
Response of Poplar and Associated Fungal Endophytic Communities to a PAH Contamination Gradient

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Microscopic Evidence of Fungal Colonization
2.2. Impact of Phenanthrene Gradient on Plant Biomass and Fitness
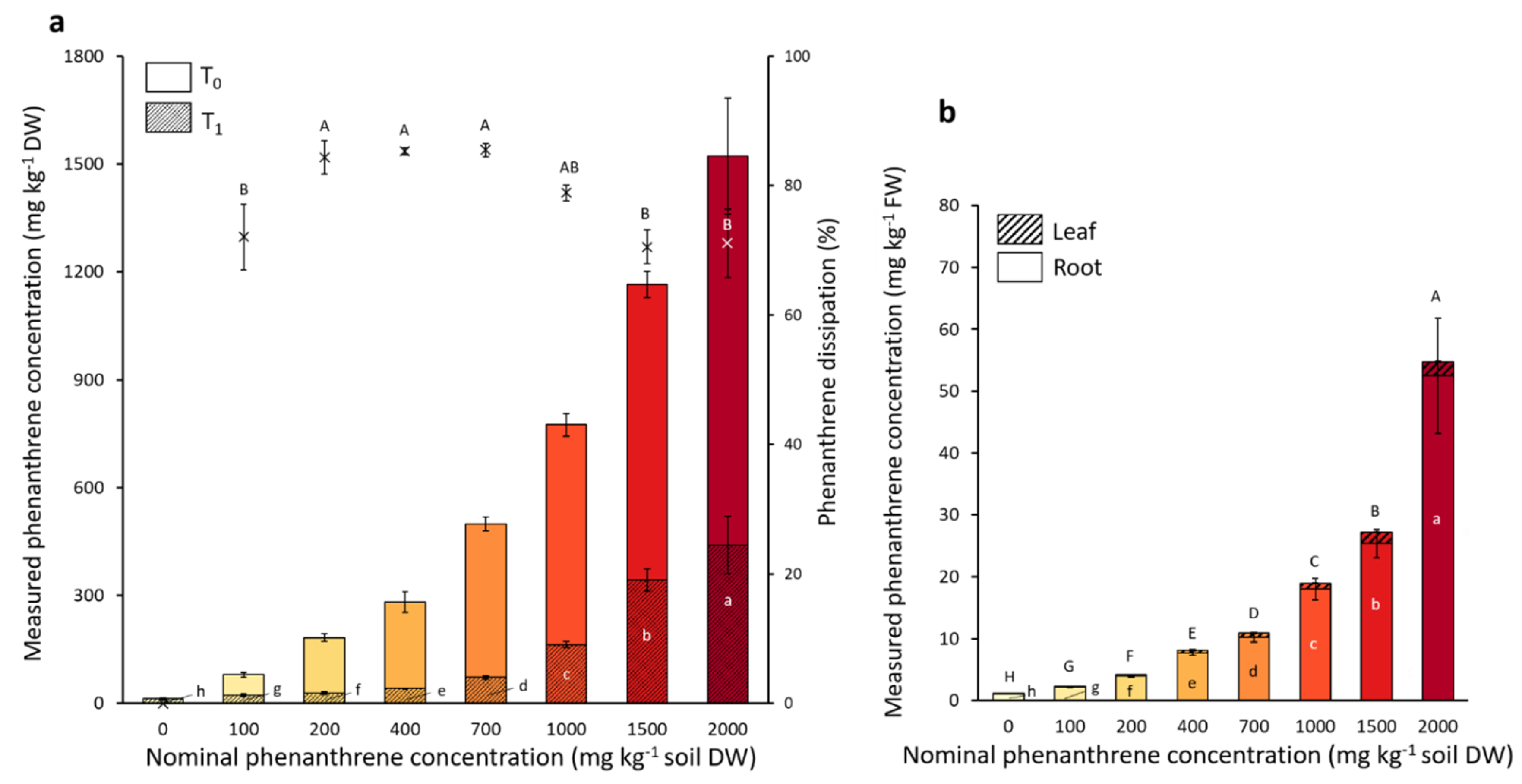
2.3. Phenanthrene Dissipation
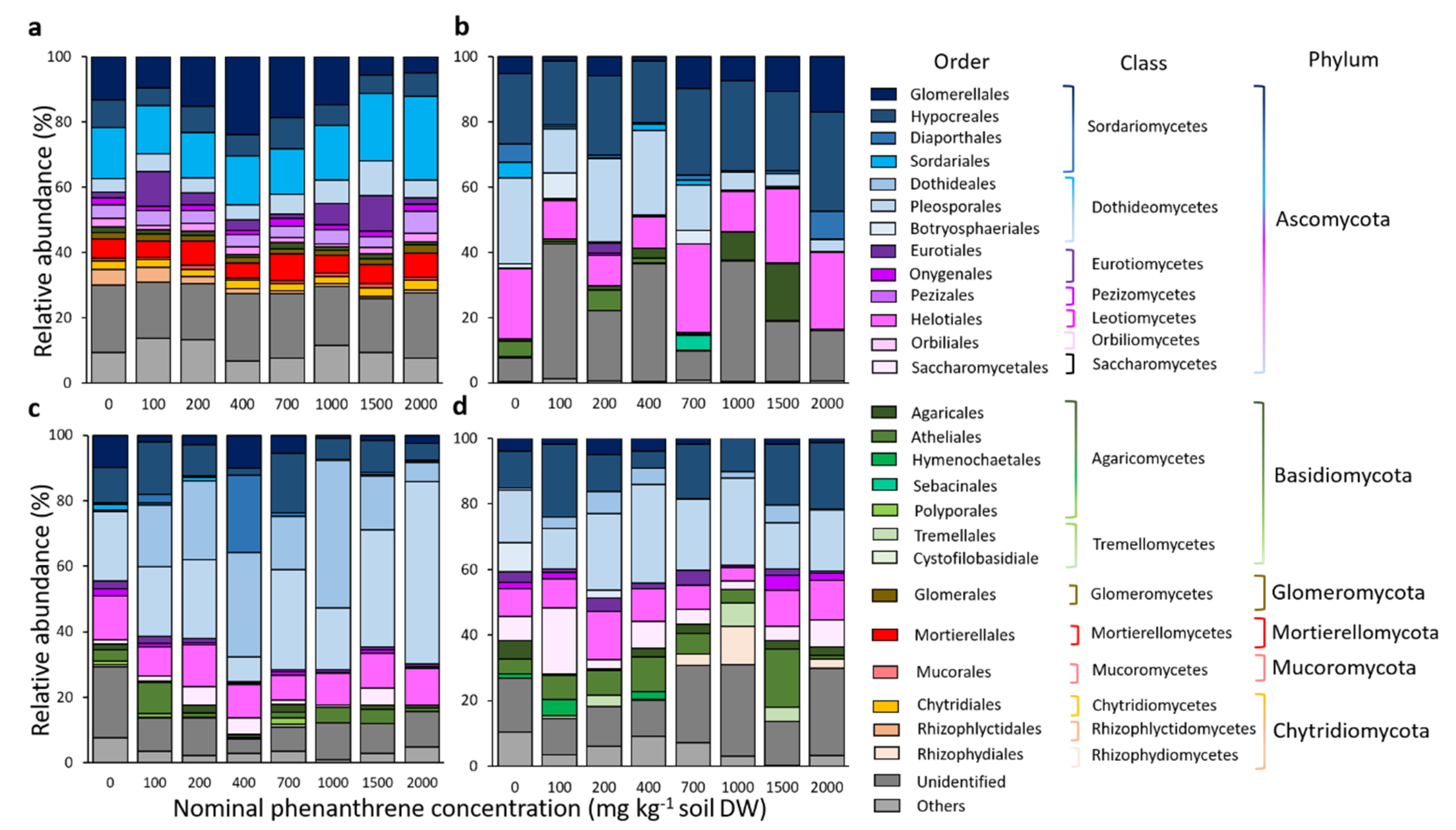
2.4. Fungal Community Diversity
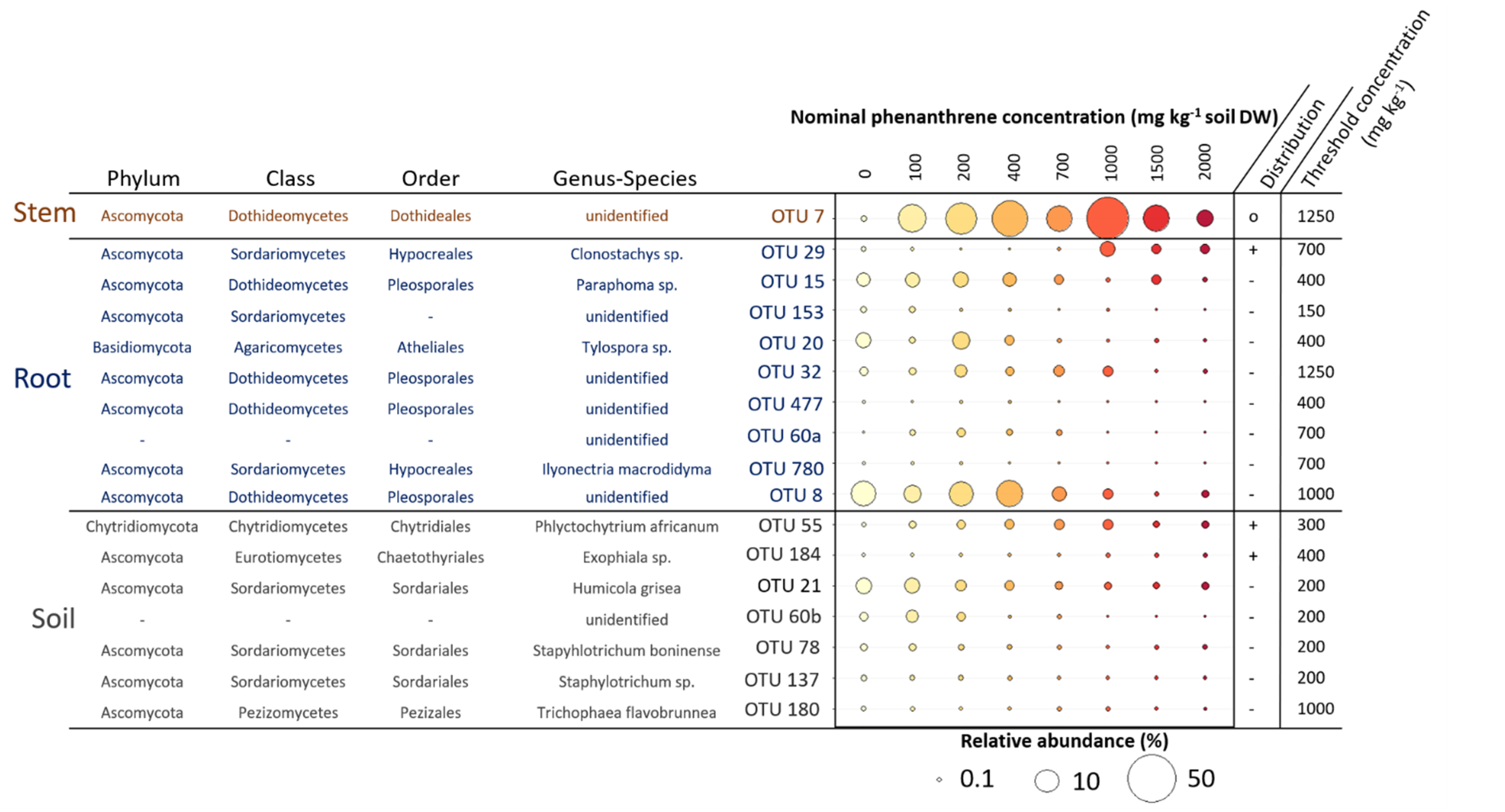
2.5. Indicator Species Linked to the PHE Contamination
3. Discussion
4. Material and Methods
4.1. Soil Sampling and Characteristics
4.2. Soil Spiking with Phenanthrene
4.3. Plant Pre-Culture and Microcosm Experiments
4.4. Fungal Colonization and Microscopy
4.5. Harvest of Soil, Root, Stem and Leaf Samples from the Second Experiment
4.6. Phenanthrene Analysis
4.7. DNA Extraction and ITS Sequencing
4.8. ITS Sequencing Data Analysis
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdel-Shafy, H.I.; Mansour, M.S.M. A Review on Polycyclic Aromatic Hydrocarbons: Source, Environmental Impact, Effect on Human Health and Remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Haritash, A.K.; Kaushik, C.P. Biodegradation Aspects of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.-H.; Xiong, A.-S.; Xue, Y.; Fu, X.-Y.; Gao, F.; Zhao, W.; Tian, Y.-S.; Yao, Q.-H. Microbial Biodegradation of Polyaromatic Hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilon-Smits, E. Phytoremediation. Annu. Rev. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef]
- Binet, P.; Portal, J.M.; Leyval, C. Fate of Polycyclic Aromatic Hydrocarbons (PAH) in the Rhizosphere and Mycorrhizosphere of Ryegrass. Plant Soil 2000, 7, 207–213. [Google Scholar] [CrossRef]
- Günther, T.; Sack, U.; Hofrichter, M.; Lätz, M. Oxidation of PAH and PAH-Derivatives by Fungal and Plant Oxidoreductases. J. Basic Microbiol. 1998, 38, 113–122. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, L. Plant Uptake, Accumulation and Translocation of Phenanthrene and Pyrene in Soils. Chemosphere 2004, 55, 1169–1178. [Google Scholar] [CrossRef]
- Liao, X.; Ma, X.; Yan, X.; Lin, L.; Shi, P.; Wu, Z. Transportation and Localization of Phenanthrene and Its Interaction with Different Species of Arsenic in Pteris Vittata L. Chemosphere 2016, 153, 307–314. [Google Scholar] [CrossRef]
