
The BCC7 Protein Contributes to the Toxoplasma Basal Pole by Interfacing between the MyoC Motor and the IMC Membrane Network

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
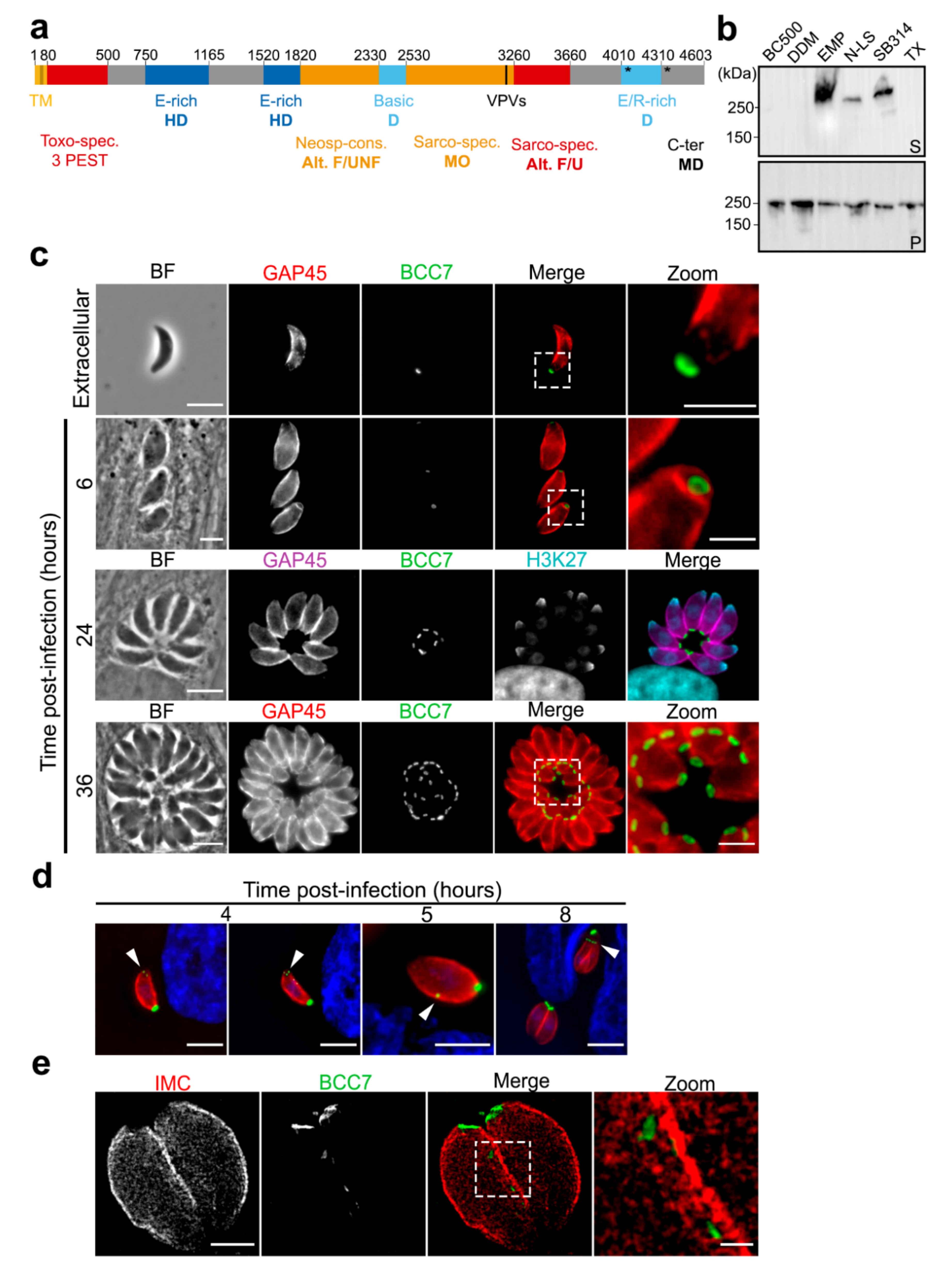
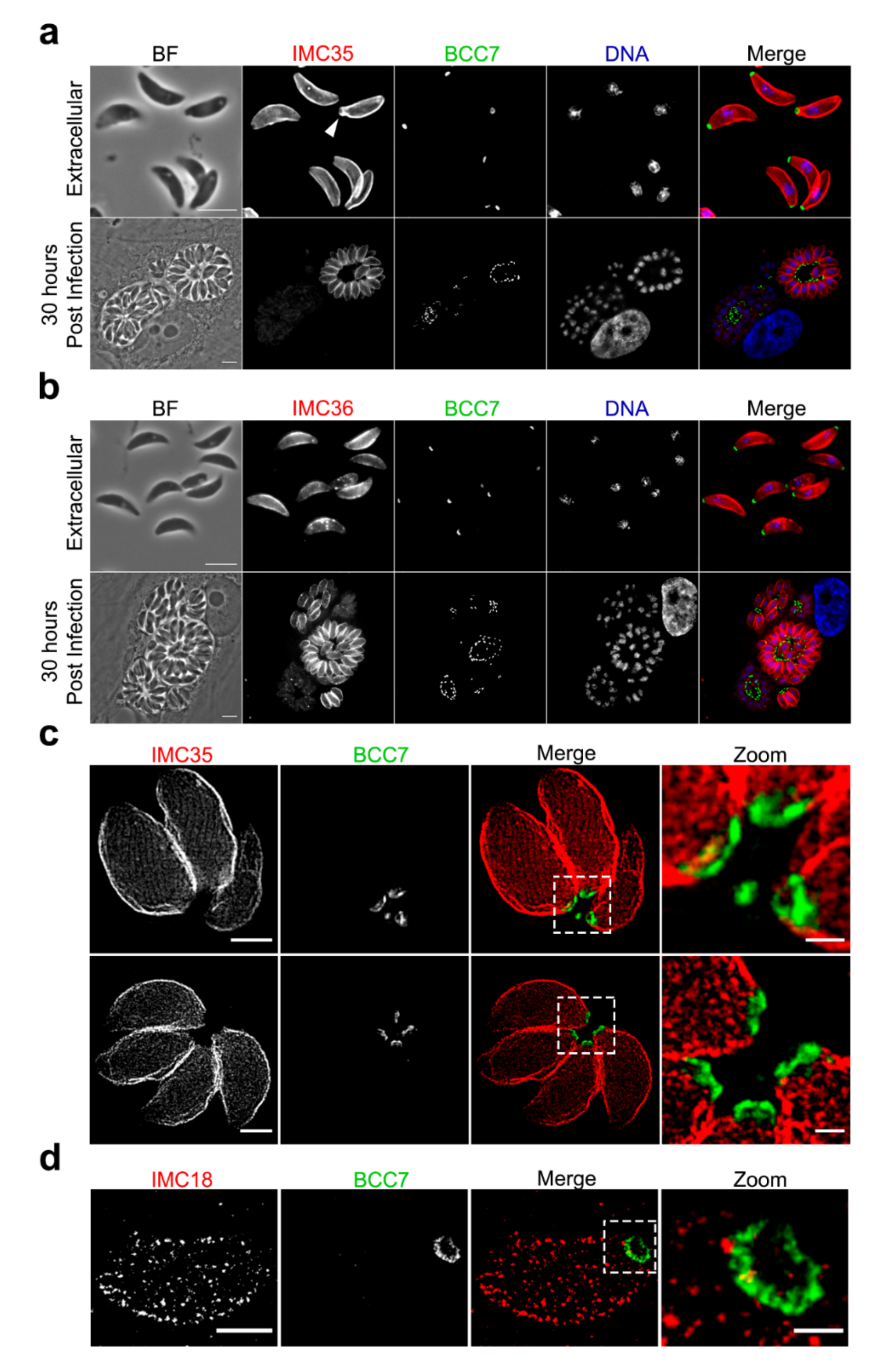
2.1. BCC7, a Coccidia Restricted Protein with a Ring-Shaped Architecture Marking the Basal Pole of the T. gondii Tachyzoite
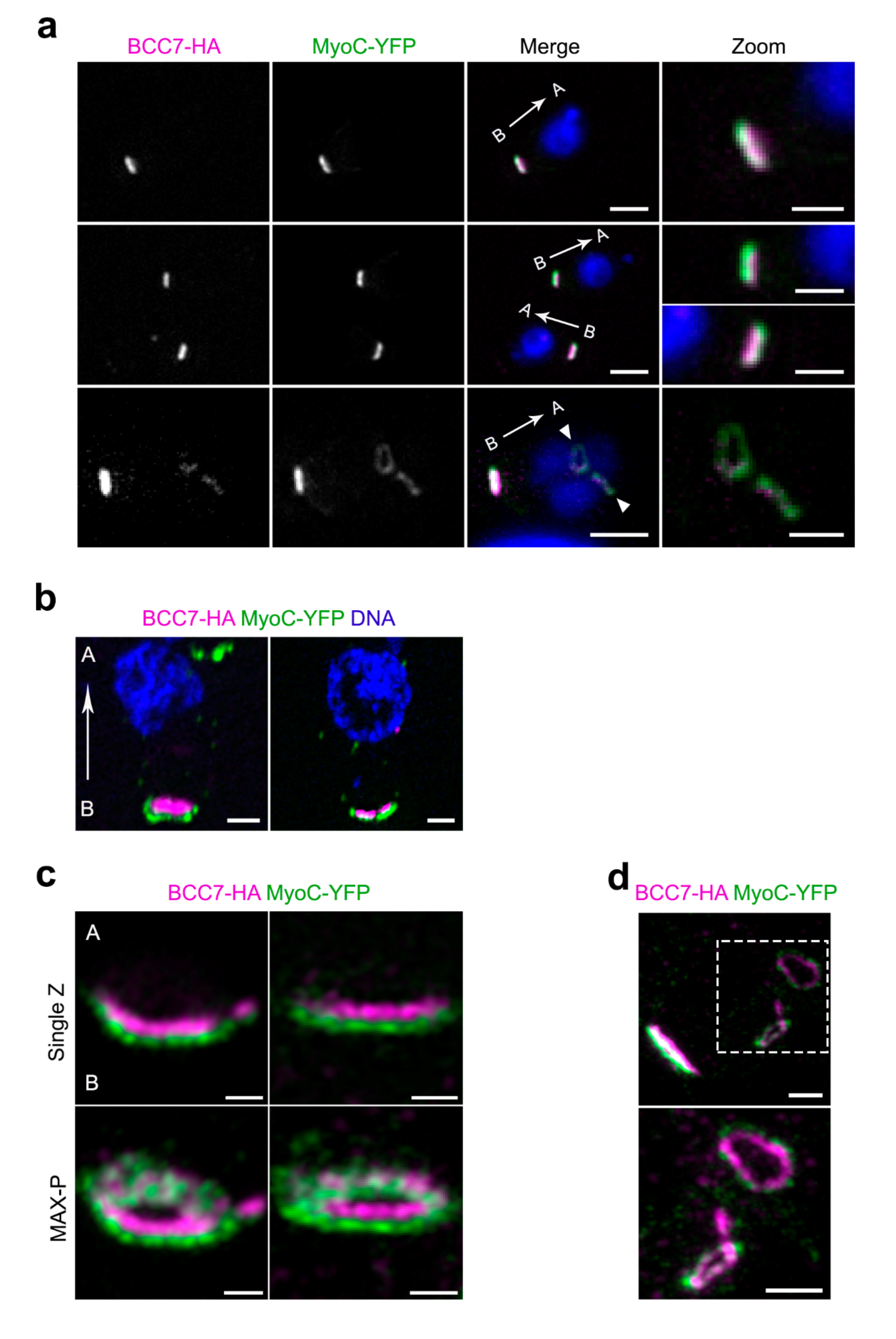
2.2. Super Resolution Microscopy Highlights the Close Proximity between MyoC and BCC7
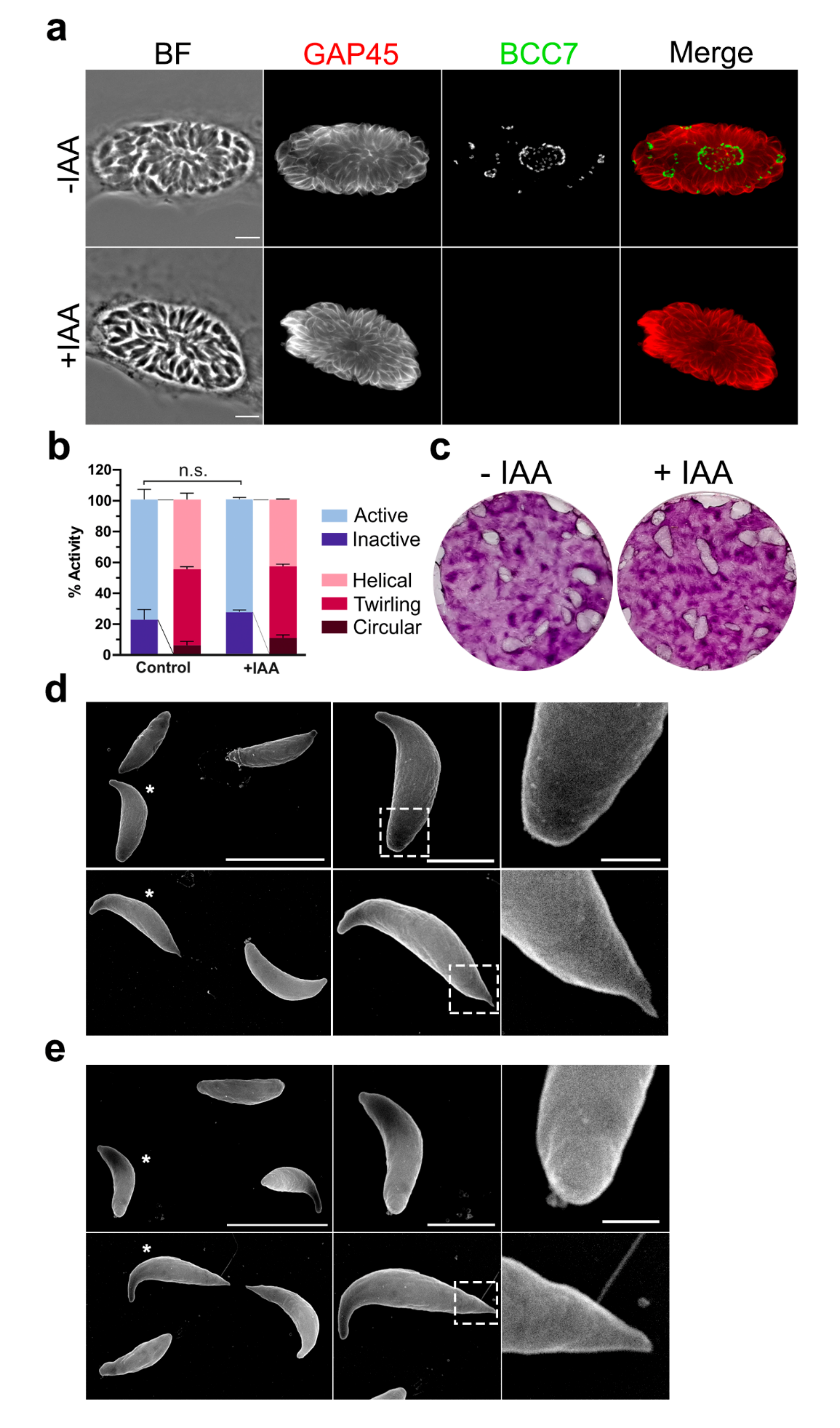
2.3. The BCC7 Protein Is Dispensable for the Tachyzoite Extracellular and Intracellular Lifestyle
2.4. Identification of Putative BCC7-Interacting Partners by Co-Immunoprecipitation Coupled to Mass Spectrometry Analysis
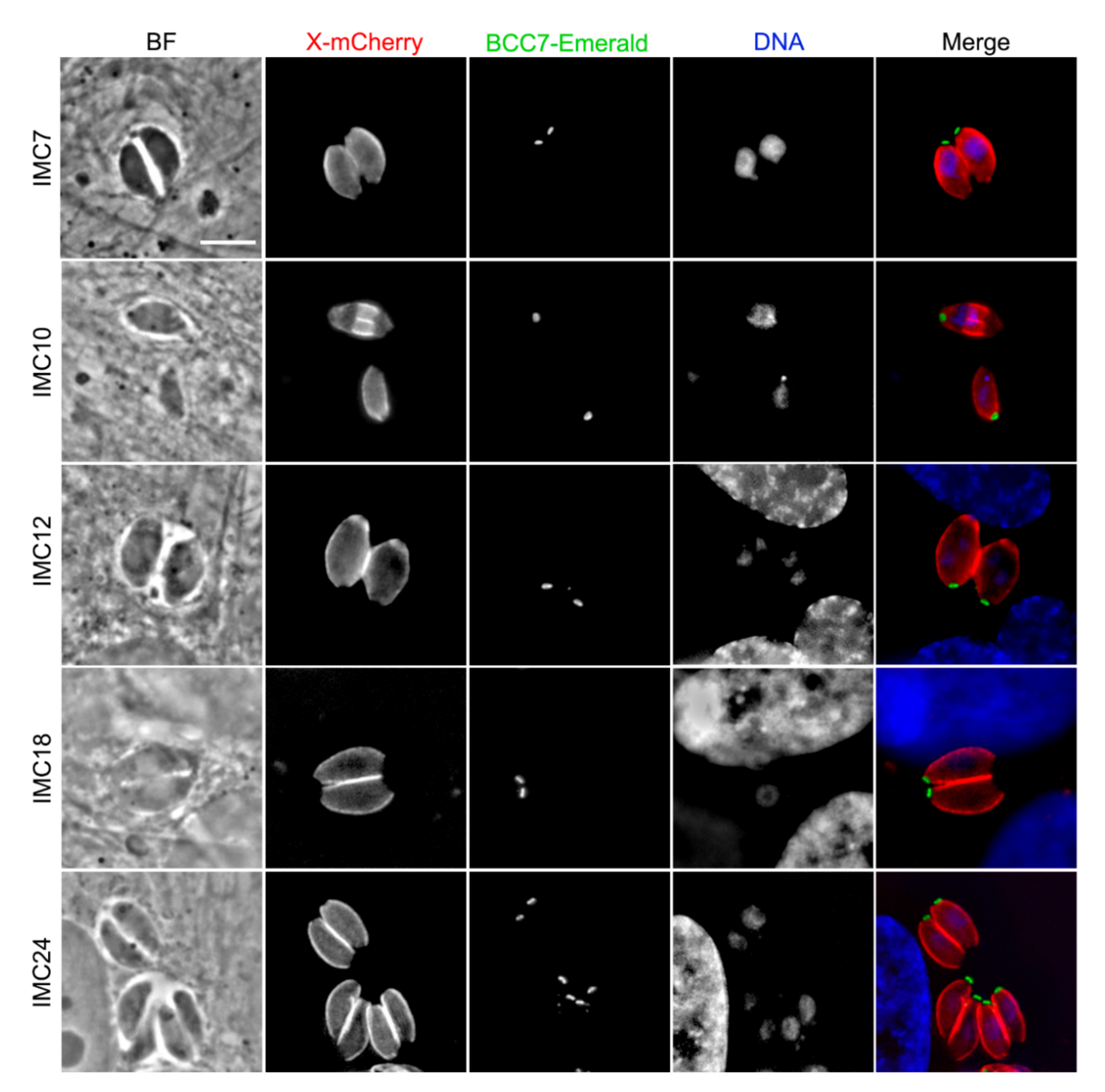
2.5. A Majority of IMC Proteins, Including Two New Ones Were Identified in the BCC7-Binding Protein Screen
3. Discussion
4. Materials and Methods
4.1. Parasites and Host Cells
4.2. Generation of Transgenic Parasites Expressing Endogenously Tagged Proteins
4.3. Bioinformatic Analyses
4.4. Auxin-Inducible Degron Plasmid Construction
4.5. Plaque Assays
4.6. BCC7 Solubilization Assays
4.7. Immunoprecipitation Procedure
4.8. Western Blots
4.9. Mass Spectrometry-Based Quantitative Proteomic Analyses
4.10. Immunofluorescence Microscopy
4.11. Video Microscopy
4.12. Tachyzoite Egress and Motility Assays
4.13. Expansion Microscopy
4.14. Scanning Electron Microscopy (SEM)
4.15. Image Processing
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carme, B.; Demar, M.; Ajzenberg, D.; Dardé, M.L. Severe Acquired Toxoplasmosis Caused by Wild Cycle of Toxoplasma gondii, French Guiana. Emerg. Infect. Dis. 2009, 15, 656–658. [Google Scholar] [CrossRef] [PubMed]
- Anderson-White, B.; Beck, J.R.; Chen, C.-T.; Meissner, M.; Bradley, P.J.; Gubbels, M.-J. Cytoskeleton Assembly in Toxoplasma gondii Cell Division. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 298, pp. 1–31. ISBN 978-0-12-394309-5. [Google Scholar]
