
Oxidative Stress in Malaria: Potential Benefits of Antioxidant Therapy


 ,
, 
{kind=link}
{kind=link}
Abstract
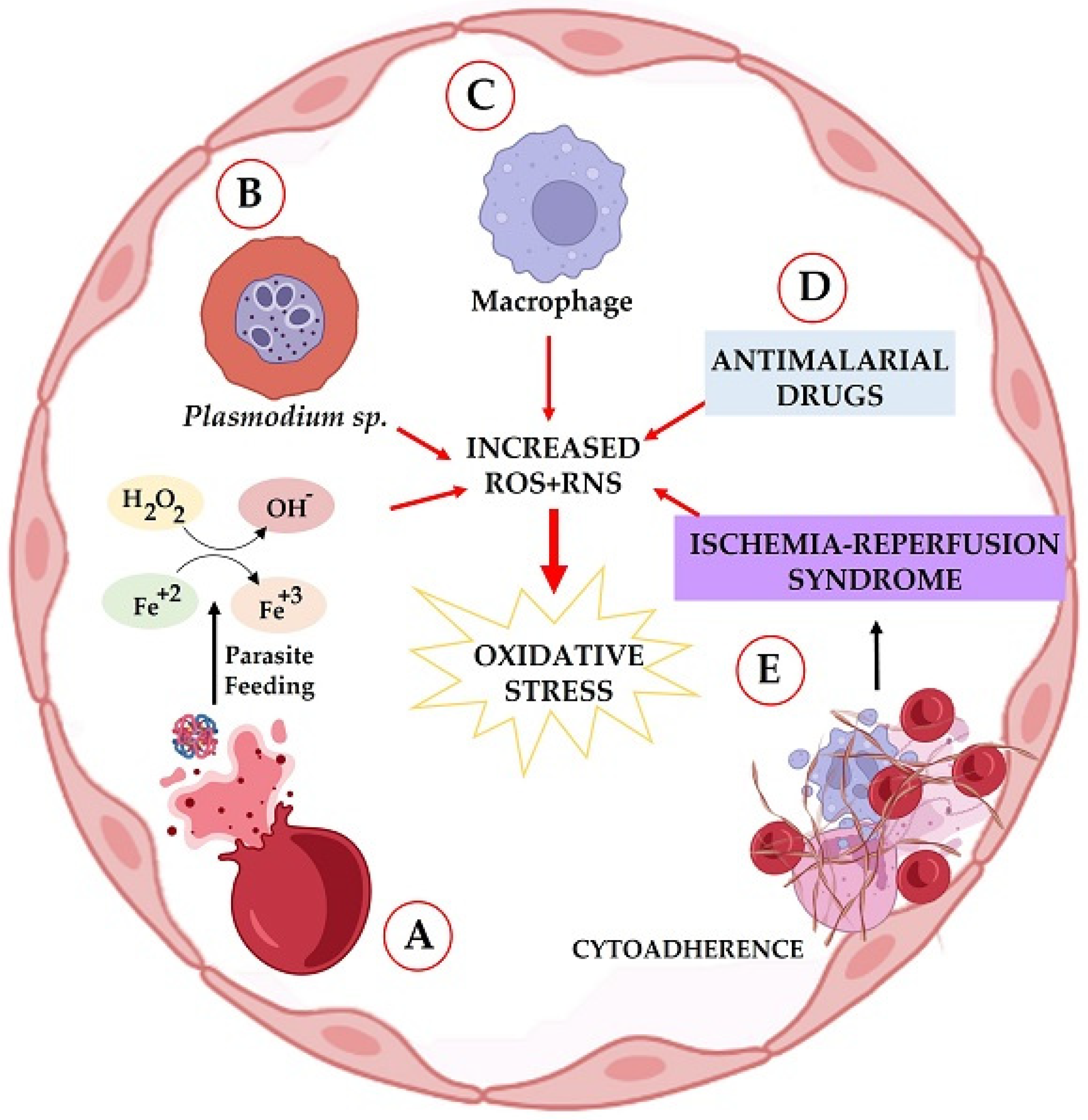
:1. Introduction
2. Oxidative Stress in the Host, Induced by Plasmodium
2.1. The Role of Pro-Oxidants and NO in Malaria
2.2. The Host’s Antioxidant Defense Mechanism against Plasmodium
3. Antioxidant Therapy in Malaria
3.1. Antioxidant Vitamin Supplementation in Malaria
3.2. Antimalarial Therapy Combined with Antioxidant Supplementation
4. Final Remarks
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. WHO: World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2022; Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (accessed on 1 January 2020).
- Organización Panamericana de la Salud. PAHO: Actualización Epidemiológica: Malaria en las Américas en el Contexto de la Pandemia de COVID-19; PAHO: Washington, DC, USA, 2020; Available online: https://iris.paho.org/handle/10665.2/52288 (accessed on 6 October 2021).
- Ashley, E.A.; Phyo, A.P.; Woodrow, C.J. Malaria. Lancet 2018, 391, 1608–1621. [Google Scholar] [CrossRef]
- Walther, B.; Walther, M. What does it take to control malaria? Ann. Trop. Med. Parasitol. 2007, 101, 657–672. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.M.; Costa, M.R.F.; Filho, F.S.S.; Vieira, J.L.F.; Nascimento, M.T.S.; Brasil, L.W.; Nogueira, F.; Silveira, H.; Reyes-Lecca, R.C.; Monteiro, W.M.; et al. Plasmodium vivax chloroquine resistance and anemia in the western Brazilian Amazon. Antimicrob. Agents Chemother. 2014, 58, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzila, A. The past, present and future of antifolates in the treatment of Plasmodium falciparum infection. J. Antimicrob. Chemother. 2006, 57, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, G.; Musset, L.; Hubert, V.; Le Bras, J.; Clain, J. Emergence of resistance to atovaquone-proguanil in malaria parasites: Insights from computational modeling and clinical case reports. Antimicrob. Agents Chemother. 2014, 58, 4504–4514. [Google Scholar] [CrossRef] [Green Version]
- Wongsrichanalai, C.; Pickard, A.L.; Wernsdorfer, W.H.; Meshnick, S.R. Epidemiology of drug-resistant malaria. Lancet Infect. Dis. 2002, 2, 209–218. [Google Scholar] [CrossRef]
- Noedl, H.; Se, Y.; Schaecher, K.; Smith, B.L.; Socheat, D.; Fukuda, M.M. Evidence of artemisinin-resistant malaria in western Cambodia. N. Engl. J. Med. 2008, 359, 2619–2620. [Google Scholar] [CrossRef]
- Van Hong, N.; Amambua-Ngwa, A.; Tuan, N.Q.; Cuong, D.D.; Giang, N.T.H.; Van Dung, N.; Erhart, A. Severe malaria not responsive to artemisinin derivatives in man returning from Angola to Vietnam. Emerg. Infect. Dis. 2014, 20, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Percário, S.; Moreira, D.R.; Gomes, B.A.; Ferreira, M.E.; Gonçalves, A.C.; Laurindo, P.S.; Green, M.D. Oxidative stress in malaria. Int. J. Mol. Sci. 2012, 13, 16346–16372. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.A.; Comim, C.M.; Hermani, F.; Silva, B.; Barichello, T.; Portella, A.C.; Castro-Faria-Neto, H.C. Cognitive dysfunction is sustained after rescue therapy in experimental cerebral malaria and is reduced by additive antioxidant therapy. PLoS Pathog. 2010, 6, e1000963. [Google Scholar] [CrossRef]
- Ezzi, A.A.A.; Salahy, M.B.A.; Shnawa, B.H.; Abed, G.H.; Mandour, A.M. Changes in levels of antioxidant markers and status of some enzyme activities among falciparum malaria patients in Yemen. J. Microbiol. Exp. 2017, 4, 00131. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.A.Q.; da Silva, L.F.; Gomes, A.R.Q.; Moreira, D.R.; Dolabela, M.F.; Santos, R.S.; Percário, S. N-acetyl cysteine and mushroom Agaricus sylvaticus supplementation decreased parasitaemia and pulmonary oxidative stress in a mice model of malaria. Malar. J. 2015, 14, 202. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, H.; Singh, S.K.; Chauhan, B.S.; Gunjan, S.; Tripathi, R. Potential cerebral malaria therapy: Intramuscular arteether and vitamin D co-administration. Parasitology 2016, 143, 1557. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Ekeh, F.N.; Ekechukwu, N.E.; Chukwuma, C.F.; Aguzie, I.O.N.; Ohanu, C.M.; Ebido, C.; Oluah, S.N. Mixed vitamin C and zinc diet supplements co-administered with artemether drug improved haematological profile and survival of mice infected with Plasmodium berghei. Food Sci. Hum. Wellness 2019, 8, 275–282. [Google Scholar] [CrossRef]
- Owusu-Agyei, S.; Newton, S.; Mahama, E.; Febir, L.G.; Ali, M.; Adjei, K.; Tanumihardjo, S.A. Impact of vitamin A with zinc supplementation on malaria morbidity in Ghana. Nutr. J. 2013, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Olofin, I.O.; Spiegelman, D.; Aboud, S.; Duggan, C.; Danaei, G.; Fawzi, W.W. Supplementation with multivitamins and vitamin A and incidence of malaria among HIV-infected Tanzanian women. J. Acquir. Immune Defic. Syndr. 2014, 67, S173–S178. [Google Scholar] [CrossRef] [Green Version]
- Yakoob, M.Y.; Qadir, M. Vitamin A supplementation for prevention and treatment of malaria during pregnancy and childhood: A systematic review and meta-analysis. J. Epidemiol. Glob. Health 2018, 8, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Du, Y.T.; Liu, J.H.; Li, Y.; Zheng, L.; Qin, X.S.; Cao, Y.M. Modulation of anti-malaria immunity by vitamin A in C57BL/6J mice infected with heterogenic Plasmodium. Int. Immunopharmacol. 2019, 76, 105882. [Google Scholar] [CrossRef]
- Ganiyu, K.A.; Akinleye, M.O.; Fola, T. A study of the effect of ascorbic acid on the antiplasmodial activity of artemether in Plasmodium berghei infected mice. J. Appl. Pharm. Sci. 2012, 2. Available online: https://www.japsonline.com/abstract.php?article_id=510&sts=2 (accessed on 16 October 2021).
