,51] | |
| rats WKY | cultured lymphocytes from WKY | Ang II (0.01–1) | concentration-dependent ↓AEA transporter activity and ↑ROS level | losartan (10 and 100) | ↓Ang II effects on AEA transporter activity and ROS level | Ang II plays a critical role in mediating the decrease in AEA transporter activity in SHR; probably via AT1Rs | [52] |
| rats SHR WKY | conscious | SHR: ↑plasma Ang II and ↑AEA level; ↓AEA transporter activity in comparison to WKY | losartan (15 or 30 mg/kg; p.o. for 2 weeks) | restoration of reduced AEA transporter activity; ↓plasma AEA level | |||
| Nervous System | |||||||
| rats Wistar | urethane- anesthetized | Ang II (0.14 nmol/rat; PVN) | ↑BP | AM251 (0.48 nmol/rat; PVN) | AM251 reduced the Ang II-mediated BP increase and slightly increased BP by itself | Ang II-induced hypertension involves CB1Rs in the PVN | [53] |
| rats Wistar | urethane- anaesthetised (microinjection into the PVN, doses in nmol/rat) | CP55940 (0.1) CP55940 (0.1) + AM251 (3 μmol/kg; i.v.) | ↓BP, ↓HR ↑BP, ↑HR | losartan (10 μmol/kg; i.v.) losartan (10 μmol/kg; i.v.) | no effect reversed ↑BP, ↑HR to ↓BP, ↓HR | presynaptic inhibitory CB1Rs on GABAergic neurons in the PVN activated by eCBs released in response to Ang II modify the glutamatergic neurotransmission enhanced by presynaptic AT1R activation | [54] |
| rats SHR WKY | conscious (all compounds microinjected into the PVN, nmol/rat) | Ang II (0.03) or CP55940 (0.1) + AM251 (3 μmol/kg; i.v.) | ↑BP stronger in SHR than in WKY | losartan (20) PD123319 (10) AM251 (30) | ↓pressor effect of Ang II and CP55940 ↓pressor effect of Ang II and CP55940 ↓pressor effect of Ang II | mutual interaction in the PVN between CB1Rs and receptors for Ang II responsible for stimulation of the pressor response (probably via stimulation of CB1R by eCBs released in response to Ang II) | [55] |
| mice | magnocellular neurosecretory cells from the supraoptic nucleus | Ang II (0.1) | ↑frequency of mEPSCs | AM251 (2) | ↑effect of Ang II | eCBs released in response to Ang II modulate the excitatory synaptic inputs via negative feedback | [56] |
| rats Wistar mice CB1+/+ CB1−/− | conscious | Ang II (191 pmol/rat; i.c.v.) Ang II (191 pmol/mouse i.c.v.) | ↓ethanol-induced gastric lesions (reduced by candesartan 5.2 and 31.7 nmol/rat; i.c.v.) gastroprotection in CB1+/+ as opposed to CB1−/− | AM251 (1.8 nmol/rat; i.c.v.) THL (0.2 nmol/rat; i.c.v.) | inhibition of the gastroprotective effect of Ang II | Ang II stimulates eCB release via activation of central AT1R receptors, and activation of CB1Rs induces gastroprotection in a vagus-mediated mechanism (inhibition by vagotomy and atropine) | [57] |
| mice CB1+/+ CB1−/ | response of the chorda tympani (CT) nerve in anesthetized mice | CB1+/+: Ang II (100–5000 ng/kg; i.p.) | gustatory nerve responses ↓ to NaCl and ↑ to sweeteners, blocked by candesartan | CB1–/–: Ang II (100–5000 ng/kg; i.p.) | gustatory nerve responses ↓ to NaCl and ↔ to sweeteners | enhancing effect of Ang II on sweet taste responses mediated by AT1 and CB1Rs; authors suggest that Ang II, via AT1Rs, stimulates the release of 2-AG that may act as an autocrine enhancer for CB1Rs on sweet taste cells | [58] |
| rats SHR WKY | astrocytes basal CB1R densities: brainstem: SHR<WKY cerebellum: SHR>WKY | Ang II (0.1) | SHR: ↓CB1R and ↑CB1R densities and phosphorylation in brainstem and cerebellar astrocytes, respectively; opposite effects in WKY | losartan (10) PD123319 (10) ACEA (0.01) | - effects of Ang II were inhibited by losartan (brainstem) and by losartan and PD123319 (cerebellum) - ACEA reduced the AT1R-mediated MAPK activation in brainstem and cerebellar astrocytes | Ang II, mostly via the AT1R, is capable of altering CB1R expression and phosphorylation in astrocytes isolated from the brainstem and cerebellum under hyper- and normotensive conditions; possible role in neuroinflammatory and attention-deficit hyperactivity disorders, respectively | [59,60] |
| rats SHR WKY | astrocytes isolated from the brainstem and from cerebellum | Ang II (0.1) | ↓IL-10 and ↑IL-1β gene expression in astrocytes from both brain regions of SHR and WKY | ACEA (0.01) | co-treatment of Ang II and ACEA resulted in the neutralization of Ang II-mediated effect in WKY but not SHR | Ang II and ACEA have opposing roles in the regulation of inflammatory gene signature in astrocytes isolated from SHR and Wistar rats (possible functional antagonism) | [61] |
| mice CB2−/− CB2+/+ | hippocampus slices | CB2−/−: ↓ACE level, and ↑aβP in comparison to WT | CB2R deletion:↑aβ neurotoxicity associated with ↓level of ACE (that degrades aβ) | activation of CB2Rs increases ACE level that degrades aβ; possible significance in Alzheimer’s disease | [62] | ||
| N2a cells overexpressing aβP | JWH133 | ↑ACE level, ↓aβP | AM630 | all JWH133 effects were attenuated | |||
| Kidney | |||||||
| humans | podocytes | Ang II (0.1) | ↑AEA, ↑2-AG ↑AT1Rs and CB1Rs | JD5037 (100) or losartan (10) | ↓ all changes induced by Ang II | peripheral CB1R blockade might possess therapeutic potential in disease(s) connected with enhanced RAS | [63] |