- Mann, T.; Gaskins, E.; Beckers, C. Proteolytic Processing of TgIMC1 during Maturation of the Membrane Skeleton of Toxoplasma gondii. J. Biol. Chem. 2002, 277, 41240–41246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissette, N.S.; Sibley, L.D. Cytoskeleton of Apicomplexan Parasites. Microbiol. Mol. Biol. Rev. 2002, 66, 21–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlou, G.; Touquet, B.; Vigetti, L.; Renesto, P.; Bougdour, A.; Debarre, D.; Balland, M.; Tardieux, I. Coupling Polar Adhesion with Traction, Spring, and Torque Forces Allows High-Speed Helical Migration of the Protozoan Parasite Toxoplasma. ACS Nano 2020, 14, 7121–7139. [Google Scholar] [CrossRef]
- Beck, J.R.; Rodriguez-Fernandez, I.A.; Cruz de Leon, J.; Huynh, M.-H.; Carruthers, V.B.; Morrissette, N.S.; Bradley, P.J. A Novel Family of Toxoplasma IMC Proteins Displays a Hierarchical Organization and Functions in Coordinating Parasite Division. PLoS Pathog. 2010, 6, e1001094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentini, G.; Kong-Hap, M.; El Hajj, H.; Francia, M.; Claudet, C.; Striepen, B.; Dubremetz, J.-F.; Lebrun, M. Identification and Characterization of Toxoplasma SIP, a Conserved Apicomplexan Cytoskeleton Protein Involved in Maintaining the Shape, Motility and Virulence of the Parasite: Cytoskeleton Protein Associated with the IMC Edges. Cell. Microbiol. 2015, 17, 62–78. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.R.; Gow, M.; Kang, J.H.; Shortt, E.; Manalis, S.R.; Meissner, M.; Lourido, S. Alveolar Proteins Stabilize Cortical Microtubules in Toxoplasma gondii. Nat. Commun. 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Johnson, J.; Florens, L.; Fraunholz, M.; Suravajjala, S.; DiLullo, C.; Yates, J.; Roos, D.S.; Murray, J.M. Cytoskeletal Components of an Invasion Machine—The Apical Complex of Toxoplasma gondii. PLoS Pathog. 2006, 2, e13. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Pacheco, N.; Tosetti, N.; Koreny, L.; Waller, R.F.; Soldati-Favre, D. Evolution, Composition, Assembly, and Function of the Conoid in Apicomplexa. Trends Parasitol. 2020, 36, 688–704. [Google Scholar] [CrossRef]
- Hu, K. Organizational Changes of the Daughter Basal Complex during the Parasite Replication of Toxoplasma gondii. PLoS Pathog. 2008, 4, e10. [Google Scholar] [CrossRef] [Green Version]