- Sun, K.; Liu, J.; Jin, L.; Gao, Y. Utilizing Pyrene-Degrading Endophytic Bacteria to Reduce the Risk of Plant Pyrene Contamination. Plant Soil 2014, 374, 251–262. [Google Scholar] [CrossRef]
- Iffis, B.; St-Arnaud, M.; Hijri, M. Petroleum Contamination and Plant Identity Influence Soil and Root Microbial Communities While AMF Spores Retrieved from the Same Plants Possess Markedly Different Communities. Front. Plant Sci. 2017, 8, 1381. [Google Scholar] [CrossRef]
- Lemmel, F.; Maunoury-Danger, F.; Leyval, C.; Cébron, A. Altered Fungal Communities in Contaminated Soils from French Industrial Brownfields. J. Hazard. Mater. 2021, 406, 124296. [Google Scholar] [CrossRef] [PubMed]
- Bourdel, G.; Roy-Bolduc, A.; St-Arnaud, M.; Hijri, M. Concentration of Petroleum-Hydrocarbon Contamination Shapes Fungal Endophytic Community Structure in Plant Roots. Front. Microbiol. 2016, 7, 685. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, N.-X.; Yu, J.; Zhao, H.-M.; Cheng, Y.-T.; Mo, C.-H.; Cai, Q.-Y.; Li, Y.-W.; Li, H.; Wong, M.-H. Efficient Phytoremediation of Organic Contaminants in Soils Using Plant–Endophyte Partnerships. Sci. Total Environ. 2017, 583, 352–368. [Google Scholar] [CrossRef]
- Hassan, S.E.-D. Plant Growth-Promoting Activities for Bacterial and Fungal Endophytes Isolated from Medicinal Plant of Teucrium Polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef]
- Golparyan, F.; Azizi, A.; Soltani, J. Endophytes of Lippia Citriodora (Syn. Aloysia Triphylla) Enhance Its Growth and Antioxidant Activity. Eur. J. Plant Pathol. 2018, 152, 759–768. [Google Scholar] [CrossRef]
- Yung, L.; Sirguey, C.; Azou-Barré, A.; Blaudez, D. Natural Fungal Endophytes from Noccaea Caerulescens Mediate Neutral to Positive Effects on Plant Biomass, Mineral Nutrition and Zn Phytoextraction. Front. Microbiol. 2021, 12, 1726. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- He, W.; Megharaj, M.; Wu, C.-Y.; Subashchandrabose, S.R.; Dai, C.-C. Endophyte-Assisted Phytoremediation: Mechanisms and Current Application Strategies for Soil Mixed Pollutants. Crit. Rev. Biotechol. 2020, 40, 31–45. [Google Scholar] [CrossRef]
- Fu, W.-Q.; Xu, M.; Sun, K.; Chen, X.-L.; Dai, C.-C.; Jia, Y. Remediation Mechanism of Endophytic Fungus Phomopsis Liquidambaris on Phenanthrene in Vivo. Chemosphere 2020, 243, 125305. [Google Scholar] [CrossRef]