| rats | Zucker diabetic fatty rats with nephropathy; control lean rats | diabetic compared to lean rats | ↑plasma Ang II and aldosterone levels; ↓AT1Rs in renal cortex | JD5037 (3 mg/kg p.o. for 3 months) losartan (20 mg/kg p.o. for 28 days) | ↓plasma Ang II and aldosterone levels; ↓AT1Rs in renal cortex ↔plasma Ang II and ↓aldosterone levels; ↓CB1Rs in renal cortex | ||
| mice | streptozotocin- induced diabetic nephropathy | ↑glomerular CB1 and ↑AT1Rs; ↔CB2Rs | AM6545 (10 mg/kg; i.p.) alone or together with perindopril (2 mg/kg; p.o.) for 14 weeks | Single treatments ↔glomerular CB1-, CB2Rs, and ↓AT1Rs; ↓progression of albuminuria, down-regulation of nephrin and podocin, ↓inflammation, and ↓expression of markers of fibrosis Combinedtreatment ↔glomerular CB1-, CB2Rs and ↓AT1Rs; also reversal of albuminuria | The superior effect of dual therapy (peripheral CB1R antagonist + ACE inhibitor) on albuminuria, nephrin loss, and inflammation suggest that CB1R blockade may be a valuable option as an additional therapy, although single and combined treatment only reduce glomerular AT1Rs without affecting CB1Rs and CB2Rs. | [64] | |
3.2.1. Angiotensin II Causes Transactivation of Cannabinoid-Mediated Effects
3.2.2. Other Forms of Interaction between Angiotensin II and Cannabinoid-Mediated Effects
3.2.3. Typology and Potential Therapeutic Significance of the Cross-Talk between (Endo) Cannabinoids and the Renin-Angiotensin System
3.3. Interaction between Angiotensin 1-7 and (Endo) Cannabinoid-Mediated Effects
| Species | Model | Basal Treatment (Concentration (μM) or Dose) | Effect | Intervention (Concentration (μM) or Dose) | Influence on the Agonist Effect | Final Conclusion of the Authors | References |
|---|---|---|---|---|---|---|---|
| rats | ovariectomized pseudopregnant | steroid treatment and bolus of oil leading to decidualization | uterus: ↓CB1Rs, ↑CB2Rs, ↔MAGL, ↓FAAH | Ang 1-7 (24 μg/kg/h; i.u. for 5 days) | uterus: ↑CB1Rs, ↑CB2Rs, ↑MAGL, ↔FAAH | Ang 1-7 augments the expression of CB1Rs, CB2Rs, and MAGL in the decidualized uterus and thus may interfere with early events of decidualization | [92] |
| rats SHR WKY | conscious (compounds microinjected into the PVN, doses in nmol/rat; if not stated otherwise) | Ang 1-7 (0.03) or CP55940 (0.1) + AM251 3 μmol/kg i.v. | both treatments: ↑BP stronger in SHR than in WKY | A-779 (3) AM251 (30) | ↓pressor effect of Ang 1-7 and CP55940 ↓pressor effect of Ang 1-7 | mutual interaction in the PVN between the CB1Rs and the receptors for Ang 1-7 responsible for stimulation of the pressor response | [55] |
3.4. Influence of Cannabinoids on ACE2 Activity and Other Parameters Relevant in the Fight against COVID-19
| Model | Agonists | Effects | Final Conclusion of Authors | References |
|---|---|---|---|---|
| human iPSC- cardiomyocytes infected with SARS-CoV-2 | WIN55212-2 | ↔ ACE2 levels; ↔viral infection and replication; ↓release of proinflammatory cytokines and cytotoxic damage | therapeutic potential of cannabinoids in protecting the heart against SARS-CoV-2 infection is not related to modification of ACE2 levels | [99] |
| in silico docking studies | CBD THC CBN | CBD: hACE2 (↓), main virus protease activity ↓ THC: hACE2 and main virus protease (↓) CBN: inactive | THC and CBD might inhibit the SARS-CoV-2 infection via their influence on hACE2 and viral proteases | [101] |
| in silico docking studies | 8 phyto-compounds derived from cannabis, including CBD, THC, and CVN | CBD and CVN showed the strongest potency in docking to ACE2, TMPRSS2, NRP1, IL-6, and TNF-α | CBD and CVN may be beneficial for the treatment of COVID-19 and post-COVID-19 neuronal symptoms | [102] |
| artificial 3D human models of oral, airway, and intestinal tissues treated with TNF-α and IFN-γ | 13 high-CBD Cannabis sativa extracts | ↓ACE2 and TMPRSS2 in oral, lung, and intestinal epithelia constituting important routes of SARS-CoV-2 invasion | Cannabis sativa extracts may become a useful and safe addition to the prevention/treatment of COVID-19 as an adjunct therapy; the modulation of ACE2 levels may be an effective strategy for decreasing disease susceptibility | [103] |
| alveolar epithelial A549 cell line macrophage cell line KG1 | extract from Cannabis sativa strain Arbel (CBD, CBG, THVC, and terpenes) | A549: ↓ACE2 expression together with ↓IL-6, IL-8, CCL2 macrophage: ↑IL-6 and ↑IL-8 levels | further studies are needed to determine the therapeutic significance of cannabis in COVID-19 treatment due to its positive (A549 cells) and negative effects (macrophages) | [104] |
| human colon Caco-2 cell line | CBD | ↓ACE2 (concentration-dependent) ↑cell viability, ↓all proinflammatory markers | further studies are needed to clarify the consequences of ACE2 down-regulation and its impact on susceptibility to SARS-CoV-2 | [105] |
| human lung fibroblast WI-38 | high-CBD/low-THC cannabis extracts CBD | ↓ACE2, TMPRSS, COX2, IL-6, and IL-8 ↓ACE2, TMPRSS | further studies are needed to identify the proper ratios of a combination of single ingredients to find an ideal formulation for future potential clinical studies/use | [31] |