- Qin, X.; Liu, J.; Du, Y.; Li, Y.; Zheng, L.; Chen, G.; Cao, Y. Different doses of vitamin C supplementation enhances the Th1 immune response to early Plasmodium yoelii 17XL infection in BALB/c mice. Int. Immunopharmacol. 2019, 70, 387–395. [Google Scholar] [CrossRef]
- He, X.; Yan, J.; Zhu, X.; Wang, Q.; Pang, W.; Qi, Z.; Cao, Y. Vitamin D inhibits the occurrence of experimental cerebral malaria in mice by suppressing the host inflammatory response. J. Immunol. 2014, 193, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Du, Y.; Feng, Y.; Wang, Q.; Pang, W.; Qi, Z.; Cao, Y. Oral administration of vitamin D and importance in prevention of cerebral malaria. Int. Immunopharmacol. 2018, 64, 356–363. [Google Scholar] [CrossRef]
- Suzuki, H.; Kume, A.; Herbas, M.S. Potential of vitamin E deficiency, induced by inhibition of α-tocopherol efflux, in murine malaria infection. Int. J. Mol. Sci. 2019, 20, 64. [Google Scholar] [CrossRef] [Green Version]
- Isah, M.B.; Ibrahim, M.A. The role of antioxidants treatment on the pathogenesis of malarial infections: A review. Parasitol. Res. 2014, 113, 801–809. [Google Scholar] [CrossRef]
- Zeba, A.N.; Sorgho, H.; Rouamba, N.; Zongo, I.; Rouamba, J.; Guiguemdé, R.T.; Ouedraogo, J.B. Major reduction of malaria morbidity with combined vitamin A and zinc supplementation in young children in Burkina Faso: A randomized double blind trial. Nutr. J. 2008, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.M.; Mugusi, F.M.; Etheredge, A.J.; Gunaratna, N.S.; Abioye, A.I.; Aboud, S.; Fawzi, W.W. Vitamin A and zinc supplementation among pregnant women to prevent placental malaria: A randomized, double-blind, placebo-controlled trial in Tanzania. Am. J. Trop. Med. Hyg. 2017, 96, 826–834. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Hill, K.E.; Li, P.; Xu, J.; Zhou, D.; Motley, A.K.; Burk, R.F. Optimization of selenoprotein P and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: A placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient Chinese subjects. Am. J. Clin. Nutr. 2010, 92, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Adebayo, A.H.; Olasehinde, G.I.; Egbeola, O.A.; Yakubu, O.F.; Adeyemi, A.O.; Adekeye, B.T. Enhanced antioxidant capacity following selenium supplemented antimalarial therapy in Plasmodium berghei infected mice. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2018; Volume 1954, p. 030019. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, T.; Li, Q.; Li, D. Prevention of Keshan disease by selenium supplementation: A systematic review and meta-analysis. Biol. Trace Elem. Res. 2018, 186, 98–105. [Google Scholar] [CrossRef]
- Sarr, D.; Cooper, C.A.; Bracken, T.C.; Martinez-Uribe, O.; Nagy, T.; Moore, J.M. Oxidative stress: A potential therapeutic target in placental malaria. ImmunoHorizons 2017, 1, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Atiku, S.M.; Louise, N.; Kasozi, D.M. Severe oxidative stress in sickle cell disease patients with uncomplicated Plasmodium falciparum malaria in Kampala, Uganda. BMC Infect. Dis. 2019, 19, 600. [Google Scholar] [CrossRef] [PubMed]
- Götz, A.; Ty, M.C.; Rodriguez, A. Oxidative stress enhances dendritic cell responses to Plasmodium falciparum. ImmunoHorizons 2019, 3, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velagapudi, R.; Kosoko, A.M.; Olajide, O.A. Induction of neuroinflammation and neurotoxicity by synthetic hemozoin. Cell. Mol. Neurobiol. 2019, 39, 1187–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narsaria, N.; Mohanty, C.; Das, B.K.; Mishra, S.P.; Prasad, R. Oxidative stress in children with severe malaria. J. Trop. Pediatr. 2012, 58, 147–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nsiah, K.; Bahaah, B.; Oppong Afranie, B.; Koffie, S.; Akowuah, E.; Donkor, S. Oxidative stress and hemoglobin level of complicated and uncomplicated malaria cases among children: A cross-sectional study in Kumasi metropolis, Ghana. J. Trop. Med. 2019, 2019, 8479076. [Google Scholar] [CrossRef] [Green Version]
- Bartoloni, A.; Zammarchi, L. Clinical aspects of uncomplicated and severe malaria. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012026. [Google Scholar] [CrossRef] [Green Version]
- Anidi, I.U.; Servinsky, L.E.; Rentsendorj, O.; Stephens, R.S.; Scott, A.L.; Pearse, D.B. CD36 and Fyn kinase mediate malaria-induced lung endothelial barrier dysfunction in mice infected with Plasmodium berghei. PLoS ONE 2013, 8, e71010. [Google Scholar] [CrossRef] [Green Version]
- Imai, T.; Iwawaki, T.; Akai, R.; Suzue, K.; Hirai, M.; Taniguchi, T.; Hisaeda, H. Evaluating experimental cerebral malaria using oxidative stress indicator OKD48 mice. Int. J. Parasitol. 2014, 44, 681–685. [Google Scholar] [CrossRef]
- Linares, M.; Marín-García, P.; Pérez-Benavente, S.; Sánchez-Nogueiro, J.; Puyet, A.; Bautista, J.M.; Diez, A. Brain-derived neurotrophic factor and the course of experimental cerebral malaria. Brain Res. 2013, 1490, 210–224. [Google Scholar] [CrossRef]
- Janka, J.J.; Koita, O.A.; Traoré, B.; Traoré, J.M.; Mzayek, F.; Sachdev, V.; Krogstad, D.J. Increased pulmonary pressures and myocardial wall stress in children with severe malaria. J. Infect. Dis. 2010, 202, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Na-Ek, P.; Punsawad, C. Expression of 4-hydroxynonenal (4-HNE) and heme oxygenase-1 (HO-1) in the kidneys of Plasmodium berghei-infected mice. J. Trop. Med. 2020, 2020, 8813654. [Google Scholar] [CrossRef]
- Aitken, E.H.; Alemu, A.; Rogerson, S.J. Neutrophils and malaria. Front. Immunol. 2008, 9, 3005. [Google Scholar] [CrossRef] [Green Version]
- Cunnington, A.J.; Njie, M.; Correa, S.; Takem, E.N.; Riley, E.M.; Walther, M. Prolonged neutrophil dysfunction after Plasmodium falciparum malaria is related to hemolysis and heme oxygenase-1 induction. J. Immunol. 2012, 189, 5336–5346. [Google Scholar] [CrossRef] [Green Version]
- Feintuch, C.M.; Saidi, A.; Seydel, K.; Chen, G.; Goldman-Yassen, A.; Mita-Mendoza, N.K.; Daily, J.P. Activated neutrophils are associated with pediatric cerebral malaria vasculopathy in Malawian children. MBio 2016, 7, e01300-15. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; de Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef] [Green Version]
- Giusti, P.; Urban, B.C.; Frascaroli, G.; Albrecht, L.; Tinti, A.; Troye-Blomberg, M.; Varani, S. Plasmodium falciparum-infected erythrocytes and β-hematin induce partial maturation of human dendritic cells and increase their migratory ability in response to lymphoid chemokines. Infect. Immun. 2011, 79, 2727–2736. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Ludlow, L.E.; Hasang, W.; Rogerson, S.J.; Jaworowski, A. Opsonization of malaria-infected erythrocytes activates the inflammasome and enhances inflammatory cytokine secretion by human macrophages. Malar. J. 2012, 11, 343. [Google Scholar] [CrossRef] [Green Version]
- Götz, A.; San Tang, M.; Ty, M.C.; Arama, C.; Ongoiba, A.; Doumtabe, D.; Rodriguez, A. Atypical activation of dendritic cells by Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2017, 114, E10568–E10577. [Google Scholar] [CrossRef] [Green Version]
- Ourives, S.S.; Borges, Q.I.; Dos Santos, D.S.A.; Melo, E.C.M.; de Souza, R.M.; Damazo, A.S. Analysis of the lymphocyte cell population during malaria caused by Plasmodium vivax and its correlation with parasitaemia and thrombocytopaenia. Malar. J. 2018, 17, 303. [Google Scholar] [CrossRef] [Green Version]