- Khan, Z.; Roman, D.; Kintz, T.; delas Alas, M.; Yap, R.; Doty, S. Degradation, Phytoprotection and Phytoremediation of Phenanthrene by Endophyte Pseudomonas Putida, PD1. Environ. Sci. Technol. 2014, 48, 12221–12228. [Google Scholar] [CrossRef] [PubMed]
- Dumas, A.-S.; Taconnat, L.; Barbas, E.; Rigaill, G.; Catrice, O.; Bernard, D.; Benamar, A.; Macherel, D.; El Amrani, A.; Berthomé, R. Unraveling the Early Molecular and Physiological Mechanisms Involved in Response to Phenanthrene Exposure. BMC Genom. 2016, 17, 818. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Liu, J.; Gao, Y.; Jin, L.; Gu, Y.; Wang, W. Isolation, Plant Colonization Potential and Phenanthrene Degradation Performance of the Endophytic Bacterium Pseudomonas Sp. Ph6-Gfp. Sci. Rep. 2015, 4, 5462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, F.; Gainza, F.; Pérez, R.; Zamudio, F. Phytoremediation of Heavy Metals using Poplars (Populus spp.): A Glimpse of the Plant Responses to Copper, Cadmium and Zinc Stress. In Handbook of Phytoremediation; Nova Science: New York, NY, USA, 2011; Volume 28, pp. 387–413. [Google Scholar]
- Gottel, N.R.; Castro, H.F.; Kerley, M.; Yang, Z.; Pelletier, D.A.; Podar, M.; Karpinets, T.; Uberbacher, E.; Tuskan, G.A.; Vilgalys, R.; et al. Distinct Microbial Communities within the Endosphere and Rhizosphere of Populus Deltoides Roots across Contrasting Soil Types. Appl. Environ. Microbiol. 2011, 77, 5934–5944. [Google Scholar] [CrossRef] [Green Version]
- Beckers, B.; Op De Beeck, M.; Weyens, N.; Boerjan, W.; Vangronsveld, J. Structural Variability and Niche Differentiation in the Rhizosphere and Endosphere Bacterial Microbiome of Field-Grown Poplar Trees. Microbiome 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Durand, A.; Maillard, F.; Alvarez-Lopez, V.; Guinchard, S.; Bertheau, C.; Valot, B.; Blaudez, D.; Chalot, M. Bacterial Diversity Associated with Poplar Trees Grown on a Hg-Contaminated Site: Community Characterization and Isolation of Hg-Resistant Plant Growth-Promoting Bacteria. Sci. Total Environ. 2018, 622–623, 1165–1177. [Google Scholar] [CrossRef] [Green Version]