- Gubbels, M.-J.; Ferguson, D.J.P.; Saha, S.; Romano, J.D.; Chavan, S.; Primo, V.A.; Michaud, C.; Coppens, I.; Engelberg, K. Toxoplasma gondii’s Basal Complex: The Other Apicomplexan Business End Is Multifunctional. Front. Cell. Infect. Microbiol. 2022, 12, 882166. [Google Scholar] [CrossRef]
- Engelberg, K.; Bechtel, T.; Michaud, C.; Weerapana, E.; Gubbels, M.-J. A More Complex Basal Complex: Novel Components Mapped to the Toxoplasma gondii Cytokinesis Machinery Portray an Expanded Hierarchy of Its Assembly and Function. bioRxiv 2021. [Google Scholar] [CrossRef]
- Frénal, K.; Krishnan, A.; Soldati-Favre, D. The Actomyosin Systems in Apicomplexa. In Myosins; Coluccio, L.M., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2020; Volume 1239, pp. 331–354. ISBN 978-3-030-38061-8. [Google Scholar]
- Frénal, K.; Jacot, D.; Hammoudi, P.-M.; Graindorge, A.; Maco, B.; Soldati-Favre, D. Myosin-Dependent Cell-Cell Communication Controls Synchronicity of Division in Acute and Chronic Stages of Toxoplasma gondii. Nat. Commun. 2017, 8, 15710. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, G.; Biesaga, M.; Touquet, B.; Lagal, V.; Balland, M.; Dufour, A.; Hakimi, M.; Tardieux, I. Toxoplasma Parasite Twisting Motion Mechanically Induces Host Cell Membrane Fission to Complete Invasion within a Protective Vacuole. Cell Host Microbe 2018, 24, 81–96.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez de León, C.T.; Díaz Martín, R.D.; Mendoza Hernández, G.; González Pozos, S.; Ambrosio, J.R.; Mondragón Flores, R. Proteomic Characterization of the Subpellicular Cytoskeleton of Toxoplasma Gondii Tachyzoites. J. Proteom. 2014, 111, 86–99. [Google Scholar] [CrossRef]
- Engelberg, K.; Ivey, F.D.; Lin, A.; Kono, M.; Lorestani, A.; Faugno-Fusci, D.; Gilberger, T.-W.; White, M.; Gubbels, M.-J. A MORN1-Associated HAD Phosphatase in the Basal Complex Is Essential for Toxoplasma Gondii Daughter Budding. Cell. Microbiol. 2016, 18, 1153–1171. [Google Scholar] [CrossRef] [Green Version]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The Eukaryotic Pathogen, Vector and Host Bioinformatics Resource Center. Nucleic Acids Res. 2022, 50, D898–D911. [Google Scholar] [CrossRef]