- Ozarslan, N.; Robinson, J.F.; Gaw, S.L. Circulating monocytes, tissue macrophages, and malaria. J. Trop. Med. 2019, 2019, 3720838. [Google Scholar] [CrossRef] [Green Version]
- Klinkhamhom, A.; Glaharn, S.; Srisook, C.; Ampawong, S.; Krudsood, S.; Ward, S.A.; Viriyavejakul, P. M1 macrophage features in severe Plasmodium falciparum malaria patients with pulmonary oedema. Malar. J. 2020, 19, 182. [Google Scholar] [CrossRef]
- Borges, T.K.S.; Alves, É.A.; Vasconcelos, H.A.; Carneiro, F.P.; Nicola, A.M.; Magalhães, K.G.; Muniz-Junqueira, M.I. Differences in the modulation of reactive species, lipid bodies, cyclooxygenase-2, 5-lipoxygenase and PPAR-γ in cerebral malaria-susceptible and resistant mice. Immunobiology 2017, 222, 604–619. [Google Scholar] [CrossRef]
- Eisenbarth, S.C. Dendritic cell subsets in T cell programming: Location dictates function. Nat. Rev. Immunol. 2019, 19, 89–103. [Google Scholar] [CrossRef]
- Ty, M.C.; Zuniga, M.; Götz, A.; Kayal, S.; Sahu, P.K.; Mohanty, A. Malaria inflammation by xanthine oxidase-produced reactive oxygen species. EMBO Mol. Med. 2019, 11, e9903. [Google Scholar] [CrossRef]
- Burda, P.C.; Crosskey, T.; Lauk, K.; Zurborg, A.; Söhnchen, C.; Liffner, B.; Gilberger, T.W. Structure-based identification and functional characterization of a lipocalin in the malaria parasite Plasmodium falciparum. Cell Rep. 2020, 31, 107817. [Google Scholar] [CrossRef]
- Wunderlich, J.; Rohrbach, P.; Dalton, J.P. The malaria digestive vacuole. Front. Biosci. 2012, 4, 1424–1448. [Google Scholar] [CrossRef] [Green Version]
- Sohail, M.; Kaul, A.; Raziuddin, M.; Adak, T. Decreased glutathione-S-transferase activity: Diagnostic and protective role in vivax malaria. Clin. Biochem. 2007, 40, 377–382. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Fattman, C.L.; Tan, R.J.; Oury, T.D. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am. J. Respir. Crit. Care Med. 2005, 172, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Jortzik, E.; Becker, K. Thioredoxin and glutathione systems in Plasmodium falciparum. Int. J. Med. Microbiol. 2012, 302, 187–194. [Google Scholar] [CrossRef]
- Lim, J.; Gowda, D.C.; Krishnegowda, G.; Luckhart, S. Induction of nitric oxide synthase in Anopheles stephensi by Plasmodium falciparum: Mechanism of signaling and the role of parasite glycosylphosphatidylinositols. Infect. Immun. 2005, 73, 2778–2789. [Google Scholar] [CrossRef] [Green Version]
- Orengo, J.M.; Leliwa-Sytek, A.; Evans, J.E.; Evans, B.; van de Hoef, D.; Nyako, M.; Rodriguez, A. Uric acid is a mediator of the Plasmodium falciparum-induced inflammatory response. PLoS ONE 2009, 4, e5194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mita-Mendoza, N.K.; van de Hoef, D.L.; Lopera-Mesa, T.M.; Doumbia, S.; Konate, D.; Doumbouya, M.; Fairhurst, R.M. A potential role for plasma uric acid in the endothelial pathology of Plasmodium falciparum malaria. PLoS ONE 2013, 8, e54481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, S.M.; Mitchell, A.J.; Cowden, W.B.; Sanni, L.A.; Dinauer, M.; Haan, J.B.; Hunt, N.H. Phagocyte-derived reactive oxygen species do not influence the progression of murine blood-stage malaria infections. Infect. Immun. 2005, 73, 4941–4947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grellier, P.; Marozienė, A.; Nivinskas, H.; Šarlauskas, J.; Aliverti, A.; Čėnas, N. Antiplasmodial activity of quinones: Roles of aziridinyl substituents and the inhibition of Plasmodium falciparum glutathione reductase. Arch. Biochem. Biophys. 2010, 494, 32–39. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, H.; Gerhard, G.S. Heme synthesis increases artemisinin-induced radical formation and cytotoxicity that can be suppressed by superoxide scavengers. Chem.-Biol. Interact. 2010, 186, 30–35. [Google Scholar] [CrossRef]
- Müller, I.B.; Hyde, J.E. Antimalarial drugs: Modes of action and mechanisms of parasite resistance. Future Microbiol. 2010, 5, 1857–1873. [Google Scholar] [CrossRef]
- Achan, J.; Talisuna, A.O.; Erhart, A.; Yeka, A.; Tibenderana, J.K.; Baliraine, F.N.; D’Alessandro, U. Quinine, an old anti-malarial drug in a modern world: Role in the treatment of malaria. Malar. J. 2011, 10, 144. [Google Scholar] [CrossRef] [Green Version]
- Gunjan, S.; Singh, S.K.; Sharma, T.; Dwivedi, H.; Chauhan, B.S.; Siddiqi, M.I.; Tripathi, R. Mefloquine induces ROS mediated programmed cell death in malaria parasite: Plasmodium. Apoptosis 2016, 21, 955–964. [Google Scholar] [CrossRef]
- Giovanella, F.; Ferreira, G.K.; Prá, S.D.D.; Carvalho-Silva, M.; Gomes, L.M.; Scaini, G.; Souza, R.P. Effects of primaquine and chloroquine on oxidative stress parameters in rats. An. Acad. Bras. Ciências 2015, 87, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Kannan, R.; Kumar, K.; Sahal, D.; Kukreti, S.; Chauhan, V.S. Reaction of artemisinin with haemoglobin: Implications for antimalarial activity. Biochem. J. 2005, 385, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2013, 69, 1005–1016. [Google Scholar] [CrossRef]
- Kannan, R.; Sahal, D.; Chauhan, V.S. Heme-artemisinin adducts are crucial mediators of the ability of artemisinin to inhibit heme polymerization. Cell Chem. Biol. 2002, 9, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, C.J.; Chia, W.N.; Loh, C.C.; Li, Z.; Lee, Y.M.; Lin, Q. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef]
- Kavishe, R.A.; Koenderink, J.B.; Alifrangis, M. Oxidative stress in malaria and artemisinin combination therapy: Pros and Cons. FEBS J. 2017, 284, 2579–2591. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.V.; Brooke, B.D. The role of oxidative stress in the longevity and insecticide resistance phenotype of the major malaria vectors Anopheles arabiensis and Anopheles funestus. PLoS ONE 2016, 11, e0151049. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Chatgilialoglu, C.; Ferreri, C.; Masi, A.; Melchiorre, M.; Sansone, A.; Terzidis, M.A.; Torreggiani, A. Free radicals in chemical biology: From chemical behavior to biomarker development. JoVE (J. Vis. Exp.) 2013, 74, e50379. [Google Scholar] [CrossRef] [Green Version]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [Green Version]
- Di Dalmazi, G.; Hirshberg, J.; Lyle, D.; Freij, J.B.; Caturegli, P. Reactive oxygen species in organ-specific autoimmunity. Autoimmun. Highlights 2016, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Zhong, J.; Brook, R.D.; Rajagopalan, S. Effect of particulate matter air pollution on cardiovascular oxidative stress pathways. Antioxid. Redox Signal. 2018, 28, 797–818. [Google Scholar] [CrossRef] [PubMed]
- Goldkorn, T.; Filosto, S.; Chung, S. Lung injury and lung cancer caused by cigarette smoke-induced oxidative stress: Molecular mechanisms and therapeutic opportunities involving the ceramide-generating machinery and epidermal growth factor receptor. Antioxid. Redox Signal. 2014, 21, 2149–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Vasudevan, D.M. Alcohol-induced oxidative stress. Life Sci. 2007, 81, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; López-Sánchez, R.C.; Rendón-Ramírez, A. Lipids and oxidative stress associated with ethanol-induced neurological damage. Oxid. Med. Cell. Longev. 2016, 2016, 1543809. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; Singh, S.; Siddiqi, N.J. Biomedical implications of heavy metals induced imbalances in redox systems. BioMed Res. Int. 2014, 2014, 640754. [Google Scholar] [CrossRef] [Green Version]