- Barra Caracciolo, A.; Grenni, P.; Garbini, G.L.; Rolando, L.; Campanale, C.; Aimola, G.; Fernandez-Lopez, M.; Fernandez-Gonzalez, A.J.; Villadas, P.J.; Ancona, V. Characterization of the Belowground Microbial Community in a Poplar-Phytoremediation Strategy of a Multi-Contaminated Soil. Front. Microbiol. 2020, 11, 2073. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Tardif, S.; Yergeau, É.; Tremblay, J.; Legendre, P.; Whyte, L.G.; Greer, C.W. The Willow Microbiome Is Influenced by Soil Petroleum-Hydrocarbon Concentration with Plant Compartment-Specific Effects. Front. Microbiol. 2016, 7, 1363. [Google Scholar] [CrossRef] [Green Version]
- Alkio, M.; Tabuchi, T.M.; Wang, X.; Colón-Carmona, A. Stress Responses to Polycyclic Aromatic Hydrocarbons in Arabidopsis Include Growth Inhibition and Hypersensitive Response-like Symptoms. J. Exp. Bot. 2005, 56, 2983–2994. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Weisman, D.; Ye, Y.; Cui, B.; Huang, Y.; Colón-Carmona, A.; Wang, Z. An Oxidative Stress Response to Polycyclic Aromatic Hydrocarbon Exposure Is Rapid and Complex in Arabidopsis Thaliana. Plant Sci. 2009, 176, 375–382. [Google Scholar] [CrossRef]
- Wei, H.; Song, S.; Tian, H.; Liu, T. Effects of Phenanthrene on Seed Germination and Some Physiological Activities of Wheat Seedling. Comptes Rendus Biol. 2014, 337, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, J.; Leglize, P.; Vincent, Q.; Zelko, I.; Mustin, C.; Ouvrard, S.; Sterckeman, T. Effect and Localization of Phenanthrene in Maize Roots. Chemosphere 2016, 149, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; El-Din Hassan, S.; Lauron-Moreau, A.; Al-Otaibi, F.; Hijri, M.; Yergeau, E.; St-Arnaud, M. Linkage between Bacterial and Fungal Rhizosphere Communities in Hydrocarbon-Contaminated Soils Is Related to Plant Phylogeny. ISME J. 2014, 8, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Törneman, N.; Yang, X.; Bååth, E.; Bengtsson, G. Spatial Covariation of Microbial Community Composition and Polycyclic Aromatic Hydrocarbon Concentration in A Creosote-Polluted Soil. Environ. Toxicol. Chem. 2008, 27, 1039. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Oszust, K. Functional Diversity of Fungal Communities in Soil Contaminated with Diesel Oil. Front. Microbiol. 2017, 8, 1862. [Google Scholar] [CrossRef]
- Zhou, Z.-F.; Wang, M.-X.; Zuo, X.-H.; Yao, Y.-H. Comparative Investigation of Bacterial, Fungal, and Archaeal Community Structures in Soils in a Typical Oilfield in Jianghan, China. Arch. Environ. Contam. Toxicol. 2017, 72, 65–77. [Google Scholar] [CrossRef]