- Barylyuk, K.; Koreny, L.; Ke, H.; Butterworth, S.; Crook, O.M.; Lassadi, I.; Gupta, V.; Tromer, E.; Mourier, T.; Stevens, T.J.; et al. A Subcellular Atlas of Toxoplasma Reveals the Functional Context of the Proteome. Cell Biol. 2020, 28, 752–766.e9. [Google Scholar]
- Gould, S.B.; Tham, W.-H.; Cowman, A.F.; McFadden, G.I.; Waller, R.F. Alveolins, a New Family of Cortical Proteins That Define the Protist Infrakingdom Alveolata. Mol. Biol. Evol. 2008, 25, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Gubbels, M.-J.; Wieffer, M.; Striepen, B. Fluorescent Protein Tagging in Toxoplasma gondii: Identification of a Novel Inner Membrane Complex Component Conserved among Apicomplexa. Mol. Biochem. Parasitol. 2004, 137, 99–110. [Google Scholar] [CrossRef]
- Mann, T. Characterization of the Subpellicular Network, a Filamentous Membrane Skeletal Component in the Parasite Toxoplasma gondii. Mol. Biochem. Parasitol. 2001, 115, 257–268. [Google Scholar] [CrossRef]
- Heaslip, A.T.; Leung, J.M.; Carey, K.L.; Catti, F.; Warshaw, D.M.; Westwood, N.J.; Ballif, B.A.; Ward, G.E. A Small-Molecule Inhibitor of T. Gondii Motility Induces the Posttranslational Modification of Myosin Light Chain-1 and Inhibits Myosin Motor Activity. PLoS Pathog. 2010, 6, e1000720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassie, A.T.; Zhao, Y.; Boyden, E.S. Expansion Microscopy: Principles and Uses in Biological Research. Nat. Methods 2019, 16, 33–41. [Google Scholar] [CrossRef]
- Brown, K.; Long, S.; Sibley, L. Conditional Knockdown of Proteins Using Auxin-Inducible Degron (AID) Fusions in Toxoplasma gondii. Bio-Protocol 2018, 8, e2728. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.-H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-Wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowthert, L.A.; Ku, N.O.; Liao, J.; Coulombe, P.A.; Omary, M.B. Empigen BB: A Useful Detergent for Solubilization and Biochemical Analysis of Keratins. Biochem. Biophys. Res. Commun. 