- Balmus, I.M.; Ciobica, A.; Antioch, I.; Dobrin, R.; Timofte, D. Oxidative stress implications in the affective disorders: Main biomarkers, animal models relevance, genetic perspectives, and antioxidant approaches. Oxid. Med. Cell. Longev. 2016, 2016, 3975101. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Cho, S.G. Correlation between oxidative stress, nutrition, and cancer initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef] [Green Version]
- Sobolewski, P.; Gramaglia, I.; Frangos, J.A.; Intaglietta, M.; Van Der Heyde, H. Plasmodium berghei resists killing by reactive oxygen species. Infect. Immun. 2005, 73, 6704–6710. [Google Scholar] [CrossRef] [Green Version]
- Balaji, S.N.; Trivedi, V. Extracellular methemoglobin mediated early ROS spike triggers osmotic fragility and RBC destruction: An insight into the enhanced hemolysis during malaria. Indian J. Clin. Biochem. 2012, 27, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, R.; Marrocco, T.; Skorokhod, O.A.; Barbosa, A.; Nhabomba, A.; Manaca, M.N.; Schwarzer, E. Blood oxidative stress markers and Plasmodium falciparum malaria in non-immune African children. Br. J. Haematol. 2014, 164, 438–450. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Trovoada, M.J.; Martins, M.; Mansour, R.B.; Sambo, M.R.; Fernandes, A.B.; Gonçalves, L.A.; Penha-Gonçalves, C. NOS2 variants reveal a dual genetic control of nitric oxide levels, susceptibility to Plasmodium infection, and cerebral malaria. Infect. Immun. 2014, 82, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Cabrales, P.; Zanini, G.M.; Meays, D.; Frangos, J.A.; Carvalho, L.J. Nitric oxide protection against murine cerebral malaria is associated with improved cerebral microcirculatory physiology. J. Infect. Dis. 2011, 203, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Jeney, V.; Ramos, S.; Bergman, M.L.; Bechmann, I.; Tischer, J.; Ferreira, A.; Soares, M.P. Control of disease tolerance to malaria by nitric oxide and carbon monoxide. Cell Rep. 2014, 8, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Dzodzomenyo, M.; Ghansah, A.; Ensaw, N.; Dovie, B.; Bimi, L.; Quansah, R.; Amoani, B. Inducible nitric oxide synthase 2 promoter polymorphism and malaria disease severity in children in Southern Ghana. PLoS ONE 2018, 13, e0202218. [Google Scholar] [CrossRef]
- Martins, Y.C.; Zanini, G.M.; Frangos, J.A.; Carvalho, L.J. Efficacy of different nitric oxide-based strategies in preventing experimental cerebral malaria by Plasmodium berghei ANKA. PLoS ONE 2012, 7, e32048. [Google Scholar] [CrossRef] [Green Version]
- Ong, P.K.; Melchior, B.; Martins, Y.C.; Hofer, A.; Orjuela-Sánchez, P.; Cabrales, P.; Carvalho, L.J. Nitric oxide synthase dysfunction contributes to impaired cerebroarteriolar reactivity in experimental cerebral malaria. PLoS Pathog. 2013, 9, e1003444. [Google Scholar] [CrossRef]
- Yeo, T.W.; Lampah, D.A.; Kenangalem, E.; Tjitra, E.; Weinberg, J.B.; Granger, D.L.; Anstey, N.M. Decreased endothelial nitric oxide bioavailability, impaired microvascular function, and increased tissue oxygen consumption in children with falciparum malaria. J. Infect. Dis. 2014, 210, 1627–1632. [Google Scholar] [CrossRef] [Green Version]
- Barber, B.E.; William, T.; Grigg, M.J.; Piera, K.A.; Chen, Y.; Wang, H.; Anstey, N.M. Nitric oxide–dependent endothelial dysfunction and reduced arginine bioavailability in Plasmodium vivax malaria but no greater increase in intravascular hemolysis in severe disease. J. Infect. Dis. 2016, 214, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, J.B.; Volkheimer, A.D.; Rubach, M.P.; Florence, S.M.; Mukemba, J.P.; Kalingonji, A.R.; Mwaikambo, E.D. Monocyte polarization in children with falciparum malaria: Relationship to nitric oxide insufficiency and disease severity. Sci. Rep. 2016, 6, 29151. [Google Scholar] [CrossRef]
- Bangirana, P.; Conroy, A.L.; Opoka, R.O.; Hawkes, M.T.; Hermann, L.; Miller, C.; Kain, K.C. Inhaled nitric oxide and cognition in pediatric severe malaria: A randomized double-blind placebo controlled trial. PLoS ONE 2018, 13, e0191550. [Google Scholar] [CrossRef] [Green Version]
- Lwanira, C.N.; Kironde, F.; Kaddumukasa, M.; Swedberg, G. Prevalence of polymorphisms in glucose-6-phosphate dehydrogenase, sickle haemoglobin and nitric oxide synthase genes and their relationship with incidence of uncomplicated malaria in Iganga, Uganda. Malar. J. 2017, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, K.P.; Bali, P.; Anwar, S.; Kaul, A.; Singh, O.P.; Sohail, M. iNOS polymorphism modulates iNOS/NO expression via impaired antioxidant and ROS content in P. vivax and P. falciparum infection. Redox Biol. 2018, 15, 192–206. [Google Scholar] [CrossRef]
- Perkins, D.J.; Were, T.; Davenport, G.C.; Kempaiah, P.; Hittner, J.B.; Ong’echa, J.M. Severe malarial anemia: Innate immunity and pathogenesis. Int. J. Biol. Sci. 2011, 7, 1427–1442. [Google Scholar] [CrossRef] [Green Version]
- Ihekwereme, C.P.; Esimone, C.O.; Nwanegbo, E.C. Hemozoin inhibition and control of clinical malaria. Adv. Pharmacol. Sci. 2014, 2014, 984150. [Google Scholar] [CrossRef] [Green Version]
- Corbett, Y.; D’Alessandro, S.; Parapini, S.; Scaccabarozzi, D.; Kalantari, P.; Zava, S.; Basilico, N. Interplay between Plasmodium falciparum haemozoin and l-arginine: Implication for nitric oxide production. Malar. J. 2018, 17, 456. [Google Scholar] [CrossRef]
- Moreira, D.R.; Uberti, A.C.M.G.; Gomes, A.R.Q.; Ferreira, M.E.S.; Barbosa, A.S.; Varela, E.L.P.; Percário, S. Dexamethasone increased the survival rate in Plasmodium berghei-infected mice. Sci. Rep. 2021, 11, 2623. [Google Scholar] [CrossRef]
- Barbosa, A.S.; Temple, M.C.R.; Varela, E.L.P.; Gomes, A.R.Q.; Silveira, E.L.; de Carvalho, E.P.; Percario, S. Inhibition of nitric oxide synthesis promotes increased mortality despite the reduction of parasitemia in Plasmodium berghei-infected mice. Res. Soc. Dev. 2021, 10, e27810111805. [Google Scholar] [CrossRef]
- Macchi, B.M.; Miranda, F.J.B.; de Souza, F.S.; de Carvalho, E.C.Q.; Albernaz, A.P.; do Nascimento, J.L.M.; DaMatta, R. Chickens treated with a nitric oxide inhibitor became more resistant to Plasmodium gallinaceum infection due to reduced anemia, thrombocytopenia and inflammation. Vet. Res. 2013, 44, 8. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, J.B.; Yeo, T.W.; Mukemba, J.P.; Florence, S.M.; Volkheimer, A.D.; Wang, H.; Anstey, N.M. Dimethylarginines: Endogenous inhibitors of nitric oxide synthesis in children with falciparum malaria. J. Infect. Dis. 2014, 210, 913–922. [Google Scholar] [CrossRef]
- Chertow, J.H.; Alkaitis, M.S.; Nardone, G.; Ikeda, A.K.; Cunnington, A.J.; Okebe, J.; Ackerman, H. Plasmodium infection is associated with impaired hepatic dimethylarginine dimethylamino hydrolase activity and disruption of nitric oxide synthase inhibitor/substrate homeostasis. PLoS Pathog. 2015, 11, e1005119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peranzoni, E.; Marigo, I.; Dolcetti, L.; Ugel, S.; Sonda, N.; Taschin, E.; Zanovello, P. Role of arginine metabolism in immunity and immunopathology. Immunobiology 2008, 212, 795–812. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Pan, Y.; Li, Y.; Cui, L.; Cao, Y. Supplement of L-Arg improves protective immunity during early-stage Plasmodium yoelii 17XL infection. Parasite Immunol. 2012, 34, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Brussee, J.M.; Yeo, T.W.; Lampah, D.A.; Anstey, N.M.; Duffull, S.B. Pharmacokinetic-pharmacodynamic model for the effect of L-arginine on endothelial function in patients with moderately severe falciparum malaria. Antimicrob. Agents Chemother. 2016, 60, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkes, M.T.; Conroy, A.L.; Opoka, R.O.; Hermann, L.; Thorpe, K.E.; McDonald, C.; Kain, K.C. Inhaled nitric oxide as adjunctive therapy for severe malaria: A randomized controlled trial. Malar. J. 2015, 14, 421. [Google Scholar] [CrossRef] [Green Version]