- Guiraud, P.; Villemain, D.; Kadri, M.; Bordjiba, O.; Steiman, R. Biodegradation Capability of Absidia Fusca Linnemann towards Environmental Pollutants. Chemosphere 2003, 52, 663–671. [Google Scholar] [CrossRef]
- Benguenab, A.; Chibani, A. Biodegradation of Petroleum Hydrocarbons by Filamentous Fungi (Aspergillus Ustus and Purpureocillium Lilacinum) Isolated from Used Engine Oil Contaminated Soil. Acta Ecol. Sin. 2021, 41, 416–423. [Google Scholar] [CrossRef]
- Soldi, E.; Casey, C.; Murphy, B.R.; Hodkinson, T.R. Fungal Endophytes for Grass Based Bioremediation: An Endophytic Consortium Isolated from Agrostis Stolonifera Stimulates the Growth of Festuca Arundinacea in Lead Contaminated Soil. J. Fungi 2020, 6, 254. [Google Scholar] [CrossRef] [PubMed]
- Hamad, A.A.; Moubasher, H.A.; Moustafa, Y.M.; Mohamed, N.H.; Abd-el rhim, E.H. Petroleum Hydrocarbon Bioremediation Using Native Fungal Isolates and Consortia. Sci. World J. 2021, 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zafra, G.; Cortés-Espinosa, D.V. Biodegradation of Polycyclic Aromatic Hydrocarbons by Trichoderma Species: A Mini Review. Environ. Sci. Pollut. Res. 2015, 22, 19426–19433. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.; Cesanelli, I.; Diánez, F.; Sánchez-Montesinos, B.; Moreno-Gavíra, A. Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses. J. Fungi 2021, 7, 939. [Google Scholar] [CrossRef]
- Isola, D.; Scano, A.; Orrù, G.; Prenafeta-Boldú, F.X.; Zucconi, L. Hydrocarbon-Contaminated Sites: Is There Something More Than Exophiala Xenobiotica? New Insights into Black Fungal Diversity Using the Long Cold Incubation Method. J. Fungi 2021, 7, 817. [Google Scholar] [CrossRef]
- Marín, F.; Navarrete, H.; Narvaez-Trujillo, A. Total Petroleum Hydrocarbon Degradation by Endophytic Fungi from the Ecuadorian Amazon. Adv. Microbiol. 2018, 8, 1029–1053. [Google Scholar] [CrossRef] [Green Version]
- Riser-Roberts, E. Biodegradation/Mineralization/Biotransformation/Bioaccumulation of Petroleum Constituents and Associated Heavy Metals. In Remediation of Petroleum Contaminated Soils; CRC Press: Boca Raton, FL, USA, 1998; pp. 115–178. [Google Scholar]
- Perfect, S.E.; Hughes, H.B.; O’Connell, R.J.; Green, J.R. Colletotrichum: A Model Genus for Studies on Pathology and Fungal–Plant Interactions. Fungal Genet. Biol. 1999, 27, 186–198. [Google Scholar] [CrossRef]