| human H1299 lung adenocarcinoma cells | industrial hemp essential oil: E-caryophyllene and α-pinene were the prominent terpenes and CBDA was the main terpenophenol | ↓gene expression of ACE2 and TMPRSS2 | hemp essential oils are promising agents to be further investigated with the final goal of optimizing their use in protective devices for counteracting the SARS-CoV-2 virus entry into the human host | [106] |
| Caco-2 293T-ACE2 Vero E6 cell lines | extracts of hemp and isolates of specific cannabinoids: CBDA and CBGA | cannabinoid acids (CBDA and CBGA) lower SARS-CoV-2 entry into Vero E6 cells through spike binding | CBDA and CBGA (allosterically) block cellular entry of pseudovirus and live SARS-CoV-2 alpha variant B.1.1.7 and beta variant B.1.351 | [107] |
3.5. Pharmacokinetic Interactions between Drugs Acting on the (Endo)Cannabinoid and Renin-Angiotensin Systems
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aleksova, A.; Gagno, G.; Sinagra, G.; Beltrami, A.P.; Janjusevic, M.; Ippolito, G.; Zumla, A.; Fluca, A.L.; Ferro, F. Effects of SARS-CoV-2 on Cardiovascular System: The Dual Role of Angiotensin-Converting Enzyme 2 (ACE2) as the Virus Receptor and Homeostasis Regulator-Review. Int. J. Mol. Sci. 2021, 22, 4526. [Google Scholar] [CrossRef]
- Hrenak, J.; Simko, F. Renin-Angiotensin System: An Important Player in the Pathogenesis of Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2020, 21, 8038. [Google Scholar] [CrossRef]
- Yalcin, H.C.; Sukumaran, V.; Al-Ruweidi, M.K.A.A.; Shurbaji, S. Do Changes in ACE-2 Expression Affect SARS-CoV-2 Virulence and Related Complications: A Closer Look into Membrane-Bound and Soluble Forms. Int. J. Mol. Sci. 2021, 22, 6703. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, B.; Baranowska-Kuczko, M.; Kicman, A.; Schlicker, E. Opportunities, Challenges and Pitfalls of Using Cannabidiol as an Adjuvant Drug in COVID-19. Int. J. Mol. Sci. 2021, 22, 1986. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Steffens, S.; Haskó, G.; Schindler, T.H.; Kunos, G. Cardiovascular effects of marijuana and synthetic cannabinoids: The good, the bad, and the ugly. Nat. Rev. Cardiol. 2018, 15, 151–166. [Google Scholar] [CrossRef]
- Pacher, P.; Kogan, N.M.; Mechoulam, R. Beyond THC and Endocannabinoids. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 637–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toczek, M.; Malinowska, B. Enhanced endocannabinoid tone as a potential target of pharmacotherapy. Life Sci. 2018, 204, 20–45. [Google Scholar] [CrossRef] [PubMed]
- Leo, L.M.; Abood, M.E. CB1 Cannabinoid Receptor Signaling and Biased Signaling. Molecules 2021, 26, 5413. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From Phytocannabinoids to Cannabinoid Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Roles Through Complex Pharmacology. Physiol. Rev. 2016, 96, 1593–1659. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.K.; Devi, L.A. The highs and lows of cannabinoid receptor expression in disease: Mechanisms and their therapeutic implications. Pharmacol. Rev. 2011, 63, 461–470. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Kicman, A.; Pędzińska-Betiuk, A.; Kozłowska, H. The potential of cannabinoids and inhibitors of endocanna binoid degradation in respiratory diseases. Eur. J. Pharmacol. 2021, 911, 174560. [Google Scholar] [CrossRef]
- Di Marzo, V. Targeting the endocannabinoid system: To enhance or reduce? Nat. Rev. Drug Discov. 2008, 7, 438–455. [Google Scholar] [CrossRef]
- O’Sullivan, S.E.; Yates, A.S.; Porter, R.K. The Peripheral Cannabinoid Receptor Type 1 (CB1) as a Molecular Target for Modulating Body Weight in Man. Molecules 2021, 26, 6178. [Google Scholar] [CrossRef] [PubMed]
- Finn, D.P.; Haroutounian, S.; Hohmann, A.G.; Krane, E.; Soliman, N.; Rice, A.S.C. Cannabinoids, the endocannabinoid system, and pain: A review of preclinical studies. Pain 2021, 162, S5–S25. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Álvarez, A.; Romero-Cordero, K.; Macías-Triana, L.; Tatum-Kuri, A.; Vera-Barrón, A.; Budde, H.; Machado, S.; Yamamoto, T.; Imperatori, C.; Murillo-Rodríguez, E. The synthetic CB1 cannabinoid receptor selective agonists: Putative medical uses and their legalization. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 110, 110301. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Aguirre, C.; Cinar, R.; Rocha, L. Targeting Endocannabinoid System in Epilepsy: For Good or for Bad. Neuroscience 2022, 482, 172–185. [Google Scholar] [CrossRef]
- Bátkai, S.; Pacher, P.; Osei-Hyiaman, D.; Radaeva, S.; Liu, J.; Harvey-White, J.; Offertáler, L.; Mackie, K.; Rudd, M.A.; Bukoski, R.D.; et al. Endocannabinoids acting at cannabinoid-1 receptors regulate cardiovascular function in hypertension. Circulation 2004, 110, 1996–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kloza, M.; Harasim-Symbor, E.; Biernacki, M.; Kasacka, I.; Malinowska, B. Beneficial Changes in Rat Vascular Endocannabinoid System in Primary Hypertension and under Treatment with Chronic Inhibition of Fatty Acid Amide Hydrolase by URB597. Int. J. Mol. Sci. 2021, 22, 4833. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2017. [Google Scholar] [CrossRef]