- Kayano, A.C.A.V.; Dos-Santos, J.C.K.; Bastos, M.F.; Carvalho, L.J.; Aliberti, J.; Costa, F.T. Pathophysiological mechanisms in gaseous therapies for severe malaria. Infect. Immun. 2016, 84, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, M.; Opoka, R.O.; Namasopo, S.; Miller, C.; Conroy, A.L.; Serghides, L.; Kain, K.C. Nitric oxide for the adjunctive treatment of severe malaria: Hypothesis and rationale. Med. Hypotheses 2011, 77, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Alkaitis, M.S.; Wang, H.; Ikeda, A.K.; Rowley, C.A.; MacCormick, I.J.C.; Chertow, J.H.; Billker, O.; Suffredini, A.F.; Roberts, D.J.; Taylor, T.E.; et al. Decreased Rate of Plasma Arginine Appearance in Murine Malaria May Explain Hypoargininemia in Children with Cerebral Malaria. J. Infect. Dis. 2016, 214, 1840–1849. [Google Scholar] [CrossRef] [Green Version]
- Gramaglia, I.; Velez, J.; Chang, Y.S.; Caparros-Wanderley, W.; Combes, V.; Grau, G.; Stins, M.F.; van der Heyde, H.C. Citrulline protects mice from experimental cerebral malaria by ameliorating hypoargininemia, urea cycle changes and vascular leak. PLoS ONE 2019, 14, e0213428. [Google Scholar] [CrossRef] [Green Version]
- Yeo, T.W.; Lampah, D.A.; Rooslamiati, I.; Gitawati, R.; Tjitra, E.; Kenangalem, E.; Price, R.N.; Duffull, S.B.; Anstey, N.M. A randomized pilot study of L-arginine infusion in severe falciparum malaria: Preliminary safety, efficacy and pharmacokinetics. PLoS ONE 2013, 8, e69587. [Google Scholar] [CrossRef]
- Awasthi, V.; Chauhan, R.; Chattopadhyay, D.; Das, J. Effect of L-arginine on the growth of Plasmodium falciparum and immune modulation of host cells. J. Vector Borne Dis. 2017, 54, 139. [Google Scholar]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Pieme, C.A.; Tatangmo, J.A.; Simo, G.; Nya, P.C.B.; Moor, V.J.A.; Moukette, B.M.; Sobngwi, E. Relationship between hyperglycemia, antioxidant capacity and some enzymatic and non-enzymatic antioxidants in African patients with type 2 diabetes. BMC Res. Notes 2017, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Skorokhod, O.A.; Khoo, S.K.; Aguilar, R.; Wiertsema, S.; Nhabomba, A.J.; LeSouëf, P.N. Plasma advanced oxidative protein products are associated with anti-oxidative stress pathway genes and malaria in a longitudinal cohort. Malar. J. 2014, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.C.; Hasan, M.; Gupta, H.; Geetha, K.; Rai, P.S.; Hande, M.H.; Satyamoorthy, K. Host genetic variations in glutathione-S-transferases, superoxide dismutases and catalase genes influence susceptibility to malaria infection in an Indian population. Mol. Genet. Genom. 2015, 290, 1155–1168. [Google Scholar] [CrossRef]
- Farombi, E.O.; Shyntum, Y.Y.; Emerole, G.O. Influence of chloroquine treatment and Plasmodium falciparum malaria infection on some enzymatic and non-enzymatic antioxidant defense indices in humans. Drug Chem. Toxicol. 2003, 26, 59–71. [Google Scholar] [CrossRef]
- Chaudhari, R.; Sharma, S.; Patankar, S. Glutathione and thioredoxin systems of the malaria parasite Plasmodium falciparum: Partners in crime? Biochem. Biophys. Res. Commun. 2017, 488, 95–100. [Google Scholar] [CrossRef]
- Babalola, A.S.; Jonathan, J.; Michael, B.E. Oxidative stress and anti-oxidants in asymptomatic malaria-positive patients: A hospital-based cross-sectional Nigerian study. Egypt. J. Intern. Med. 2020, 32, 32. [Google Scholar] [CrossRef]
- Tyagi, A.G.; Tyagi, R.A.; Choudhary, P.R.; Shekhawat, J.S. Study of antioxidant status in malaria patients. Int. J. Res. Med. Sci. 2017, 5, 1649. [Google Scholar] [CrossRef] [Green Version]
- Oluba, O.M. Erythrocyte lipid and antioxidant changes in Plasmodium falciparum-infected children attending mother and child hospital in Akure, Nigeria. Pak. J. Biol. Sci. 2019, 22, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, C.; Mascarenhas-Netto, R.C.; Lalwani, P.; Melo, G.C.; Magalhães, B.M.; Alexandre, M.A.; Lima, E.S. Lipid peroxidation and antioxidant enzymes activity in Plasmodium vivax malaria patients evolving with cholestatic jaundice. Malar. J. 2013, 12, 315. [Google Scholar] [CrossRef] [Green Version]
- Schulze, P.C.; Liu, H.; Choe, E.; Yoshioka, J.; Shalev, A.; Bloch, K.D.; Lee, R.T. Nitric oxide–dependent suppression of thioredoxin-interacting protein expression enhances thioredoxin activity. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2666–2672. [Google Scholar] [CrossRef] [Green Version]
- Belorgey, D.; Antoine Lanfranchi, D.; Davioud-Charvet, E. 1, 4-naphthoquinones and other NADPH-dependent glutathione reductase-catalyzed redox cyclers as antimalarial agents. Curr. Pharm. Des. 2013, 19, 2512–2528. [Google Scholar] [CrossRef]
- Vega-Rodríguez, J.; Pastrana-Mena, R.; Crespo-Lladó, K.N.; Ortiz, J.G.; Ferrer-Rodríguez, I.; Serrano, A.E. Implications of glutathione levels in the Plasmodium berghei response to chloroquine and artemisinin. PLoS ONE 2015, 10, e0128212. [Google Scholar] [CrossRef] [Green Version]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef]
- Aqeel, S.; Naheda, A.; Raza, A.; Khan, K.; Khan, W. Differential status and significance of non-enzymatic antioxidants (reactive oxygen species scavengers) in malaria and dengue patients. Acta Trop. 2019, 195, 127–134. [Google Scholar] [CrossRef]
- Nwachukwu, N.F.; Mishra, M.; Marcelle, T.; Owolabi, J.B. Effects of vitamins in augmentation of malaria treatment: A review. Asian Pac. J. Health Sci. 2016, 3, 34–43. [Google Scholar] [CrossRef]
- Barffour, M.A.; Schulze, K.J.; Moss, W.J.; West Jr, K.P.; Chileshe, J.; Siamusantu, W.; Palmer, A.C. Relative contributions of malaria, inflammation, and deficiencies of iron and vitamin A to the burden of anemia during low and high malaria seasons in rural Zambian children. J. Pediatr. 2019, 213, 74–81. [Google Scholar] [CrossRef]
- Burki, T.K. Malaria and malnutrition: Niger’s twin crises. Lancet 2013, 382, 587–588. [Google Scholar] [CrossRef]
- Abdulkareem, B.O.; Adam, A.O.; Ahmed, A.O.; Mariam, A.A.; Samuel, U.U. Malaria-induced anaemia and serum micronutrients in asymptomatic Plasmodium falciparum infected patients. J. Parasit. Dis. 2017, 41, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Akpotuzor, J.O.; Udoh, A.E.; Etukudo, M.H. Total antioxidant status and other antioxidant agent levels in children with P. falciparum infection in Calabar, Nigeria. Int. J. Biomed. Lab. Sci. 2012, 1, 35–39. [Google Scholar]
- Uwah, A.F.; Otitoju, O.; Ndem, J.I.; Akpanabiatu, M.I.; Akpanyung, E.O. Effect of malaria infection on serum copper, iron and beta-carotene levels in Nigerian children. Eur. J. Sci. Res. 2012, 86, 573–579. [Google Scholar]