- Kia, S.H.; Jurkechova, M.; Glynou, K.; Piepenbring, M.; Maciá-Vicente, J.G. The Effects of Fungal Root Endophytes on Plant Growth Are Stable along Gradients of Abiotic Habitat Conditions. FEMS Microbiol. Ecol. 2018, 94, fix162. [Google Scholar] [CrossRef]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Zhang, X.; Shi, W.; Wang, H.; Yu, D. Biocontrol Potential of Endophytic Fungi in Medicinal Plants from Wuhan Botanical Garden in China. Biol. Control 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Veach, A.M.; Yip, D.; Engle, N.L.; Yang, Z.K.; Bible, A.; Morrell-Falvey, J.; Tschaplinski, T.J.; Kalluri, U.C.; Schadt, C.W. Modification of Plant Cell Wall Chemistry Impacts Metabolome and Microbiome Composition in Populus PdKOR1 RNAi Plants. Plant Soil 2018, 429, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Baron, N.C.; Pagnocca, F.C.; Otsuka, A.A.; Prenafeta-Boldú, F.X.; Vicente, V.A.; Attili de Angelis, D. Black Fungi and Hydrocarbons: An Environmental Survey for Alkylbenzene Assimilation. Microorganisms 2021, 9, 1008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yao, Y.-F. Endophytic Fungal Communities Associated with Vascular Plants in the High Arctic Zone Are Highly Diverse and Host-Plant Specific. PLoS ONE 2015, 10, e0130051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.C.; Dombrowski, J.E. Isolation and Identification of Fungal Endophytes from Grasses along the Oregon Coast. Am. J. Political Sci. 2015, 6, 3216–3230. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, S.; Zalar, P.; van den Ende, B.G.; Gunde-Cimerman, N. Relation of Halotolerance to Human Pathogenicity in the Fungal Tree of Life: An Overview of Ecology and Evolution under Stress. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya, Cellular Origin, Life in Extreme Habitats and Astrobiology; Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 9, pp. 372–395. [Google Scholar]
- Trouvelot, A.; Kough, J.; Gianinazzi-Pearson, V. Mesure Du Taux de Mycorhization VA d’un Système Radiculaire. In Recherches et Méthodes Avancées Ayant une Signifcation Fonctionnelle; Gianinazzi, S., Gininazzi-Pearson, V., Eds.; INRA: Paris, France, 1986; pp. 217–220. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Yourstone, S.; Mieczkowski, P.; Jones, C.D.; Dangl, J.L. Practical Innovations for High-Throughput Amplicon Sequencing. Nat. Methods 2013, 10, 999–1002. [Google Scholar] [CrossRef]
