
Fluctuations in Corneal Endothelial LAP2 Expression Levels Correlate with Passage Dependent Declines in Their Cell Proliferative Activity

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Corneal Endothelial Cell Culture
2.3. Quantitative Real Time (RT)-PCR
2.4. Immunofluorescence (IF)
2.5. Growth Factors Treatment
2.6. Western Blot
3. Results
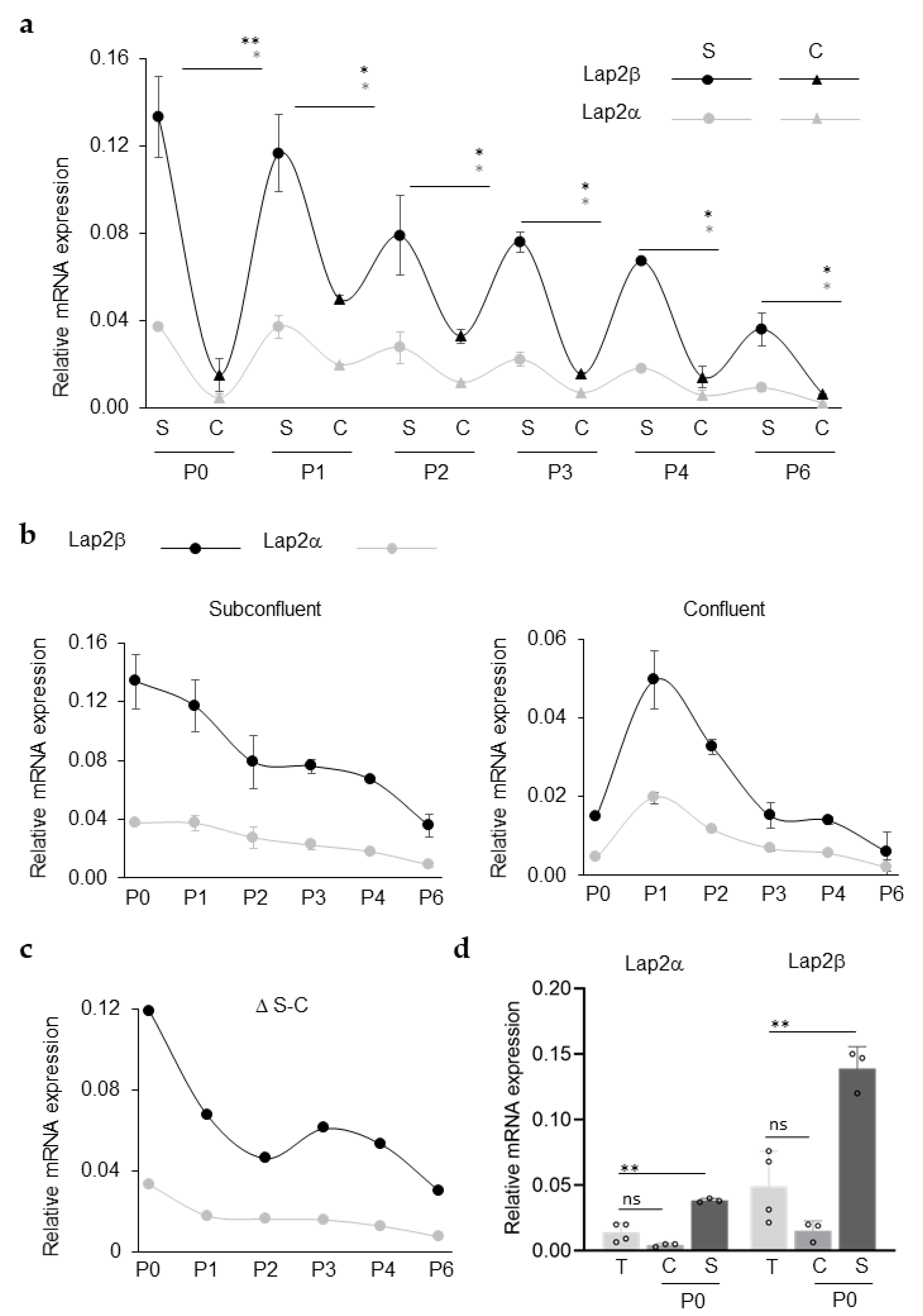
3.1. LAP2 RNA Expression in Primary rCEnCs
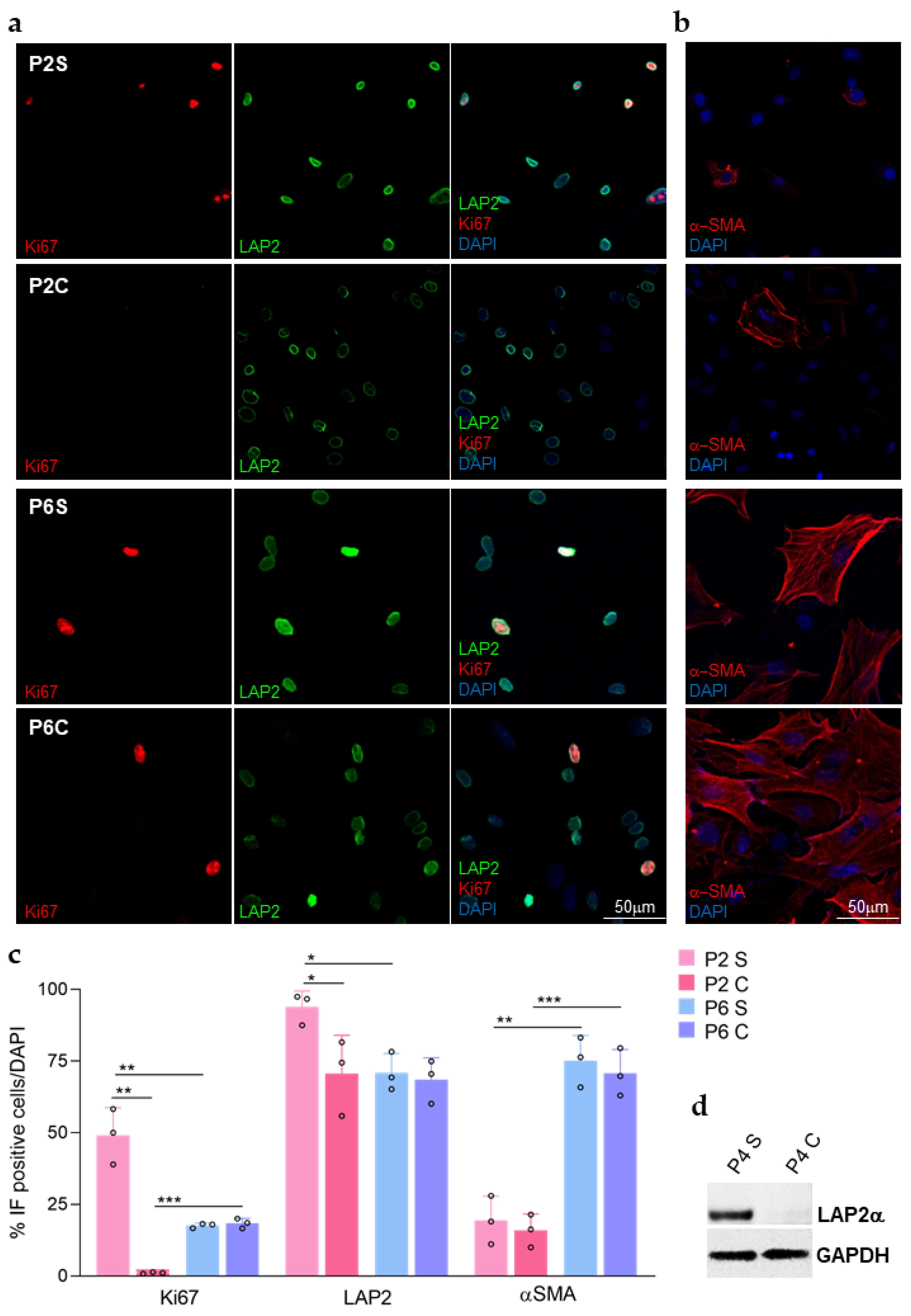
3.2. LAP2 and ki67 Protein Expression in Primary rCEnCs
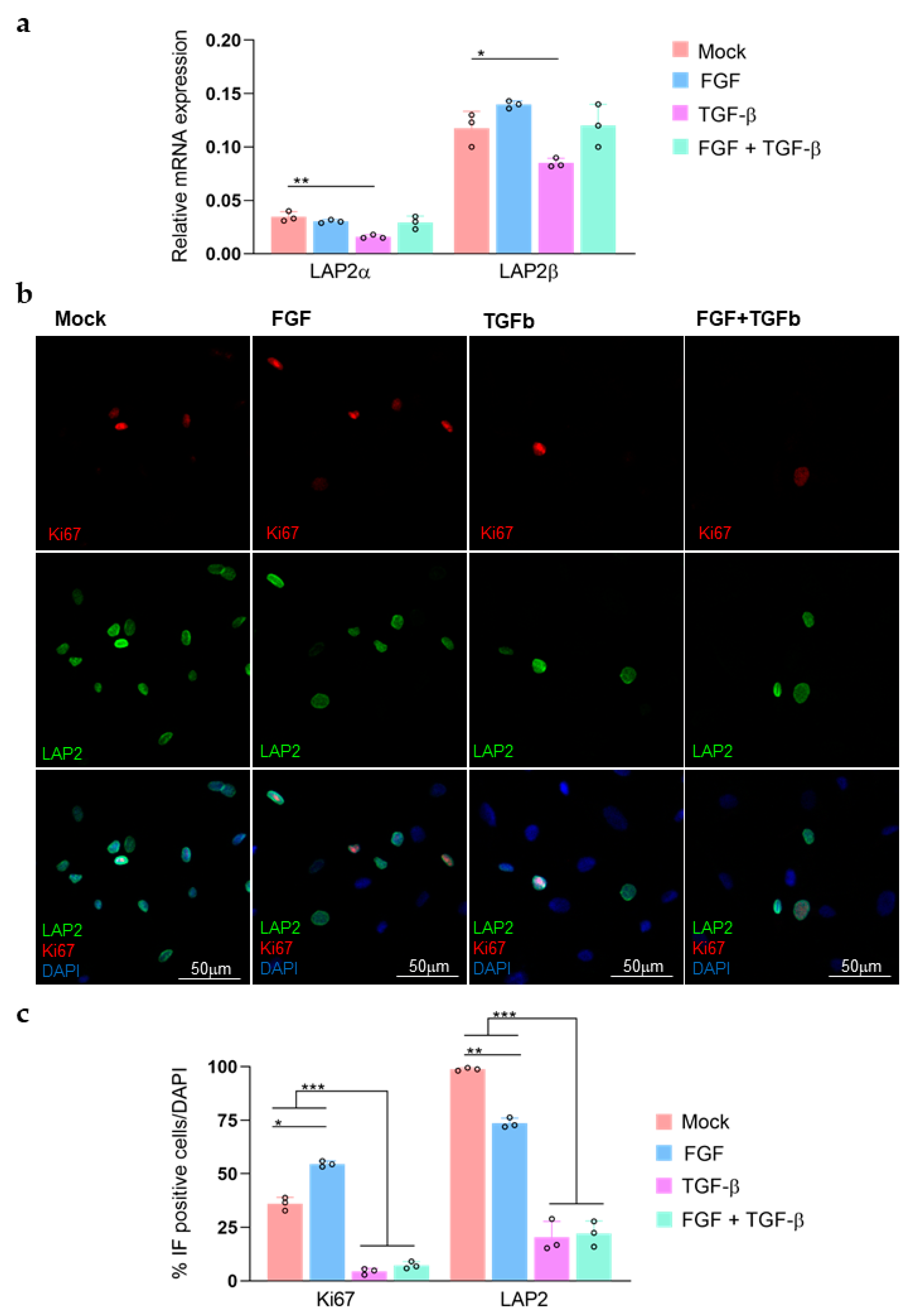
3.3. TGFβ-FGF Treatment in Primary rCEnCs
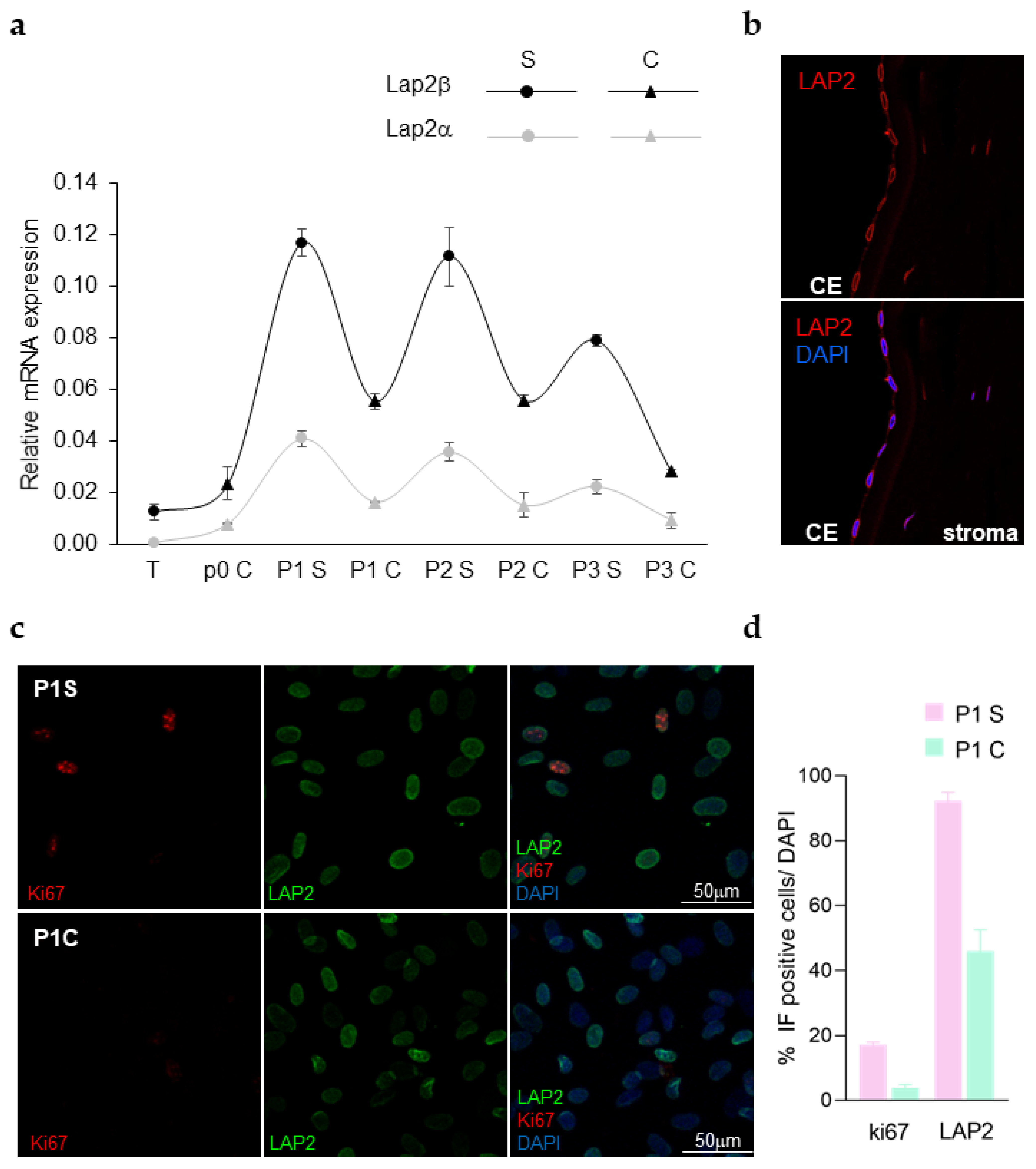
3.4. Primary Human (H)CEnCs and LAP2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Català, P.; Thuret, G.; Skottman, H.; Mehta, J.S.; Parekh, M.; Dhubhghaill, S.N.; Collin, R.W.; Nuijts, R.M.; Ferrari, S.; LaPointe, V.L.; et al. Approaches for corneal endothelium regenerative medicine. Prog. Retin. Eye Res. 2021, 87, 100987. [Google Scholar] [CrossRef] [PubMed]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, G.; Ardigò, D.; Milazzo, G.; Iotti, G.; Guatelli, P.; Pelosi, D.; De Luca, M. Navigating market authorization: The path holoclar took to become the first stem cell product approved in the European Union. Stem Cells Transl. Med. 2018, 7, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuft, S.; Coster, D. The corneal endothelium. Eye 1990, 4, 389–424. [Google Scholar] [CrossRef]
- Peh, G.S.; Chng, Z.; Ang, H.-P.; Cheng, T.Y.; Adnan, K.; Seah, X.-Y.; George, B.L.; Toh, K.-P.; Tan, D.T.; Yam, G.H.; et al. Propagation of human corneal endothelial cells: A novel dual media approach. Cell Transplant. 2015, 24, 287–304. [Google Scholar] [CrossRef]