1995, 206, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Hettmann, C.; Herm, A.; Geiter, A.; Frank, B.; Schwarz, E.; Soldati, T.; Soldati, D. A Dibasic Motif in the Tail of a Class XIV Apicomplexan Myosin Is an Essential Determinant of Plasma Membrane Localization. Mol. Biol. Cell 2000, 11, 1385–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Döşkaya, M.; Liang, L.; Jain, A.; Can, H.; Gülçe İz, S.; Felgner, P.L.; Değirmenci Döşkaya, A.; Davies, D.H.; Gürüz, A.Y. Discovery of New Toxoplasma gondii Antigenic Proteins Using a High Throughput Protein Microarray Approach Screening Sera of Murine Model Infected Orally with Oocysts and Tissue Cysts. Parasit. Vectors 2018, 11, 393. [Google Scholar] [CrossRef]
- Besteiro, S.; Michelin, A.; Poncet, J.; Dubremetz, J.-F.; Lebrun, M. Export of a Toxoplasma gondii Rhoptry Neck Protein Complex at the Host Cell Membrane to Form the Moving Junction during Invasion. PLoS Pathog. 2009, 5, e1000309. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.L.; Kim, E.W.; Toh, J.Y.; Vashisht, A.A.; Rashoff, A.Q.; Van, C.; Huang, A.S.; Moon, A.S.; Bell, H.N.; Bentolila, L.A.; et al. Novel Components of the Toxoplasma Inner Membrane Complex Revealed by BioID. mBio 2015, 6, e02357-14. [Google Scholar] [CrossRef] [Green Version]
- Semenovskaya, K.; Lévêque, M.F.; Berry, L.; Bordat, Y.; Dubremetz, J.; Lebrun, M.; Besteiro, S. TgZFP2 Is a Novel Zinc Finger Protein Involved in Coordinating Mitosis and Budding in Toxoplasma. Cell. Microbiol. 2020, 22, e13120. [Google Scholar] [CrossRef] [PubMed]
- Anderson-White, B.R.; Ivey, F.D.; Cheng, K.; Szatanek, T.; Lorestani, A.; Beckers, C.J.; Ferguson, D.J.P.; Sahoo, N.; Gubbels, M.-J. A Family of Intermediate Filament-like Proteins Is Sequentially Assembled into the Cytoskeleton of Toxoplasma gondii: IMC Proteins in Toxoplasma Cell Division. Cell. Microbiol. 2011, 13, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Pacheco, N.; Tosetti, N.; Krishnan, A.; Haase, R.; Maco, B.; Suarez, C.; Ren, B.; Soldati-Favre, D. Revisiting the Role of Toxoplasma Gondii ERK7 in the Maintenance and Stability of the Apical Complex. mBio 2021, 12, e02057-21. [Google Scholar] [CrossRef]