- Cregger, M.A.; Veach, A.M.; Yang, Z.K.; Crouch, M.J.; Vilgalys, R.; Tuskan, G.A.; Schadt, C.W. The Populus Holobiont: Dissecting the Effects of Plant Niches and Genotype on the Microbiome. Microbiome 2018, 6, 31. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Pascal, G.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; et al. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial. R Package Version 2011, 1, 1–43. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Põlme, S.; Abarenkov, K.; Henrik Nilsson, R.; Lindahl, B.D.; Tedersoo, L.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; et al. FungalTraits: A User-Friendly Traits Database of Fungi and Fungus-like Stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.4. 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 15 October 2021).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. 2021. Available online: https://cran.r-project.org/web/packages/corrplot/vignettes/corrplot-intro.html (accessed on 3 December 2021).
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An Interactive Venn Diagram Viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.E.; King, R.S. A New Method for Detecting and Interpreting Biodiversity and Ecological Community Thresholds: Threshold Indicator Taxa ANalysis (TITAN). Methods Ecol. Evol. 2010, 1, 25–37. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gréau, L.; Blaudez, D.; Heintz, D.; Zumsteg, J.; Billet, D.; Cébron, A. Response of Poplar and Associated Fungal Endophytic Communities to a PAH Contamination Gradient. Int. J. Mol. Sci. 2022, 23, 5909. https://doi.org/10.3390/ijms23115909
Gréau L, Blaudez D, Heintz D, Zumsteg J, Billet D, Cébron A. Response of Poplar and Associated Fungal Endophytic Communities to a PAH Contamination Gradient. International Journal of Molecular Sciences. 2022; 23(11):5909. https://doi.org/10.3390/ijms23115909
Chicago/Turabian StyleGréau, Lilian, Damien Blaudez, Dimitri Heintz, Julie Zumsteg, David Billet, and Aurélie Cébron. 2022. "Response of Poplar and Associated Fungal Endophytic Communities to a PAH Contamination Gradient" International Journal of Molecular Sciences 23, no. 11: 5909. https://doi.org/10.3390/ijms23115909