
Genome-Wide Identification and Expression Analyses of the Aquaporin Gene Family in Passion Fruit (Passiflora edulis), Revealing PeTIP3-2 to Be Involved in Drought Stress

Abstract
:1. Introduction
2. Results
2.1. Identification of the Passion Fruit AQP Family
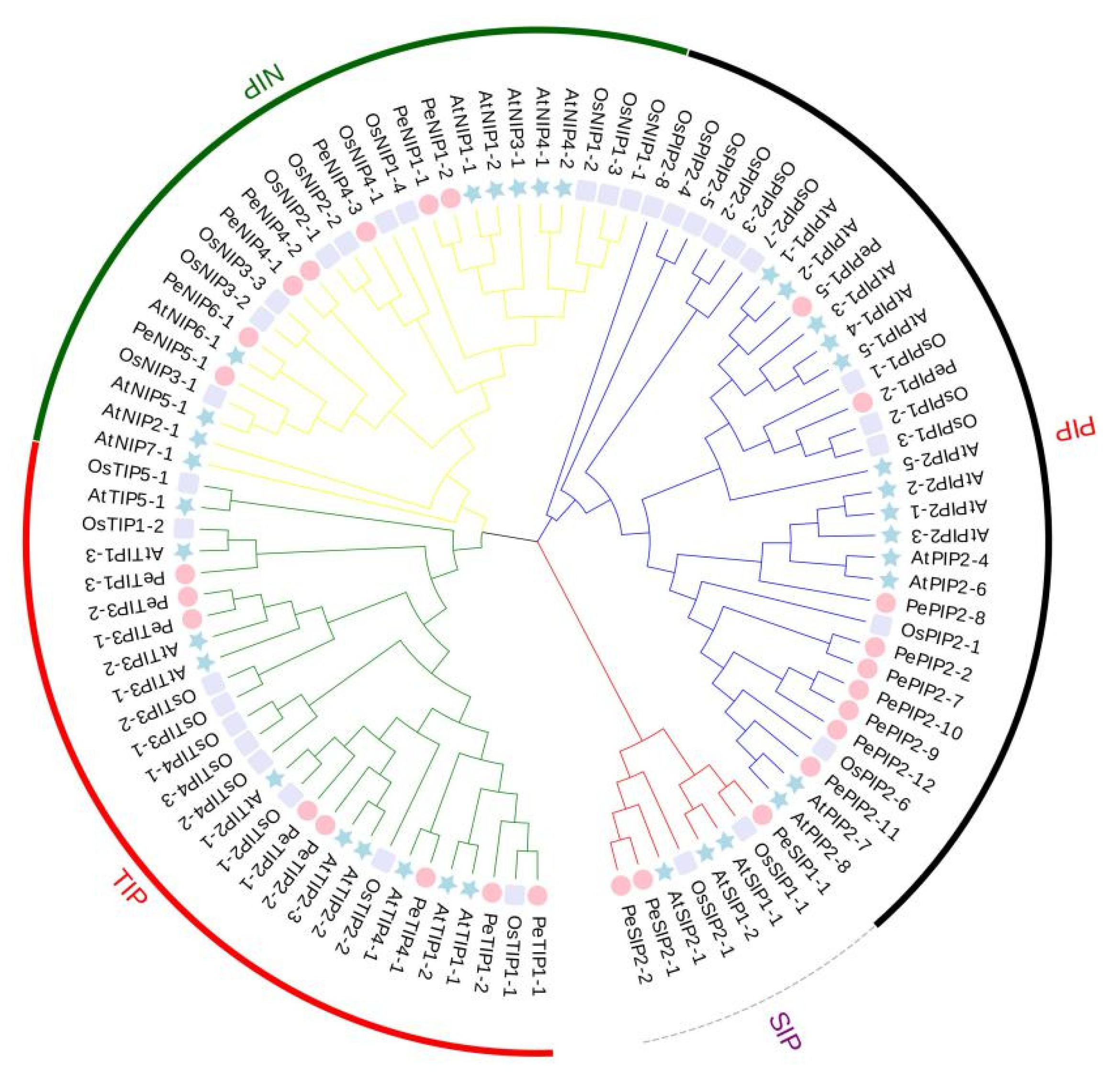