- Broyd, S.J.; Van Hell, H.H.; Beale, C.; Yücel, M.; Solowij, N. Acute and Chronic Effects of Cannabinoids on Human Cognition—A Systematic. Review. Biol. Psychiatry 2016, 79, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.F.; Nappe, T.M. Cannabinoid Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- An, D.; Peigneur, S.; Hendrickx, L.A.; Tytgat, J. Targeting Cannabinoid Receptors: Current Status and Prospects of Natural Products. Int. J. Mol. Sci. 2020, 21, 5064. [Google Scholar] [CrossRef]
- Bekassy, Z.; Lopatko Fagerström, I.; Bader, M.; Karpman, D. Crosstalk between the renin-angiotensin, complement and kallikrein-kinin systems in inflammation. Nat. Rev. Immunol. 2021, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Labandeira-Garcia, J.L.; Labandeira, C.M.; Valenzuela, R.; Pedrosa, M.A.; Quijano, A.; Rodriguez-Perez, A.I. Drugs Modulating Renin-Angiotensin System in COVID-19 Treatment. Biomedicines 2022, 10, 502. [Google Scholar] [CrossRef]
- Verma, K.; Pant, M.; Paliwal, S.; Dwivedi, J.; Sharma, S. An Insight on Multicentric Signaling of Angiotensin II in Cardiovascular system: A Recent Update. Front. Pharmacol. 2021, 12, 734917. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sun, Q.; Yuan, S.; Wang, J.; Li, F.; Gao, H.; Chen, X.; Yang, R.; Xu, J. AT1 Receptors: Their Actions from Hypertension to Cognitive Impairment. Cardiovasc. Toxicol. 2022, 22, 311–325. [Google Scholar] [CrossRef]
- Santos, R.A.S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/Angiotensin-(1-7)/MAS Axis of the Renin-Angiotensin System: Focus on Angiotensin-(1-7). Physiol. Rev. 2018, 98, 505–553. [Google Scholar] [CrossRef] [Green Version]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: Celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, D.; Fiselier, A.; Kovalchuk, I.; Kovalchuk, O. New AKT-dependent mechanisms of anti-COVID-19 action of high-CBD Cannabis sativa extracts. Cell Death Discov. 2022, 8, 110. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Turu, G.; Simon, A.; Gyombolai, P.; Szidonya, L.; Bagdy, G.; Lenkei, Z.; Hunyady, L. The role of diacylglycerol lipase in constitutive and angiotensin AT1 receptor-stimulated cannabinoid CB1 receptor activity. J. Biol. Chem. 2007, 282, 7753–7757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turu, G.; Várnai, P.; Gyombolai, P.; Szidonya, L.; Offertaler, L.; Bagdy, G.; Kunos, G.; Hunyady, L. Paracrine transactivation of the CB1 cannabinoid receptor by AT1 angiotensin and other Gq/11 protein-coupled receptors. J. Biol. Chem. 2009, 284, 16914–16921. [Google Scholar] [CrossRef] [Green Version]
- Rozenfeld, R.; Gupta, A.; Gagnidze, K.; Lim, M.P.; Gomes, I.; Lee-Ramos, D.; Nieto, N.; Devi, L.A. AT1R-CB₁R heteromerization reveals a new mechanism for the pathogenic properties of angiotensin II. EMBO J. 2011, 30, 2350–2363. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, M.; Nádasy, G.L.; Turu, G.; Soltész-Katona, E.; Benyó, Z.; Offermanns, S.; Ruisanchez, É.; Szabó, E.; Takáts, Z.; Bátkai, S.; et al. Endocannabinoid-mediated modulation of Gq/11 protein-coupled receptor signaling-induced vasoconstriction and hypertension. Mol. Cell. Endocrinol. 2015, 403, 46–56. [Google Scholar] [CrossRef]
- Szekeres, M.; Nádasy, G.L.; Turu, G.; Soltész-Katona, E.; Tóth, Z.E.; Balla, A.; Catt, K.J.; Hunyady, L. Angiotensin II induces vascular endocannabinoid release, which attenuates its vasoconstrictor effect via CB1 cannabinoid receptors. J. Biol. Chem. 2012, 287, 31540–31550. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, M.; Nádasy, G.L.; Soltész-Katona, E.; Hunyady, L. Control of myogenic tone and agonist induced contraction of intramural coronary resistance arterioles by cannabinoid type 1 receptors and endocanna binoids. Prostaglandins Other Lipid Mediat. 2018, 134, 77–83. [Google Scholar] [CrossRef]
- Karpińska, O.; Baranowska-Kuczko, M.; Kloza, M.; Ambrożewicz, E.; Kozłowski, T.; Kasacka, I.; Malinowska, B.; Kozłowska, H. Activation of CB1 receptors by 2-arachidonoylglycerol attenuates vasoconstriction induced by U46619 and angiotensin II in human and rat pulmonary arteries. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R883–R893. [Google Scholar] [CrossRef] [Green Version]
- Pulgar, V.M.; Yamaleyeva, L.M.; Varagic, J.; McGee, C.M.; Bader, M.; Dechend, R.; Howlett, A.C.; Brosnihan, K.B. Increased angiotensin II contraction of the uterine artery at early gestation in a transgenic model of hypertensive pregnancy is reduced by inhibition of endocannabinoid hydrolysis. Hypertension 2014, 64, 619–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiyerili, V.; Zimmer, S.; Jung, S.; Wassmann, K.; Naehle, C.P.; Lütjohann, D.; Zimmer, A.; Nickenig, G.; Wassmann, S. CB1 receptor inhibition leads to decreased vascular AT1 receptor expression, inhibition of oxidative stress and improved endothelial function. Basic Res. Cardiol. 2010, 105, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Miklós, Z.; Wafa, D.; Nádasy, G.L.; Tóth, Z.E.; Besztercei, B.; Dörnyei, G.; Laska, Z.; Benyó, Z.; Ivanics, T.; Hunyady, L.; et al. Angiotensin II-Induced Cardiac Effects Are Modulated by Endocannabinoid-Mediated CB1 Receptor Activation. Cells 2021, 10, 724. [Google Scholar] [CrossRef]