- Behnke, M.S.; Wootton, J.C.; Lehmann, M.M.; Radke, J.B.; Lucas, O.; Nawas, J.; Sibley, L.D.; White, M.W. Coordinated Progression through Two Subtranscriptomes Underlies the Tachyzoite Cycle of Toxoplasma gondii. PLoS ONE 2010, 5, e12354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Striepen, B.; Jordan, C.N.; Reiff, S.; van Dooren, G.G. Building the Perfect Parasite: Cell Division in Apicomplexa. PLoS Pathog. 2007, 3, e78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouologuem, D.T.; Roos, D.S. Dynamics of the Toxoplasma gondii Inner Membrane Complex. J. Cell Sci. 2014, 127, 3320–3330. [Google Scholar] [CrossRef] [Green Version]
- Gubbels, M.-J.; Vaishnava, S.; Boot, N.; Dubremetz, J.-F.; Striepen, B. A MORN-Repeat Protein Is a Dynamic Component of the Toxoplasma gondii Cell Division Apparatus. J. Cell Sci. 2006, 119, 2236–2245. [Google Scholar] [CrossRef] [Green Version]
- Uversky, V.N. The Alphabet of Intrinsic Disorder: II. Various Roles of Glutamic Acid in Ordered and Intrinsically Disordered Proteins. Intrinsically Disord. Proteins 2013, 1, e24684. [Google Scholar] [CrossRef] [Green Version]
- McBride, A.E.; Conboy, A.K.; Brown, S.P.; Ariyachet, C.; Rutledge, K.L. Specific Sequences within Arginine-Glycine-Rich Domains Affect MRNA-Binding Protein Function. Nucleic Acids Res. 2009, 37, 4322–4330. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.B.; Kraft, L.G.K.; van Dooren, G.G.; Goodman, C.D.; Ford, K.L.; Cassin, A.M.; Bacic, A.; McFadden, G.I.; Waller, R.F. Ciliate Pellicular Proteome Identifies Novel Protein Families with Characteristic Repeat Motifs That Are Common to Alveolates. Mol. Biol. Evol. 2011, 28, 1319–1331. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Xu, M.; Pan, S.; Wang, H.; Peng, H.; Liu, Z. ITRAQ-Based Phosphoproteomic Analysis of Toxoplasma gondii Tachyzoites Provides Insight Into the Role of Phosphorylation for Its Invasion and Egress. Front. Cell. Infect. Microbiol. 2020, 10, 586466. [Google Scholar] [CrossRef] [PubMed]
- Silmon de Monerri, N.C.; Yakubu, R.R.; Chen, A.L.; Bradley, P.J.; Nieves, E.; Weiss, L.M.; Kim, K. The Ubiquitin Proteome of Toxoplasma gondii Reveals Roles for Protein Ubiquitination in Cell-Cycle Transitions. Cell Host Microbe 2015, 18, 621–633. [Google Scholar] [CrossRef] [Green Version]
- Huynh, M.-H.; Carruthers, V.B. Tagging of Endogenous Genes in a Toxoplasma gondii Strain Lacking Ku80. Eukaryot. Cell 2009, 8, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Services. Available online: https://services.healthtech.dtu.dk (accessed on 8 April 2022).