- Alexandre, M.A.A.; Benzecry, S.G.; Siqueira, A.M.; Vitor-Silva, S.; Melo, G.C.; Monteiro, W.M.; Alecrim, M.D.G.C. The association between nutritional status and malaria in children from a rural community in the Amazonian region: A longitudinal study. PLoS Negl. Trop. Dis. 2015, 9, e0003743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakwe, N.; Bigoga, J.; Ngondi, J.; Njeambosay, B.; Esemu, L.; Kouambeng, C.; Oben, J. Relationship between malaria, anaemia, nutritional and socio-economic status amongst under-ten children, in the North Region of Cameroon: A cross-sectional assessment. PLoS ONE 2019, 14, e0218442. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.C.; Siamusantu, W.; Chileshe, J.; Schulze, K.J.; Barffour, M.; Craft, N.E.; West Jr, K.P. Provitamin A–biofortified maize increases serum β-carotene, but not retinol, in marginally nourished children: A cluster-randomized trial in rural Zambia. Am. J. Clin. Nutr. 2016, 104, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic effects of inflammation on vitamin A and carotenoids in humans and animal models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Aghedo, F.I.; Shehu, R.A.; Umar, R.A.; Jiya, M.N.; Erhabor, O. Antioxidant vitamin levels among preschool children with uncomplicated Plasmodium falciparum malaria in Sokoto, Nigeria. J. Multidiscip. Healthc. 2013, 6, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Lawal, O.A.; Adegoke, S.A.; Oseni, S.B.; Oyelami, O.A. Low serum vitamin A is prevalent in underfive children with severe malaria and is associated with increased risk of death. J. Infect. Dev. Ctries. 2018, 12, 365–372. [Google Scholar] [CrossRef]
- Bivona, G.; Agnello, L.; Sasso, B.L.; Scazzone, C.; Butera, D.; Gambino, C.M.; Ciaccio, M. Vitamin D in malaria: More hypotheses than clues. Heliyon 2019, 5, e01183. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, C.E.; Guerin, P.J.; Berthé, F.; Grais, R.F.; Isanaka, S. Malaria and nutritional status among children with severe acute malnutrition in Niger: A prospective cohort study. Clin. Infect. Dis. 2018, 67, 1027–1034. [Google Scholar] [CrossRef]
- Camara, A.; Haddad, M.; Reybier, K.; Traoré, M.S.; Baldé, M.A.; Royo, J.; Aubouy, A. Terminalia albida treatment improves survival in experimental cerebral malaria through reactive oxygen species scavenging and anti-inflammatory properties. Malar. J. 2019, 18, 431. [Google Scholar] [CrossRef]
- Ouattara, L.P.; Sanon, S.; Mahiou-Leddet, V.; Gansané, A.; Baghdikian, B.; Traoré, A.; Sirima, S.B. In vitro antiplasmodial activity of some medicinal plants of Burkina Faso. Parasitol. Res. 2014, 113, 405–416. [Google Scholar] [CrossRef]
- Sadiq, M.B.; Tharaphan, P.; Chotivanich, K.; Tarning, J.; Anal, A.K. In vitro antioxidant and antimalarial activities of leaves, pods and bark extracts of Acacia nilotica (L.) Del. BMC Complement. Altern. Med. 2017, 17, 372. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.Y.; Zhang, Y.R.; Ji, J.B.; Xing, J. Investigation of the component in Artemisia annua L. leading to enhanced antiplasmodial potency of artemisinin via regulation of its metabolism. J. Ethnopharmacol. 2017, 207, 86–91. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; Gong, M.; Wang, M. Combination of artemisinin-based natural compounds from Artemisia annua L. for the treatment of malaria: Pharmacodynamic and pharmacokinetic studies. Phytother. Res. 2018, 32, 1415–1420. [Google Scholar] [CrossRef]
- Conrad, O.A.; Uche, A.I. Assessment of in vivo antioxidant properties of Dacryodes edulis and Ficus exasperata as anti-malarial plants. Asian Pac. J. Trop. Dis. 2013, 3, 294–300. [Google Scholar] [CrossRef]
- Omonhinmin, C.A.; Dike, I.P.; Rotimi, S.O. Phytochemical, cytotoxicity and antioxidant activities of five anti-malaria plants. Res. J. Med. Plants 2015, 9, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Djehoue, R.; Amoussa, A.M.O.; Sanni, A.; Lagnika, L. Phytochemical composition and antioxidant property of Dissotis rotundifolia used for malaria management in south Benin. J. Med. Plants 2020, 8, 23–29. [Google Scholar]
- Bankole, A.E.; Adekunle, A.A.; Sowemimo, A.A.; Umebese, C.E.; Abiodun, O.; Gbotosho, G.O. Phytochemical screening and in vivo antimalarial activity of extracts from three medicinal plants used in malaria treatment in Nigeria. Parasitol. Res. 2016, 115, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Kpadonou, D.; Kpoviessi, S.E.; Bero, J.; Agbani, P.; Gbaguidi, F.; Kpadonou-Kpoviessi, B.E.E.; Quetin-Leclercq, J. Chemical composition, in vitro antioxidant and antiparasitic properties of the essential oils of three plants used in traditional medicine in Benin. J. Med. Plants Res. 2019, 13, 384–395. [Google Scholar] [CrossRef]
- Franco, C.J.P.; Ferreira, O.O.; Moraes, A.A.B.; Varela, E.L.P.; Nascimento, L.D.N.; Percario, S.; Andrade, E.H.A. Chemical composition and antioxidant activity of essential oils from Eugenia patrisii vahl, E. punicifolia (Kunth) dc., and Myrcia tomentosa (Aubl.) dc., leaf of family Myrtaceae. Molecules 2021, 26, 3292. [Google Scholar] [CrossRef]
- Brandão, D.L.N.; Martins, M.T.; Silva, A.O.; Almeida, A.D.; de Paula, R.C.; de Oliveira, A.B.; Ferreira, M.C.; Gomes, A.T.A.; Vasconcelos, F.; Pereira, W.L.A.; et al. Anti-malarial activity and toxicity of Aspidosperma nitidum Benth: A plant used in traditional medicine in the Brazilian Amazon. Res. Soc. Dev. 2020, 9, e5059108817. [Google Scholar] [CrossRef]
- Vale, V.V.; Cruz, J.N.; Viana, G.M.R.; Póvoa, M.M.; Brasil, D.D.S.B.; Dolabela, M.F. Naphthoquinones isolated from Eleutherine plicata herb: In vitro antimalarial activity and molecular modeling to investigate their binding modes. Med. Chem. Res. 2020, 29, 487–494. [Google Scholar] [CrossRef]
- Gomes, A.R.Q.; Brígido, H.P.C.; Vale, V.V.; Correa-Barbosa, J.; Percário, S.; Dolabela, M.F. Antimalarial potential of quinones isolated from plants: An integrative review. Res. Soc. Dev. 2021, 10, e38210212507. [Google Scholar] [CrossRef]
- Gomes, A.R.Q.; da Rocha Galucio, N.C.; de Albuquerque, K.C.O.; Brígido, H.P.C.; Varela, E.L.P.; Castro, A.L.G.; Vale, V.V.; Bahia, M.O.; Burbano, R.M.R.; de Molfeta, F.A.; et al. Toxicity evaluation of Eleutherine plicata Herb. extracts and possible cell death mechanism. Toxicol. Rep. 2021, 8, 1480–1487. [Google Scholar] [CrossRef]
- Nagy, P. Kinetics and mechanisms of thiol–disulfide exchange covering direct substitution and thiol oxidation-mediated pathways. Antioxid. Redox Signal. 2013, 18, 1623–1641. [Google Scholar] [CrossRef] [Green Version]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Kedarisetty, C.K.; Samaga, B.L.; Vidyasagar, S.; Venkataraman, J. Oral melatonin improves the detection of parasitemia in malaria. J. Infect. Dev. Ctries. 2020, 14, 1327–1331. [Google Scholar] [CrossRef]
- Scarpelli, P.H.; Tessarin-Almeida, G.; Viçoso, K.L.; Lima, W.R.; Borges-Pereira, L.; Meissner, K.A.; Wrenger, C.; Raffaello, A.; Rizzuto, R.; Pozzan, T.; et al. Melatonin activates FIS 1, DYN 1, and DYN 2 Plasmodium falciparum related-genes for mitochondria fission: Mitoemerald-GFP as a tool to visualize mitochondria structure. J. Pineal Res. 2019, 66, e12484. [Google Scholar] [CrossRef] [Green Version]
- Dias, B.K.; Nakabashi, M.; Alves, M.R.R.; Portella, D.P.; Santos, B.M.; Almeida, F.C.S.; Ribeiro, R.Y.; Schuck, D.C.; Jordão, A.K.; Garcia, C.R.S. The Plasmodium falciparum eIK1 kinase (PfeIK1) is central for melatonin synchronization in the human malaria parasite. Melatotosil blocks melatonin action on parasite cell cycle. J. Pineal Res. 2020, 69, e12685. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.K.; Tessarin-Almeida, G.; Dias, B.K.M.; Pereira, P.S.; Costa, F.; Przyborski, J.M.; Garcia, C.R.S. A nuclear protein, PfMORC confers melatonin dependent synchrony of the human malaria parasite P. falciparum in the asexual stage. Sci. Rep. 2021, 11, 2057. [Google Scholar] [CrossRef] [PubMed]