- Rajesh, M.; Bátkai, S.; Kechrid, M.; Mukhopadhyay, P.; Lee, W.S.; Horváth, B.; Holovac, E.; Cinar, R.; Liaudet, L.; Mackie, K.; et al. Cannabinoid 1 receptor promotes cardiac dysfunction, oxidative stress, inflammation, and fibrosis in diabetic cardiomyopathy. Diabetes 2012, 61, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Franco-Vadillo, A.; Toledo-Blass, M.; Rivera-Herrera, Z.; Guevara-Balcazar, G.; Orihuela-Rodriguez, O.; Morales-Carmona, J.A.; Kormanovski-Kovzova, A.; Lopez-Sanchez, P.; Rubio-Gayosso, I.; Castillo-Hernandez, M.D.C. Cannabidiol-mediated RISK PI3K/AKT and MAPK/ERK pathways decreasing reperfusion myocardial damage. Pharmacol. Res. Perspect. 2021, 9, e00784. [Google Scholar] [CrossRef]
- García-Martín, A.; Navarrete, C.; Garrido-Rodríguez, M.; Prados, M.E.; Caprioglio, D.; Appendino, G.; Muñoz, E. EHP-101 alleviates angiotensin II-induced fibrosis and inflammation in mice. Biomed. Pharmacother. 2021, 142, 112007. [Google Scholar] [CrossRef]
- Ho, W.S.; Gardiner, S.M. Acute hypertension reveals depressor and vasodilator effects of cannabinoids in conscious rats. Br. J. Pharmacol. 2009, 156, 94–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaich, C.L.; Shaltout, H.A.; Brosnihan, K.B.; Howlett, A.C.; Diz, D.I. Acute and chronic systemic CB1 cannabinoid receptor blockade improves blood pressure regulation and metabolic profile in hypertensive (mRen2)27 rats. Physiol. Rep. 2014, 2, e12108. [Google Scholar] [CrossRef] [PubMed]
- Schaich, C.L.; Grabenauer, M.; Thomas, B.F.; Shaltout, H.A.; Gallagher, P.E.; Howlett, A.C.; Diz, D.I. Medullary Endocannabinoids Contribute to the Differential Resting Baroreflex Sensitivity in Rats with Altered Brain Renin-Angiotensin System Expression. Front. Physiol. 2016, 7, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janiak, P.; Poirier, B.; Bidouard, J.P.; Cadrouvele, C.; Pierre, F.; Gouraud, L.; Barbosa, I.; Dedio, J.; Maffrand, J.P.; Le Fur, G.; et al. Blockade of cannabinoid CB1 receptors improves renal function, metabolic profile, and increased survival of obese Zucker rats. Kidney Int. 2007, 72, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Mattace Raso, G.; Simeoli, R.; Russo, R.; Santoro, A.; Pirozzi, C.; d’Emmanuele di Villa Bianca, R.; Mitidieri, E.; Paciello, O.; Pagano, T.B.; Orefice, N.S.; et al. N-Palmitoylethanolamide protects the kidney from hypertensive injury in spontaneously hypertensive rats via inhibition of oxidative stress. Pharmacol. Res. 2013, 76, 67–76. [Google Scholar] [CrossRef]
- Mattace Raso, G.; Pirozzi, C.; d’Emmanuele di Villa Bianca, R.; Simeoli, R.; Santoro, A.; Lama, A.; Di Guida, F.; Russo, R.; De Caro, C.; Sorrentino, R.; et al. Palmitoylethanolamide treatment reduces blood pressure in spontaneously hypertensive rats: Involvement of cytochrome p450-derived eicosanoids and renin angiotensin system. PLoS ONE 2015, 10, e0123602. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.Z.; Hu, C.P.; Luo, D.; Li, D.; Pan, W.; Li, S.X.; Yang, T.L.; Li, Y.J.; Zhang, G.G. Decreased anandamide transporter activity and calcitonin gene-related peptide production in spontaneously hypertensive rats: Role of angiotensin II. Eur. J. Pharmacol. 2012, 680, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Gyombolai, P.; Pap, D.; Turu, G.; Catt, K.J.; Bagdy, G.; Hunyady, L. Regulation of endocannabinoid release by G proteins: A paracrine mechanism of G protein-coupled receptor action. Mol. Cell. Endocrinol. 2012, 353, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzęda, E.; Schlicker, E.; Toczek, M.; Zalewska, I.; Baranowska-Kuczko, M.; Malinowska, B. CB1 receptor activation in the rat paraventricular nucleus induces bi-directional cardiovascular effects via modification of glutamatergic and GABAergic neurotransmission. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2017, 390, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mińczuk, K.; Schlicker, E.; Malinowska, B. Cross-Talk between CB1, AT1, AT2 and Mas Receptors Responsible for Blood Pressure Control in the Paraventricular Nucleus of Hypothalamus in Conscious Spontaneously Hypertensive Rats and Their Normotensive Controls. Cells 2022, 11, 1542. [Google Scholar] [CrossRef]
- Yokoyama, T.; Saito, T.; Ohbuchi, T.; Hashimoto, H.; Suzuki, H.; Otsubo, H.; Fujihara, H.; Nagatomo, T.; Ueta, Y. TRPV1 gene deficiency attenuates miniature EPSC potentiation induced by mannitol and angiotensin II in supraoptic magnocellular neurons. J. Neurosci. 2010, 30, 876–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyires, K.; Rónai, A.Z.; Zádori, Z.S.; Tóth, V.E.; Németh, J.; Szekeres, M.; Hunyady, L. Angiotensin II-induced activation of central AT1 receptors exerts endocannabinoid-mediated gastroprotective effect in rats. Mol. Cell. Endocrinol. 2014, 382, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Shigemura, N.; Iwata, S.; Yasumatsu, K.; Ohkuri, T.; Horio, N.; Sanematsu, K.; Yoshida, R.; Margolskee, R.F.; Ninomiya, Y. Angiotensin II modulates salty and sweet taste sensitivities. J. Neurosci. 2013, 33, 6267–6277. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Heterologous regulation of the cannabinoid type 1 receptor by angiotensin II in astrocytes of spontaneously hypertensive rats. J. Neurochem. 2016, 139, 523–536. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. MAPK activation patterns of AT1R and CB1R in SHR versus Wistar astrocytes: Evidence of CB1R hypofunction and crosstalk between AT1R and CB1R. Cell Signal. 2017, 40, 81–90. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Contrasting Roles of Ang II and ACEA in the Regulation of IL10 and IL1β Gene Expression in Primary SHR Astroglial Cultures. Molecules 2021, 26, 3012. [Google Scholar] [CrossRef]