- EMBOSS: Epestfind. Available online: https://emboss.bioinformatics.nl/cgi-bin/emboss/epestfind/ (accessed on 8 April 2022).
- ExPASy-ProtParam Tool. Available online: https://web.expasy.org/protparam/ (accessed on 8 April 2022).
- FoldIndex(c). Available online: https://fold.proteopedia.org/cgi-bin/findex (accessed on 8 April 2022).
- Treeck, M.; Sanders, J.L.; Elias, J.E.; Boothroyd, J.C. The Phosphoproteomes of Plasmodium Falciparum and Toxoplasma gondii Reveal Unusual Adaptations within and beyond the Parasites’ Boundaries. Cell Host Microbe 2011, 10, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.M.; Long, S.; Sibley, L.D. Plasma Membrane Association by N-Acylation Governs PKG Function in Toxoplasma gondii. mBio 2017, 8, e00375-17. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Hackett, C.G.; Tran, F.; Westwood, N.J.; Lourido, S. Efficient Genome Engineering of Toxoplasma gondii Using CRISPR/Cas9. PLoS ONE 2014, 9, e100450. [Google Scholar] [CrossRef] [Green Version]
- Casabona, M.G.; Vandenbrouck, Y.; Attree, I.; Couté, Y. Proteomic Characterization of Pseudomonas Aeruginosa PAO1 Inner Membrane. Proteomics 2013, 13, 2419–2423. [Google Scholar] [CrossRef]
- Bouyssié, D.; Hesse, A.-M.; Mouton-Barbosa, E.; Rompais, M.; Macron, C.; Carapito, C.; Gonzalez de Peredo, A.; Couté, Y.; Dupierris, V.; Burel, A.; et al. Proline: An Efficient and User-Friendly Software Suite for Large-Scale Proteomics. Bioinformatics 2020, 36, 3148–3155. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, S.; Combes, F.; Lazar, C.; Giai Gianetto, Q.; Gatto, L.; Dorffer, A.; Hesse, A.-M.; Couté, Y.; Ferro, M.; Bruley, C.; et al. DAPAR & ProStaR: Software to Perform Statistical Analyses in Quantitative Discovery Proteomics. Bioinformatics 2017, 33, 135–136. [Google Scholar] [CrossRef] [Green Version]
- Asano, S.M.; Gao, R.; Wassie, A.T.; Tillberg, P.W.; Chen, F.; Boyden, E.S. Expansion Microscopy: Protocols for Imaging Proteins and RNA in Cells and Tissues. Curr. Protoc. Cell Biol. 2018, 80, e56. [Google Scholar] [CrossRef]
- Molecular Devices. Available online: https://www.moleculardevices.com (accessed on 8 April 2022).