2.2. Evolutionary Characterization of AQP Genes
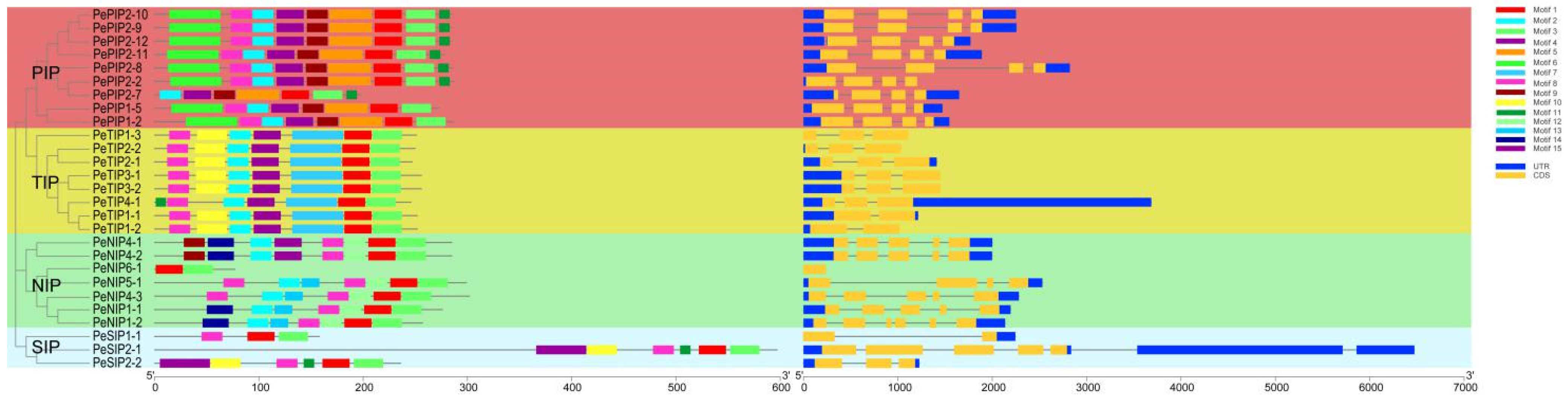
2.3. Gene Structure and Conserved Motif Analysis
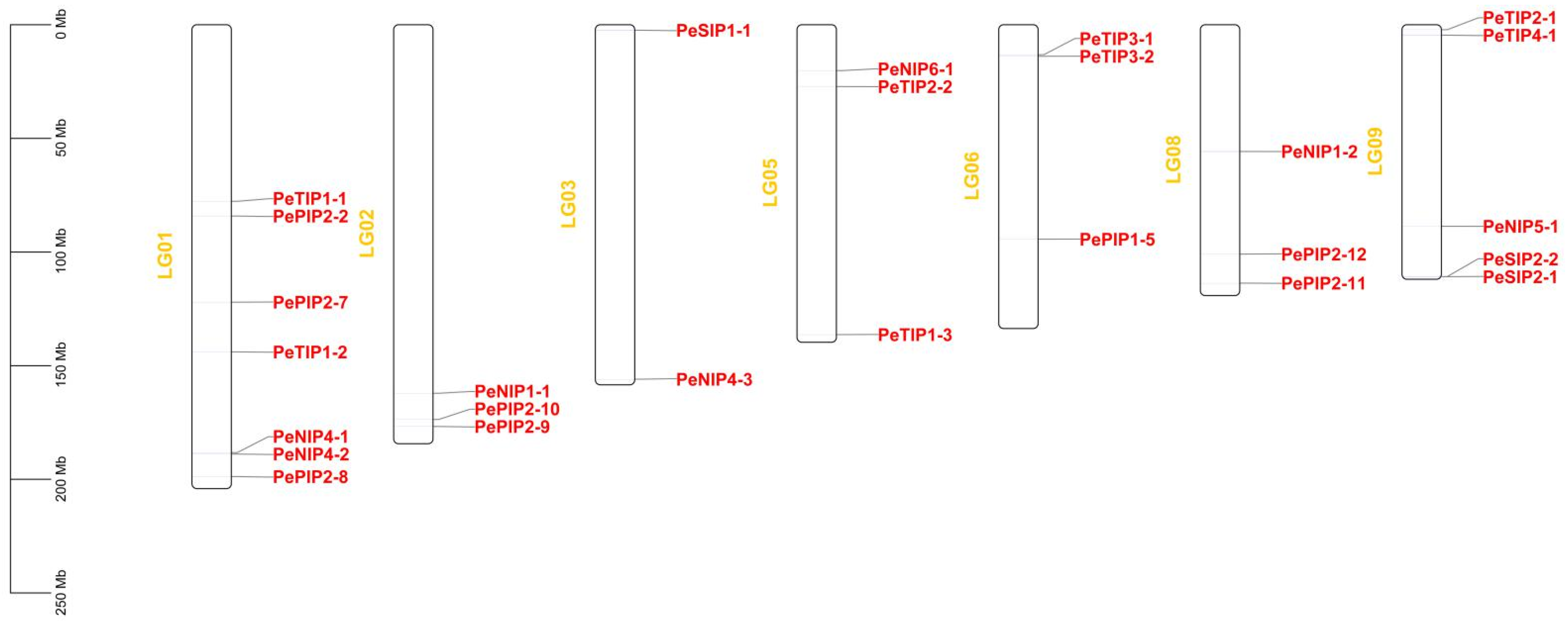