- Wang, L.; Shi, F.X.; Xu, W.Q.; Cao, Y.; Li, N.; Li, M.; Wang, Q.; Wang, J.Z.; Tian, Q.; Yu, L.K.; et al. The Down-Expression of ACE and IDE Exacerbates Exogenous Amyloid-β Neurotoxicity in CB2R-/- Mice. J. Alzheimers. Dis. 2018, 64, 957–971. [Google Scholar] [CrossRef]
- Jourdan, T.; Szanda, G.; Rosenberg, A.Z.; Tam, J.; Earley, B.J.; Godlewski, G.; Cinar, R.; Liu, Z.; Liu, J.; Ju, C.; et al. Overactive cannabinoid 1 receptor in podocytes drives type 2 diabetic nephropathy. Proc. Natl. Acad. Sci. USA 2014, 111, E5420–E5428. [Google Scholar] [CrossRef] [Green Version]
- Barutta, F.; Bellini, S.; Mastrocola, R.; Gambino, R.; Piscitelli, F.; di Marzo, V.; Corbetta, B.; Vemuri, V.K.; Makriyannis, A.; Annaratone, L.; et al. Reversal of albuminuria by combined AM6545 and perindopril therapy in experimental diabetic nephropathy. Br. J. Pharmacol. 2018, 175, 4371–4385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckenstaler, R.; Sandori, J.; Gekle, M.; Benndorf, R.A. Angiotensin II receptor type 1—An update on structure, expression and pathology. Biochem. Pharmacol. 2021, 192, 114673. [Google Scholar] [CrossRef] [PubMed]
- Karpińska, O.; Baranowska-Kuczko, M.; Kloza, M.; Kozłowska, H. Endocannabinoids modulate Gq/11 protein-coupled receptor agonist-induced vasoconstriction via a negative feedback mechanism. J. Pharm. Pharmacol. 2018, 70, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, K.; Whalen, E.J.; Violin, J.D.; Stiber, J.A.; Rosenberg, P.B.; Premont, R.T.; Coffman, T.M.; Rockman, H.A.; Lefkowitz, R.J. Beta-arrestin2-mediated inotropic effects of the angiotensin II type 1A receptor in isolated cardiac myocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 16284–16289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jama, H.A.; Muralitharan, R.R.; Xu, C.; O’Donnell, J.A.; Bertagnolli, M.; Broughton, B.R.S.; Head, G.A.; Marques, F.Z. Rodent models of hypertension. Br. J. Pharmacol. 2022, 179, 918–937. [Google Scholar] [CrossRef]
- Vargas Vargas, R.A.; Varela Millán, J.M.; Fajardo Bonilla, E. Renin-angiotensin system: Basic and clinical aspects-A general perspective. Endocrinol. Diabetes Nutr. 2022, 69, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, B.; Baranowska-Kuczko, M.; Schlicker, E. Triphasic blood pressure responses to cannabinoids: Do we understand the mechanism? Br. J. Pharmacol. 2012, 165, 2073–2088. [Google Scholar] [CrossRef] [Green Version]
- Malinowska, B.; Toczek, M.; Pędzińska-Betiuk, A.; Schlicker, E. Cannabinoids in arterial, pulmonary and portal hypertension-mechanisms of action and potential therapeutic significance. Br. J. Pharmacol. 2019, 176, 1395–1411. [Google Scholar] [CrossRef]
- Goncharuk, V.D. The hypothalamus and its role in hypertension. Handb. Clin. Neurol. 2021, 182, 333–354. [Google Scholar] [CrossRef]
- Ma, H.; Chen, S.R.; Chen, H.; Pan, H.L. Endogenous AT1 receptor-protein kinase C activity in the hypothalamus augments glutamatergic input and sympathetic outflow in hypertension. J. Physiol. 2019, 597, 4325–4340. [Google Scholar] [CrossRef]
- Elsaafien, K.; de Kloet, A.D.; Krause, E.G.; Sumners, C. Brain Angiotensin Type-1 and Type-2 Receptors in Physiological and Hypertensive Conditions: Focus on Neuroinflammation. Curr. Hypertens. Rep. 2020, 22, 48. [Google Scholar] [CrossRef]
- Fatima, N.; Patel, S.N.; Hussain, T. Angiotensin II Type 2 Receptor: A Target for Protection Against Hypertension, Metabolic Dysfunction, and Organ Remodeling. Hypertension 2021, 77, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Dupont, A.G.; Légat, L. GABA is a mediator of brain AT1 and AT2 receptor-mediated blood pressure responses. Hypertens. Res. 2020, 43, 995–1005. [Google Scholar] [CrossRef]
- Soya, A.; Serino, R.; Fujihara, H.; Onaka, T.; Ozaki, Y.; Saito, T.; Nakamura, J.; Ueta, Y. Cannabinoids modulate synaptic activity in the rat supraoptic nucleus. J. Neuroendocrinol. 2005, 17, 609–615. [Google Scholar] [CrossRef]
- Yoshida, R.; Ohkuri, T.; Jyotaki, M.; Yasuo, T.; Horio, N.; Yasumatsu, K.; Sanematsu, K.; Shigemura, N.; Yamamoto, T.; Margolskee, R.F.; et al. Endocannabinoids selectively enhance sweet taste. Proc. Natl. Acad. Sci. USA 2010, 107, 935–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.L.; Schelb, V.; Trendelenburg, A.U.; Starke, K. Enhancement of noradrenaline release by angiotensin II and bradykinin in mouse atria: Evidence for cross-talk between G(q/11) protein- and G(i/o) protein-coupled receptors. Br. J. Pharmacol. 2000, 129, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Trendelenburg, A.U.; Meyer, A.; Klebroff, W.; Guimarães, S.; Starke, K. Crosstalk between presynaptic angiotensin receptors, bradykinin receptors and alpha 2-autoreceptors in sympathetic neurons: A study in alpha 2-adrenoceptor-deficient mice. Br. J. Pharmacol. 2003, 138, 1389–1402. [Google Scholar] [CrossRef] [Green Version]