- Okumura, N.; Kakutani, K.; Numata, R.; Nakahara, M.; Schlötzer-Schrehardt, U.; Kruse, F.; Kinoshita, S.; Koizumi, N. Laminin-511 and-521 enable efficient in vitro expansion of human corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2933–2942. [Google Scholar] [CrossRef] [Green Version]
- Nakahara, M.; Okumura, N.; Kay, E.P.; Hagiya, M.; Imagawa, K.; Hosoda, Y.; Kinoshita, S.; Koizumi, N. Corneal endothelial expansion promoted by human bone marrow mesenchymal stem cell-derived conditioned medium. PLoS ONE 2013, 8, e69009. [Google Scholar] [CrossRef]
- Kinoshita, S.; Koizumi, N.; Ueno, M.; Okumura, N.; Imai, K.; Tanaka, H.; Yamamoto, Y.; Nakamura, T.; Inatomi, T.; Bush, J. Injection of cultured cells with a ROCK inhibitor for bullous keratopathy. N. Engl. J. Med. 2018, 378, 995–1003. [Google Scholar] [CrossRef]
- Frausto, R.F.; Swamy, V.S.; Peh, G.S.; Boere, P.M.; Hanser, E.M.; Chung, D.; George, B.L.; Morselli, M.; Kao, L.; Azimov, R.; et al. Phenotypic and functional characterization of corneal endothelial cells during in vitro expansion. Sci. Rep. 2020, 10, 1–22. [Google Scholar] [CrossRef]
- Yamashita, K.; Hatou, S.; Inagaki, E.; Higa, K.; Tsubota, K.; Shimmura, S. A rabbit corneal endothelial dysfunction model using endothelial-mesenchymal transformed cells. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Okumura, N.; Hirano, H.; Numata, R.; Nakahara, M.; Ueno, M.; Hamuro, J.; Kinoshita, S.; Koizumi, N. Cell surface markers of functional phenotypic corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7610–7618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bogerd, B.; Zakaria, N.; Adam, B.; Matthyssen, S.; Koppen, C.; Dhubhghaill, S.N. Corneal endothelial cells over the past decade: Are we missing the mark (er)? Transl. Vis. Sci. Technol. 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naetar, N.; Korbei, B.; Kozlov, S.; Kerenyi, M.A.; Dorner, D.; Kral, R.; Gotic, I.; Fuchs, P.; Cohen, T.V.; Bittner, R.; et al. Loss of nucleoplasmic LAP2α–lamin A complexes causes erythroid and epidermal progenitor hyperproliferation. Nat. Cell Biol. 2008, 10, 1341–1348. [Google Scholar] [CrossRef]
- Dreesen, O.; Chojnowski, A.; Ong, P.F.; Zhao, T.Y.; Common, J.E.; Lunny, D.; Lane, E.B.; Lee, S.J.; Vardy, L.A.; Stewart, C.L.; et al. Lamin B1 fluctuations have differential effects on cellular proliferation and senescence. J. Cell Biol. 2013, 200, 605–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delfarah, A.; Parrish, S.; Junge, J.A.; Yang, J.; Seo, F.; Li, S.; Mac, J.; Wang, P.; Fraser, S.E.; Graham, N.A. Inhibition of nucleotide synthesis promotes replicative senescence of human mammary epithelial cells. J. Biol. Chem. 2019, 294, 10564–10578. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.T.; Nair, R.R.; Poll, B.G.; Pineda, C.M.; Hobbs, R.P.; Matunis, M.J.; Coulombe, P.A. Keratin 17 regulates nuclear morphology and chromatin organization. J. Cell Sci. 2020, 133, jcs254094. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Sansonetti, P.J. Quantitative RT-PCR profiling of the rabbit immune response: Assessment of acute Shigella flexneri infection. PLoS ONE 2012, 7, e36446. [Google Scholar] [CrossRef] [Green Version]