- Lima, W.R.; Tessarin-Almeida, G.; Rozanski, A.; Parreira, K.S.; Moraes, M.S.; Martins, D.C.; Hashimoto, R.F.; Galante, P.A.F.; Garcia, C.R.S. Signaling transcript profile of the asexual intraerythrocytic development cycle of Plasmodium falciparum induced by melatonin and cAMP. Genes Cancer 2016, 7, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Olayan, E.M.; El-Khadragy, M.F.; Abdel Moneim, A.E. The protective properties of melatonin against aluminium-induced neuronal injury. Int. J. Exp. Pathol. 2015, 96, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Lai, C.J.; Tsai, M.H.; Wu, Y.C.; Chen, K.T.; Jou, M.J.; Fu, P.I.; Wu, C.H.; Wei, I.H. Effects of melatonin on the nitric oxide system and protein nitration in the hypobaric hypoxic rat hippocampus. BMC Neurosci. 2015, 16, 61. [Google Scholar] [CrossRef] [Green Version]
- Lopez, A.; Ortiz, F.; Doerrier, C.; Venegas, C.; Fernández-Ortiz, M.; Aranda, P.; Díaz-Casado, M.E.; Fernández-Gil, B.; Barriocanal-Casado, E.; Escames, G.; et al. Mitochondrial impairment and melatonin protection in parkinsonian mice do not depend of inducible or neuronal nitric oxide synthases. PLoS ONE 2017, 12, e0183090. [Google Scholar] [CrossRef]
- Suwannakot, K.; Sritawan, N.; Prajit, R.; Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welba, J.U. Melatonin protects against the side-effects of 5-fluorouracil on hippocampal neurogenesis and ameliorates antioxidant activity in an adult rat hippocampus and prefrontal cortex. Antioxidants 2021, 10, 615. [Google Scholar] [CrossRef]
- Ataide, B.J.D.A.; Kauffmann, N.; Mendes, N.S.F.; Torres, M.L.M.; Anjos, L.M.; Passos, A.C.F.; Moraes, S.A.S.; Batista, E.J.O.; Herculano, A.M.; Oliveira, K.R.H.M. Melatonin prevents brain damage and neurocognitive impairment induced by Plasmodium berghei ANKA infection in murine model of cerebral malaria. Front. Cell. Infect. Microbiol. 2020, 516. [Google Scholar] [CrossRef]
- Villaverde, C.; Namazzi, R.; Shabani, E.; Park, G.S.; Datta, D.; Hanisch, B.; Opoka, R.O.; John, C.C. Retinopathy-positive cerebral malaria is associated with greater inflammation, blood-brain barrier breakdown, and neuronal damage than retinopathy-negative cerebral malaria. J. Pediatr. Infect. Dis. Soc. 2020, 9, 580–586. [Google Scholar] [CrossRef]
- Verma, A.K.; Singh, S.; Garg, G.; Rizvi, S.I. Melatonin exerts neuroprotection in a chronodisrupted rat model through reduction in oxidative stress and modulation of autophagy. Chronobiol. Int. 2022, 39, 45–56. [Google Scholar] [CrossRef]
- Zhao, S.; Duan, H.; Yang, Y.; Yan, X.; Fan, K. Fenozyme protects the integrity of the blood–brain barrier against experimental cerebral malaria. Nano Lett. 2019, 19, 8887–8895. [Google Scholar] [CrossRef]
- Bastian, T.W.; Rao, R.; Tran, P.V.; Georgieff, M.K. The effects of early-life iron deficiency on brain energy metabolism. Neurosci. Insights 2020, 15, 1–12. [Google Scholar] [CrossRef]
- Sahu, T.; Boisson, B.; Lacroix, C.; Bischoff, E.; Richier, Q.; Formaglio, P.; Thiberge, S.; Dobrescu, I.; Ménard, R.; Baldacci, P. ZIPCO, a putative metal ion transporter, is crucial for Plasmodium liver-stage development. EMBO Mol. Med. 2014, 6, 1387–1397. [Google Scholar] [CrossRef]
- Gwamaka, M.; Kurtis, J.D.; Sorensen, B.E.; Holte, S.; Morrison, R.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. Iron deficiency protects against severe Plasmodium falciparum malaria and death in young children. Clin. Infect. Dis. 2012, 54, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.A.; Goheen, M.M.; Cerami, C. Influence of host iron status on Plasmodium falciparum infection. Front. Pharmacol. 2014, 5, 84. [Google Scholar] [CrossRef]
- Thipubon, P.; Uthaipibull, C.; Kamchonwongpaisan, S.; Tipsuwan, W.; Srichairatanakool, S. Inhibitory effect of novel iron chelator, 1-(N-acetyl-6-aminohexyl)-3-hydroxy-2-methylpyridin-4-one (CM1) and green tea extract on growth of Plasmodium falciparum. Malar. J. 2015, 14, 382. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.; Checkley, L.; Ferdig, M.T.; Vennerstrom, J.L.; Miller, M.J. Synthesis and antimalarial activity of amide and ester conjugates of siderophores and ozonides. Biometals 2022, 1–6. [Google Scholar] [CrossRef]
- Dey, S.; Bindu, S.; Goyal, M.; Pal, C.; Alam, A.; Iqbal, M.S.; Kumar, R.; Sarkar, S.; Bandyopadhyay, U. Impact of intravascular hemolysis in malaria on liver dysfunction: Involvement of hepatic free heme overload, NF-κB activation, and neutrophil infiltration. J. Biol. Chem. 2012, 287, 26630–26646. [Google Scholar] [CrossRef] [Green Version]
- Chandrasiri, U.P.; Fowkes, F.J.I.; Richards, J.S.; Langer, C.; Fan, Y.M.; Taylor, S.M.; Beeson, J.G.; Dewey, K.G.; Maleta, K.; Ashorn, P.; et al. The impact of lipid-based nutrient supplementation on anti-malarial antibodies in pregnant women in a randomized controlled trial. Malar. J. 2015, 14, 193. [Google Scholar] [CrossRef] [Green Version]
- Barua, P.; Chandrasiri, U.P.; Beeson, J.G.; Dewey, K.G.; Maleta, K.; Ashorn, P.; Rogerson, S.J. Effect of nutrient supplementation on the acquisition of humoral immunity to Plasmodium falciparum in young Malawian children. Malar. J. 2018, 17, 74. [Google Scholar] [CrossRef] [Green Version]
- Prado, E.L.; Ashorn, U.; Phuka, J.; Maleta, K.; Sadalaki, J.; Oaks, B.M.; Haskell, M.; Allen, L.H.; Vosti, S.A.; Ashorn, P.; et al. Associations of maternal nutrition during pregnancy and post-partum with maternal cognition and caregiving. Matern. Child Nutr. 2018, 14, e12546. [Google Scholar] [CrossRef]
- Mireku, M.O.; Davidson, L.L.; Boivin, M.J.; Zoumenou, R.; Massougbodji, A.; Cot, M.; Bodeau-Livinec, F. Prenatal iron deficiency, neonatal ferritin, and infant cognitive function. Pediatrics 2016, 138, e20161319. [Google Scholar] [CrossRef] [Green Version]
- Brabin, B.; Gies, S.; Roberts, S.A.; Diallo, S.; Lompo, O.M.; Kazienga, A.; Brabin, L.; Ouedraogo, S.; Tinto, H. Excess risk of preterm birth with periconceptional iron supplementation in a malaria endemic area: Analysis of secondary data on birth outcomes in a double blind randomized controlled safety trial in Burkina Faso. Malar. J. 2019, 18, 161. [Google Scholar] [CrossRef] [Green Version]
- Gies, S.; Roberts, S.A.; Diallo, S.; Lompo, O.M.; Tinto, H.; Brabin, L. Risk of malaria in young children after periconceptional iron supplementation. Matern. Child Nutr. 2021, 17, e13106. [Google Scholar] [CrossRef]
- Brabin, L.; Roberts, S.A.; Gies, S.; Nelson, A.; Diallo, S.; Stewart, C.J.; Kazienga, A.; Birtles, J.; Ouedraogo, S.; Claeys, Y.; et al. Effects of long-term weekly iron and folic acid supplementation on lower genital tract infection–a double blind, randomised controlled trial in Burkina Faso. BMC Med. 2017, 15, 206. [Google Scholar] [CrossRef] [Green Version]
- Gies, S.; Diallo, S.; Stewart, C.J.; Kazienga, A.; Powney, M.; Brabin, L.; Ouedraogo, S.; Swinkels, D.W.; Geurts-Moespot, A.J.; Claeys, Y.; et al. Effects of weekly iron and folic acid supplements on malaria risk in nulliparous women in Burkina Faso: A periconceptional, double-blind, randomized controlled noninferiority trial. J. Infect. Dis. 2018, 218, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Penna-Coutinho, J.; Aguiar, A.C.C.; Krettly, A.U. Commercial drugs containing flavonoids are active in mice with malaria and in vitro against chloroquine-resistant Plasmodium falciparum. Memórias Inst. Oswaldo Cruz 2018, 113, e180279. [Google Scholar] [CrossRef]
- Mishra, S.K.; Singh, P.; Rath, S.K. Protective effect of quercetin on chloroquine-induced oxidative stress and hepatotoxicity in mice. Malar. Res. Treat. 2013, 2013, 141734. [Google Scholar] [CrossRef] [Green Version]