- Clayton, P.; Subah, S.; Venkatesh, R.; Hill, M.; Bogoda, N. Palmitoylethanolamide: A Potential Alternative to Cannabidiol. J. Diet. Suppl. 2021, 28, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Kicman, A.; Toczek, M. The Effects of Cannabidiol, a Non-Intoxicating Compound of Cannabis, on the Cardiovascular System in Health and Disease. Int. J. Mol. Sci. 2020, 21, 6740. [Google Scholar] [CrossRef]
- Fang, S.; Livergood, M.C.; Nakagawa, P.; Wu, J.; Sigmund, C.D. Role of the Peroxisome Proliferator Activated Receptors in Hypertension. Circ. Res. 2021, 128, 1021–1039. [Google Scholar] [CrossRef]
- Khundmiri, S.J.; Ecelbarger, C.M.; Amponsem, J.; Ji, H.; Sandberg, K.; Lee, D.L. PPAR-α knockout leads to elevated blood pressure response to angiotensin II infusion associated with an increase in renal α-1 Na+/K+ ATPase protein expression and activity. Life Sci. 2022, 296, 120444. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Molecular Basis of the Brain Renin Angiotensin System in Cardiovascular and Neurologic Disorders: Uncovering a Key Role for the Astroglial Angiotensin Type 1 Receptor AT1R. J. Pharmacol. Exp. Ther. 2018, 366, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinar, R.; Iyer, M.R.; Kunos, G. The therapeutic potential of second and third generation CB1R antagonists. Pharmacol. Ther. 2020, 208, 107477. [Google Scholar] [CrossRef]
- Deeba, F.; Kumar, A.; Mukherjee, M.; Sharma, A.K.; Sharma, M. Targeting the endocannabinoid system in diabesity: Fact or fiction? Drug Discov. Today 2021, 26, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.; François, H. Cannabinoid Receptor 1 Inhibition in Chronic Kidney Disease: A New Therapeutic Toolbox. Front. Endocrinol. 2021, 12, 720734. [Google Scholar] [CrossRef]
- Jorgačević, B.; Vučević, D.; Samardžić, J.; Mladenović, D.; Vesković, M.; Vukićević, D.; Ješić, R.; Radosavljević, T. The Effect of CB1 Antagonism on Hepatic Oxidative/Nitrosative Stress and Inflammation in Nonalcoholic Fatty Liver Disease. Curr. Med. Chem. 2021, 28, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Cinar, R.; Iyer, M.R.; Kunos, G. Dual inhibition of CB1 receptors and iNOS, as a potential novel approach to the pharmacological management of acute and long COVID-19. Br. J. Pharmacol. 2022, 179, 2121–2127. [Google Scholar] [CrossRef]
- Quarta, C.; Cota, D. Anti-obesity therapy with peripheral CB1 blockers: From promise to safe(?) practice. Int. J. Obes. 2020, 44, 2179–2193. [Google Scholar] [CrossRef]
- Brosnihan, K.B.; Pulgar, V.M.; Gallagher, P.E.; Neves, L.A.; Yamaleyeva, L.M. Local uterine Ang-(1-7) infusion augments the expression of cannabinoid receptors and differentially alters endocannabinoid metabolizing enzymes in the decidualized uterus of pseudopregnant rats. Reprod. Biol. Endocrinol. 2015, 13, 5. [Google Scholar] [CrossRef] [Green Version]
- Gurm, H.; Hirota, J.A.; Raha, S. Cannabinoid Signalling in Immune-Reproductive Crosstalk during Human Pregnancy. Biomedicines 2021, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Grzeda, E.; Schlicker, E.; Łuczaj, W.; Harasim, E.; Baranowska-Kuczko, M.; Malinowska, B. Bi-directional CB1 receptor-mediated cardiovascular effects of cannabinoids in anaesthetized rats: Role of the paraventricular nucleus. J. Physiol. Pharmacol. 2015, 66, 343–353. [Google Scholar]
- Reindl-Schwaighofer, R.; Hödlmoser, S.; Eskandary, F.; Poglitsch, M.; Bonderman, D.; Strassl, R.; Aberle, J.H.; Oberbauer, R.; Zoufaly, A.; Hecking, M. ACE2 Elevation in Severe COVID-19. Am. J. Respir. Crit. Care Med. 2021, 203, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Fagyas, M.; Fejes, Z.; Sütő, R.; Nagy, Z.; Székely, B.; Pócsi, M.; Ivády, G.; Bíró, E.; Bekő, G.; Nagy, A.; et al. Circulating ACE2 activity predicts mortality and disease severity in hospitalized COVID-19 patients. Int. J. Infect. Dis. 2022, 115, 8–16. [Google Scholar] [CrossRef]
- Esposito, G.; Pesce, M.; Seguella, L.; Sanseverino, W.; Lu, J.; Corpetti, C.; Sarnelli, G. The potential of cannabidiol in the COVID-19 pandemic. Br. J. Pharmacol. 2020, 177, 4967–4970. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Sharma, C.; Goyal, S.N.; Kumar, S.; Ojha, S. CB2 receptor-selective agonists as candidates for targeting infection, inflammation, and immunity in SARS-CoV-2 infections. Drug Dev. Res. 2021, 82, 7–11. [Google Scholar] [CrossRef]
- Aragão, L.G.H.S.; Oliveira, J.T.; Temerozo, J.R.; Mendes, M.A.; Salerno, J.A.; Pedrosa, C.S.G.; Puig-Pijuan, T.; Veríssimo, C.P.; Ornelas, I.M.; Torquato, T.; et al. WIN 55,212-2 shows anti-inflammatory and survival properties in human iPSC-derived cardiomyocytes infected with SARS-CoV-2. PeerJ 2021, 9, e12262. [Google Scholar] [CrossRef]
- National Institutes of Health. COVID-19 Treatment Guidelines Panel. Coronavirus Disease 2019 (COVID-19) Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov (accessed on 2 May 2022).