- Maurizi, E.; Schiroli, D.; Zini, R.; Limongelli, A.; Mistò, R.; Macaluso, C.; Pellegrini, G. A fine-tuned β-catenin regulation during proliferation of corneal endothelial cells revealed using proteomics analysis. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Mirza, A.N.; McKellar, S.A.; Urman, N.M.; Brown, A.S.; Hollmig, T.; Aasi, S.Z.; Oro, A.E. LAP2 proteins chaperone GLI1 movement between the lamina and chromatin to regulate transcription. Cell 2019, 176, 198–212.e115. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Lu, Z.; Reinach, P.; Zhang, J.; Dai, W.; Lu, L.; Xu, M. TGF-β2 inhibits AKT activation and FGF-2-induced corneal endothelial cell proliferation. Exp. Cell Res. 2006, 312, 3631–3640. [Google Scholar] [CrossRef]
- Naetar, N.; Georgiou, K.; Knapp, C.; Bronshtein, I.; Zier, E.; Fichtinger, P.; Dechat, T.; Garini, Y.; Foisner, R. LAP2alpha maintains a mobile and low assembly state of A-type lamins in the nuclear interior. Elife 2021, 10, e63476. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.; Theodor, L.; Shoham, J.; Gokkel, E.; Brok-Simoni, F.; Avraham, K.B.; Copeland, N.G.; Jenkins, N.A.; Rechavi, G.; Simon, A.J. The characterization and localization of the mouse thymopoietin/lamina-associated polypeptide 2 gene and its alternatively spliced products. Genome Res. 1996, 6, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gant, T.M.; Harris, C.A.; Wilson, K.L. Roles of LAP2 proteins in nuclear assembly and DNA replication: Truncated LAP2β proteins alter lamina assembly, envelope formation, nuclear size, and DNA replication efficiency in Xenopus laevis extracts. J. Cell Biol. 1999, 144, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishijima, Y.; Toda, T.; Matsushita, H.; Yoshida, M.; Kimura, N. Expression of thymopoietin β/lamina-associated polypeptide 2 (TPβ/LAP2) and its family proteins as revealed by specific antibody induced against recombinant human thymopoietin. Biochem. Biophys. Res. Commun. 1996, 226, 431–438. [Google Scholar] [CrossRef]
- Vlcek, S.; Foisner, R. Lamins and lamin-associated proteins in aging and disease. Curr. Opin. Cell Biol. 2007, 19, 298–304. [Google Scholar] [CrossRef]
- Parise, P.; Finocchiaro, G.; Masciadri, B.; Quarto, M.; Francois, S.; Mancuso, F.; Müller, H. Lap2α expression is controlled by E2F and deregulated in various human tumors. Cell Cycle 2006, 5, 1331–1341. [Google Scholar] [CrossRef]
- Sobecki, M.; Mrouj, K.; Colinge, J.; Gerbe, F.; Jay, P.; Krasinska, L.; Dulic, V.; Fisher, D. Cell-cycle regulation accounts for variability in Ki-67 expression levels. Cancer Res. 2017, 77, 2722–2734. [Google Scholar] [CrossRef] [Green Version]
- Vlcek, S.; Just, H.; Dechat, T.; Foisner, R. Functional diversity of LAP2α and LAP2β in postmitotic chromosome association is caused by an α-specific nuclear targeting domain. EMBO J. 1999, 18, 6370–6384. [Google Scholar] [CrossRef] [Green Version]
- Pekovic, V.; Harborth, J.; Broers, J.L.; Ramaekers, F.C.; van Engelen, B.; Lammens, M.; von Zglinicki, T.; Foisner, R.; Hutchison, C.; Markiewicz, E. Nucleoplasmic LAP2α–lamin A complexes are required to maintain a proliferative state in human fibroblasts. J. Cell Biol. 2007, 176, 163–172. [Google Scholar] [CrossRef]