- Nwalor, C.I.G.; Chinyere, U.; Nwoke, B.; Cletus, E.; Oluchi, O. Influence of nutritional status on pro and anti inflammatory cytokine balance in Plasmodium falciparum malaria children in Imo State, Nigeria. Int. J. Infect. Dis. 2016, 45, 370. [Google Scholar] [CrossRef] [Green Version]
- Ukaga, C.N.; Okoro, C.; Nwoke, B.; Ezeunala, M. The possible role of nutritional status on the pro and anti-inflammatory cytokine balance of children with malaria from Imo State, Nigeria. Int. J. Infect. Dis. 2016, 45, 384. [Google Scholar] [CrossRef] [Green Version]
- Gaston, R.T.; Ramroop, S. Prevalence of and factors associated with malaria in children under five years of age in Malawi, using malaria indicator survey data. Heliyon 2020, 6, e03946. [Google Scholar] [CrossRef] [PubMed]
- Ojezele, M.O.; Moke, E.G.; Onyesom, I. Impact of generic antimalarial or Phyllanthus amarus and vitamin co-administration on antioxidant status of experimental mice infested with Plasmodium berghei. Beni-Suef Univ. J. Basic Appl. Sci. 2017, 6, 260–265. [Google Scholar] [CrossRef]
- Ojezele, M.O.; Igbe, I.; Okhuarobo, A. Reproductive indices in malaria infested mice treated with antimalarials, Phyllanthus amarus combined with vitamins. Bull. Fac. Pharm. Cairo Univ. 2018, 56, 179–184. [Google Scholar] [CrossRef]
- Hamzah, J.; Batty, K.T.; Davis, W.A.; Mori, T.A.; Ching, S.Y.L.; Croft, K.D.; Davis, T.M.E. Retinol supplementation in murine Plasmodium berghei malaria: Effects on tissue levels, parasitaemia and lipid peroxidation. Int. J. Parasitol. 2007, 37, 525–537. [Google Scholar] [CrossRef]
- Mwanga-Amumpaire, J.; Ndeezi, G.; Tumwine, J.K. Effect of vitamin A adjunct therapy for cerebral malaria in children admitted to Mulago hospital: A randomized controlled trial. Afr. Health Sci. 2012, 12, 90–97. [Google Scholar] [CrossRef]
- Luong, K.V.Q.; Nguyen, L.T.H. The role of vitamin D in malaria. J. Infect. Dev. Ctries. 2015, 9, 008–019. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Iwagami, M.; Seki, T.; Kano, S.; Ota, N.; Ato, M. Dual antiplasmodial activity of vitamin D3 and its analog, 22-oxacalcitriol, by direct and indirect mechanisms. Parasitol. Int. 2017, 66, 89–99. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Isah, M.B.; Okafor, A.I.; Bashir, M.; Bisalla, M.; Umar, I.A. Effects of combined administration of vitamins C and E on some Plasmodium berghei-induced pathological changes and oxidative stress in mice. Comp. Clin. Pathol. 2012, 21, 1677–1682. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Zuwahu, M.M.; Isah, M.B.; Jatau, I.D.; Aliyu, A.B.; Umar, I.A. Effects of vitamin E administration on Plasmodium berghei induced pathological changes and oxidative stress in mice. Trop. Biomed. 2012, 29, 98–106. [Google Scholar] [CrossRef]
- Awodele, O.; Emeka, P.M.; Akintonwa, A.; Aina, O.O. Antagonistic effect of vitamin E on the efficacy of artesunate against Plasmodium berghei infection in mice. Afr. J. Biomed. Res. 2007, 10. [Google Scholar] [CrossRef]
- Sussmann, R.A.; Fotoran, W.L.; Kimura, E.A.; Katzin, A.M. Plasmodium falciparum uses vitamin E to avoid oxidative stress. Parasites Vectors 2017, 10, 461. [Google Scholar] [CrossRef] [Green Version]
- Herbas, M.S.; Okazaki, M.; Terao, E.; Xuan, X.; Arai, H.; Suzuki, H. α-Tocopherol transfer protein inhibition is effective in the prevention of cerebral malaria in mice. Am. J. Clin. Nutr. 2010, 91, 200–207. [Google Scholar] [CrossRef]
- Herbas, M.S.; Ueta, Y.Y.; Ichikawa, C.; Chiba, M.; Ishibashi, K.; Shichiri, M.; Suzuki, H. Alpha-tocopherol transfer protein disruption confers resistance to malarial infection in mice. Malar. J. 2010, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Chugh, M.; Sundararaman, V.; Kumar, S.; Reddy, V.S.; Siddiqui, W.A.; Stuart, K.D.; Malhotra, P. Protein complex directs hemoglobin-to-hemozoin formation in Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2013, 110, 5392–5397. [Google Scholar] [CrossRef] [Green Version]
- Rudrapal, M.; Chetia, D. Endoperoxide antimalarials: Development, structural diversity and pharmacodynamic aspects with reference to 1,2,4-trioxane-based structural scaffold. Drug Des. Devel. 2016, 10, 3575–3590. [Google Scholar] [CrossRef] [Green Version]
- Egwu, C.O.; Augereau, J.M.; Reybier, K.; Benoit-Vical, F. Reactive Oxygen Species as the Brainbox in Malaria Treatment. Antioxidants 2021, 10, 1872. [Google Scholar] [CrossRef]
- Bopp, S.; Magistrado, P.; Wong, W.; Schaffner, S.F.; Mukherjee, A.; Lim, P.; Dhorda, M.; Amaratunga, C.; Woodrow, C.J.; Ashley, E.A.; et al. Plasmepsin II-III copy number accounts for bimodal piperaquine resistance among Cambodian Plasmodium falciparum. Nat. Commun. 2018, 9, 1769. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO: Artemisinin Resistance and Artemisinin-Based Combination Therapy Efficacy (Status Report—August 2018). Available online: https://apps.who.int/iris/bitstream/handle/10665/274362/WHO-CDS-GMP-2018.18-eng.pdf?ua=1 (accessed on 12 May 2022).
- Fitri, L.E.; Iskandar, A.; Permatasari, N.; Gunawan, J.A.; Indrawan, K. Efek kombinasi artemisinin dan N-acetylcysteine Terhadap kadar malondialdehyde (MDA) otak dan paru mencit galur Balb/c yang diinfeksi Plasmodium berghei. J. Kedokt. Brawijaya 2013, 24. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, A.; Sudjari, S. Efek sinergis klorokuin dan N-acetyl cysteine terhadap penurunan parasitemia dan penurunan kadar malondyaldehyde eritrosit mencit yang diinfeksi Plasmodium berghei. Indones. J. Clin. Pathol. Med. Lab. 2016, 20, 6–11. [Google Scholar] [CrossRef]
- McKoy, M.G.; Kong-Quee, P., III; Pepple, D.J. In vitro effects of co-incubation of blood with artemether/lumefantrine & vitamin C on the viscosity & elasticity of blood. Indian J. Med. Res. 2016, 143, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marwaha, N. Ascorbic acid co-administration with artemisinin based combination therapies in falciparum malaria. Indian J. Med. Res. 2016, 143, 539. [Google Scholar] [CrossRef] [Green Version]
- Iyawe, H.O.T.; Onigbinde, A.O. Chloroquine and vitamin combination effects on P. berghei induced oxidative stress. Int. J. Biochem. Res. Rev. 2012, 2, 120. [Google Scholar] [CrossRef] [Green Version]
- Ebohon, O.; Irabor, F.; Omoregie, E.S. Ascorbic acid coadministration with artesunate–amodiaquine, up-regulated antioxidant enzymes gene expression in bone marrow cells and elicited biochemical changes in Plasmodium berghei-infected mice. SN Appl. Sci. 2021, 3, 6. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, A.R.Q.; Cunha, N.; Varela, E.L.P.; Brígido, H.P.C.; Vale, V.V.; Dolabela, M.F.; De Carvalho, E.P.; Percário, S. Oxidative Stress in Malaria: Potential Benefits of Antioxidant Therapy. Int. J. Mol. Sci. 2022, 23, 5949. https://doi.org/10.3390/ijms23115949
Gomes ARQ, Cunha N, Varela ELP, Brígido HPC, Vale VV, Dolabela MF, De Carvalho EP, Percário S. Oxidative Stress in Malaria: Potential Benefits of Antioxidant Therapy. International Journal of Molecular Sciences. 2022; 23(11):5949. https://doi.org/10.3390/ijms23115949
Chicago/Turabian StyleGomes, Antonio Rafael Quadros, Natasha Cunha, Everton Luiz Pompeu Varela, Heliton Patrick Cordovil Brígido, Valdicley Vieira Vale, Maria Fâni Dolabela, Eliete Pereira De Carvalho, and Sandro Percário. 2022. "Oxidative Stress in Malaria: Potential Benefits of Antioxidant Therapy" International Journal of Molecular Sciences 23, no. 11: 5949. https://doi.org/10.3390/ijms23115949