- Pitakbut, T.; Nguyen, G.N.; Kayser, O. Activity of THC, CBD, and CBN on Human ACE2 and SARS-CoV1/2 Main Protease to Understand Antiviral Defense Mechanism. Planta Med. 2021. [Google Scholar] [CrossRef]
- Sarkar, I.; Sen, G.; Bhattacharya, M.; Bhattacharyya, S.; Sen, A. In silico inquest reveals the efficacy of Cannabis in the treatment of post-Covid-19 related neurodegeneration. J. Biomol. Struct. Dyn. 2021, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I.; Kovalchuk, O. In search of preventive strategies: Novel high-CBD Cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging 2020, 12, 22425–22444. [Google Scholar] [CrossRef]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11, 1462. [Google Scholar] [CrossRef]
- Corpetti, C.; Del Re, A.; Seguella, L.; Palenca, I.; Rurgo, S.; De Conno, B.; Pesce, M.; Sarnelli, G.; Esposito, G. Cannabidiol inhibits SARS-Cov-2 spike (S) protein-induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase-1 signaling suppression in Caco-2 cell line. Phytother. Res. 2021, 35, 6893–6903. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’Antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation. Antibiotics 2021, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Muchiri, R.N.; Bates, T.A.; Weinstein, J.B.; Leier, H.C.; Farley, S.; Tafesse, F.G. Cannabinoids Block Cellular Entry of SARS-CoV-2 and the Emerging Variants. J. Nat. Prod. 2022, 85, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Ferber, S.G.; Namdar, D.; Hen-Shoval, D.; Eger, G.; Koltai, H.; Shoval, G.; Shbiro, L.; Weller, A. The “Entourage Effect”: Terpenes Coupled with Cannabinoids for the Treatment of Mood Disorders and Anxiety Disorders. Curr. Neuropharmacol. 2020, 18, 87–96. [Google Scholar] [CrossRef]
- Finlay, D.B.; Sircombe, K.J.; Nimick, M.; Jones, C.; Glass, M. Terpenoids From Cannabis Do Not Mediate an Entourage Effect by Acting at Cannabinoid Receptors. Front. Pharmacol. 2020, 11, 359. [Google Scholar] [CrossRef]
- Millar, S.A.; Maguire, R.F.; Yates, A.S.; O’Sullivan, S.E. Towards Better Delivery of Cannabidiol (CBD). Pharmaceuticals 2020, 13, 219. [Google Scholar] [CrossRef]
- Alsherbiny, M.A.; Li, C.G. Medicinal Cannabis-Potential Drug Interactions. Medicines 2018, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Doohan, P.T.; Oldfield, L.D.; Arnold, J.C.; Anderson, L.L. Cannabinoid Interactions with Cytochrome P450 Drug Metabolism: A Full-Spectrum Characterization. AAPS J. 2021, 23, 91. [Google Scholar] [CrossRef] [PubMed]
- Brow, J.D.; Winterstein, A.G. Potential Adverse Drug Events and Drug-Drug Interactions with Medical and Consumer Cannabidiol (CBD) Use. J. Clin. Med. 2019, 8, 989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocis, P.T.; Vrana, K.E. Delta-9-Tetrahydrocannabinol and Cannabidiol Drug-Drug Interactions. Med. Cannabis Cannabinoids 2020, 3, 61–73. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mińczuk, K.; Baranowska-Kuczko, M.; Krzyżewska, A.; Schlicker, E.; Malinowska, B. Cross-Talk between the (Endo)Cannabinoid and Renin-Angiotensin Systems: Basic Evidence and Potential Therapeutic Significance. Int. J. Mol. Sci. 2022, 23, 6350. https://doi.org/10.3390/ijms23116350
Mińczuk K, Baranowska-Kuczko M, Krzyżewska A, Schlicker E, Malinowska B. Cross-Talk between the (Endo)Cannabinoid and Renin-Angiotensin Systems: Basic Evidence and Potential Therapeutic Significance. International Journal of Molecular Sciences. 2022; 23(11):6350. https://doi.org/10.3390/ijms23116350
Chicago/Turabian StyleMińczuk, Krzysztof, Marta Baranowska-Kuczko, Anna Krzyżewska, Eberhard Schlicker, and Barbara Malinowska. 2022. "Cross-Talk between the (Endo)Cannabinoid and Renin-Angiotensin Systems: Basic Evidence and Potential Therapeutic Significance" International Journal of Molecular Sciences 23, no. 11: 6350. https://doi.org/10.3390/ijms23116350