- Harris, D.L.; Joyce, N.C. Transforming growth factor-beta suppresses proliferation of rabbit corneal endothelial cells in vitro. J. Interferon Cytokine Res. 1999, 19, 327–334. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, W.-I.; Smith, R.E.; Kay, E.P. Role of p27Kip1 in cAMP-and TGF-β2–Mediated Antiproliferation in Rabbit Corneal Endothelial Cells. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3142–3149. [Google Scholar]
- Zhu, Y.-T.; Chen, H.-C.; Chen, S.-Y.; Tseng, S.C. Nuclear p120 catenin unlocks mitotic block of contact-inhibited human corneal endothelial monolayers without disrupting adherent junctions. J. Cell Sci. 2012, 125, 3636–3648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, N.C. Cell cycle status in human corneal endothelium. Exp. Eye Res. 2005, 81, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T. Lamin A-dependent nuclear defects in human aging. Science 2006, 312, 1059–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.G.; Ko, M.K.; Kay, E.P. Endothelial mesenchymal transformation mediated by IL-1β-induced FGF-2 in corneal endothelial cells. Exp. Eye Res. 2012, 95, 35–39. [Google Scholar] [CrossRef]
- Lee, J.G.; Jung, E.; Heur, M. Fibroblast growth factor 2 induces proliferation and fibrosis via SNAI1-mediated activation of CDK2 and ZEB1 in corneal endothelium. J. Biol. Chem. 2018, 293, 3758–3769. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Specifications | Sequence (5′ to 3′) | Size (bp) |