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- de Chaumont, F.; Dallongeville, S.; Chenouard, N.; Hervé, N.; Pop, S.; Provoost, T.; Meas-Yedid, V.; Pankajakshan, P.; Lecomte, T.; Le Montagner, Y.; et al. Icy: An Open Bioimage Informatics Platform for Extended Reproducible Research. Nat. Methods 2012, 9, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, A.; Beichel, R.; Kalpathy-Cramer, J.; Finet, J.; Fillion-Robin, J.-C.; Pujol, S.; Bauer, C.; Jennings, D.; Fennessy, F.; Sonka, M.; et al. 3D Slicer as an Image Computing Platform for the Quantitative Imaging Network. Magn. Reson. Imaging 2012, 30, 1323–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Partner | TGGT1 Accession Number | Description | MW (kDa) | Proteomic Data | Subcellular Location | Phenotype Score | |
|---|---|---|---|---|---|---|---|
| Log2FC | p-Value | ||||||
| - | 311230 | BCC7 | 494.8 | 8.71 | 2.68 × 10−7 | unknown | 0.74 |
| 1 | 222220 | IMC7 | 46.8 | 3.44 | 1.96 × 10−3 | IMC | −0.64 |
| 2 | 258470 | IMC24 [32] | 31.7 | 3.26 | 1.10 × 10−3 | IMC | 2.12 |
| 3 | 230940 | hypothetical protein | 155.2 | 2.95 | 7.63 × 10−3 | unknown | −4.92 |
| 4 | 233450 | SAG-related sequence SRS29A | 44.2 | 2.71 | 4.61 × 10−4 | PM-peripheral 1 | 2.42 |
| 5 | 248700 | IMC12 | 29.8 | 2.38 | 6.88 × 10−3 | IMC | −0.17 |
| 6 | 212260 | TgZFP2 [33] | 38 | 2.47 | 4.11 × 10−2 | unknown | −4.5 |
| 7 | 216670 | FUSE-binding protein 2 | 100 | 2.19 | 4.86 × 10−2 | nucleus-non chromatin | −3.31 |
| 8 | 230160 | hypothetical protein | 15.8 | 2.19 | 1.54 × 10−2 | IMC | 2.11 |
| 9 | 236950 | hypothetical protein | 12.5 | 2.13 | 1.07 × 10−2 | IMC | 0.96 |
| 10 | 230210 | IMC10 | 61.2 | 2.05 | 1.19 × 10−2 | IMC | −4.7 |
| 11 | 295360 | IMC18 [32] | 29.6 | 2.00 | 1.45 × 10−2 | IMC | 1.13 |
| 12 | 315610 | hypothetical protein | 15.9 | 2.67 | 8.17 × 10−4 | unknown | 1.33 |
| 13 | 231160 | hypothetical protein | 18.3 | 2.41 | 3.11 × 10−3 | IMC | −1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigetti, L.; Labouré, T.; Roumégous, C.; Cannella, D.; Touquet, B.; Mayer, C.; Couté, Y.; Frénal, K.; Tardieux, I.; Renesto, P. The BCC7 Protein Contributes to the Toxoplasma Basal Pole by Interfacing between the MyoC Motor and the IMC Membrane Network. Int. J. Mol. Sci. 2022, 23, 5995. https://doi.org/10.3390/ijms23115995
Vigetti L, Labouré T, Roumégous C, Cannella D, Touquet B, Mayer C, Couté Y, Frénal K, Tardieux I, Renesto P. The BCC7 Protein Contributes to the Toxoplasma Basal Pole by Interfacing between the MyoC Motor and the IMC Membrane Network. International Journal of Molecular Sciences. 2022; 23(11):5995. https://doi.org/10.3390/ijms23115995
Chicago/Turabian StyleVigetti, Luis, Tatiana Labouré, Chloé Roumégous, Dominique Cannella, Bastien Touquet, Claudine Mayer, Yohann Couté, Karine Frénal, Isabelle Tardieux, and Patricia Renesto. 2022. "The BCC7 Protein Contributes to the Toxoplasma Basal Pole by Interfacing between the MyoC Motor and the IMC Membrane Network" International Journal of Molecular Sciences 23, no. 11: 5995. https://doi.org/10.3390/ijms23115995