2.4. Chromosome Distribution of AQP Genes
2.5. Promoter Analysis of PeAQP Genes
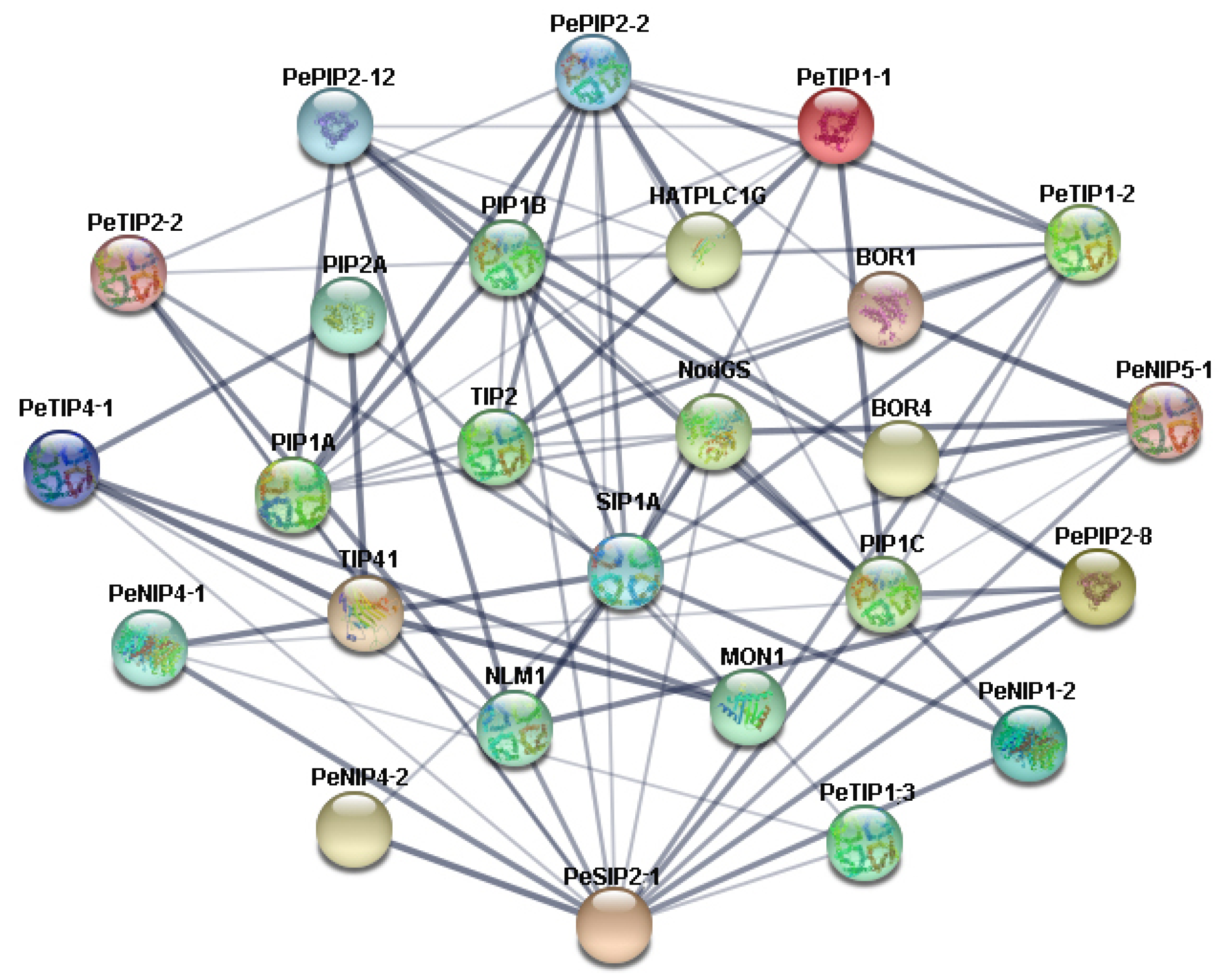