|---|---|---|---|
| LAP2α and β | Forward_rabbit and Human | ATT GTG GGA ACA ACC AGG AA | |
| LAP2α | Reverse_rabbit | CCC TAG TGG ACT TCA CTT TCT | 205 |
| Reverse_Human | CCA CCA GAG GGA GTA GTT C | 248 | |
| LAP2β | Reverse_rabbit | CCC TTT AGC GGT TCT CTC T | 212 |
| Reverse_Human | TTT GCT CTG CCC TTT AGT GG | 221 | |
| GAPDH | Forward_rabbit | TGA CGA CAT CAA GAA GGT GGT G | 120 [17] |
| Reverse_rabbit | GAA GGT GGA GGA GTG GGT GTC | ||
| Forward_ Human | GTC TCC TCT GAC TTC AAC AGC G | 131 | |
| Reverse_ Human | ACC ACC CTG TTG CTG TAG CCA A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurizi, E.; Merra, A.; Schiroli, D.; Ghezzi, B.; Macaluso, C.; Pellegrini, G. Fluctuations in Corneal Endothelial LAP2 Expression Levels Correlate with Passage Dependent Declines in Their Cell Proliferative Activity. Int. J. Mol. Sci. 2022, 23, 5859. https://doi.org/10.3390/ijms23105859
Maurizi E, Merra A, Schiroli D, Ghezzi B, Macaluso C, Pellegrini G. Fluctuations in Corneal Endothelial LAP2 Expression Levels Correlate with Passage Dependent Declines in Their Cell Proliferative Activity. International Journal of Molecular Sciences. 2022; 23(10):5859. https://doi.org/10.3390/ijms23105859
Chicago/Turabian StyleMaurizi, Eleonora, Alessia Merra, Davide Schiroli, Benedetta Ghezzi, Claudio Macaluso, and Graziella Pellegrini. 2022. "Fluctuations in Corneal Endothelial LAP2 Expression Levels Correlate with Passage Dependent Declines in Their Cell Proliferative Activity" International Journal of Molecular Sciences 23, no. 10: 5859. https://doi.org/10.3390/ijms23105859