2.6. Interaction Network Analysis of PeAQPs
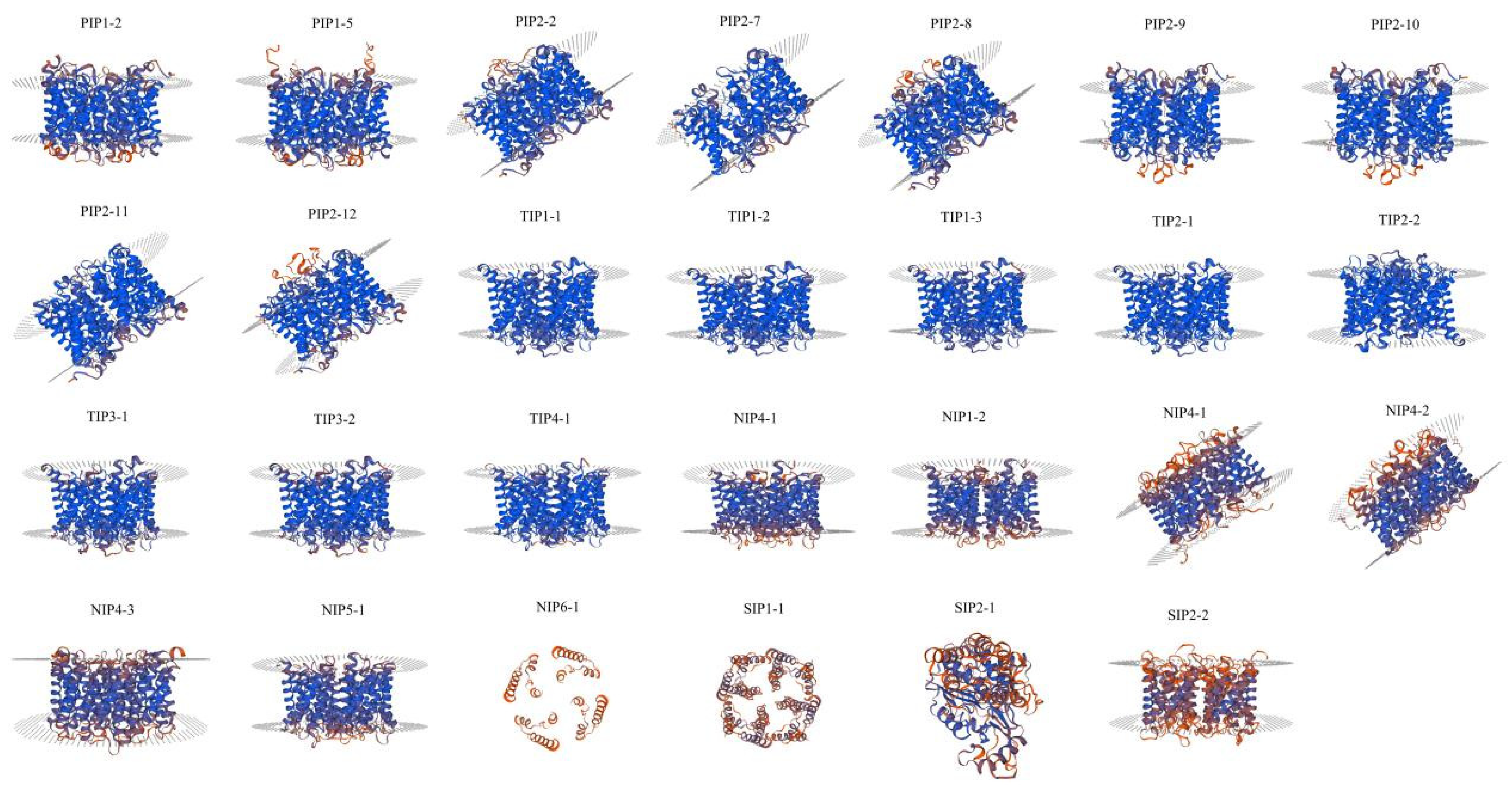
2.7. Homology Modeling
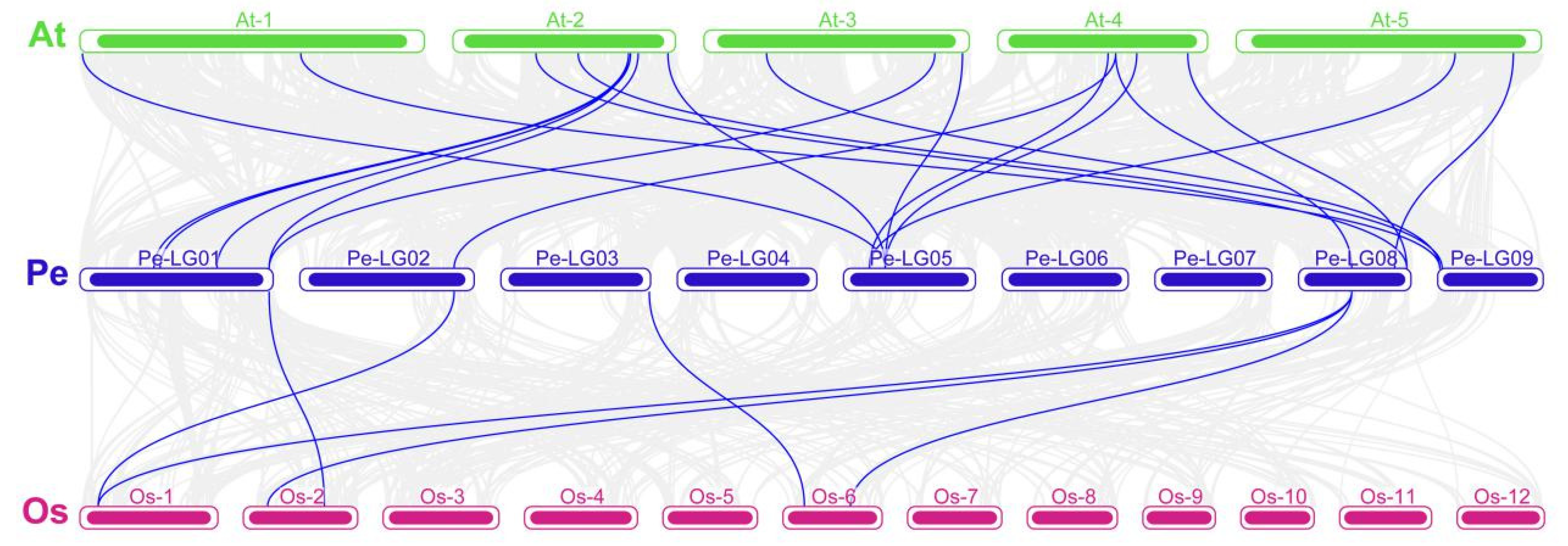
2.8. Collinearity Analysis
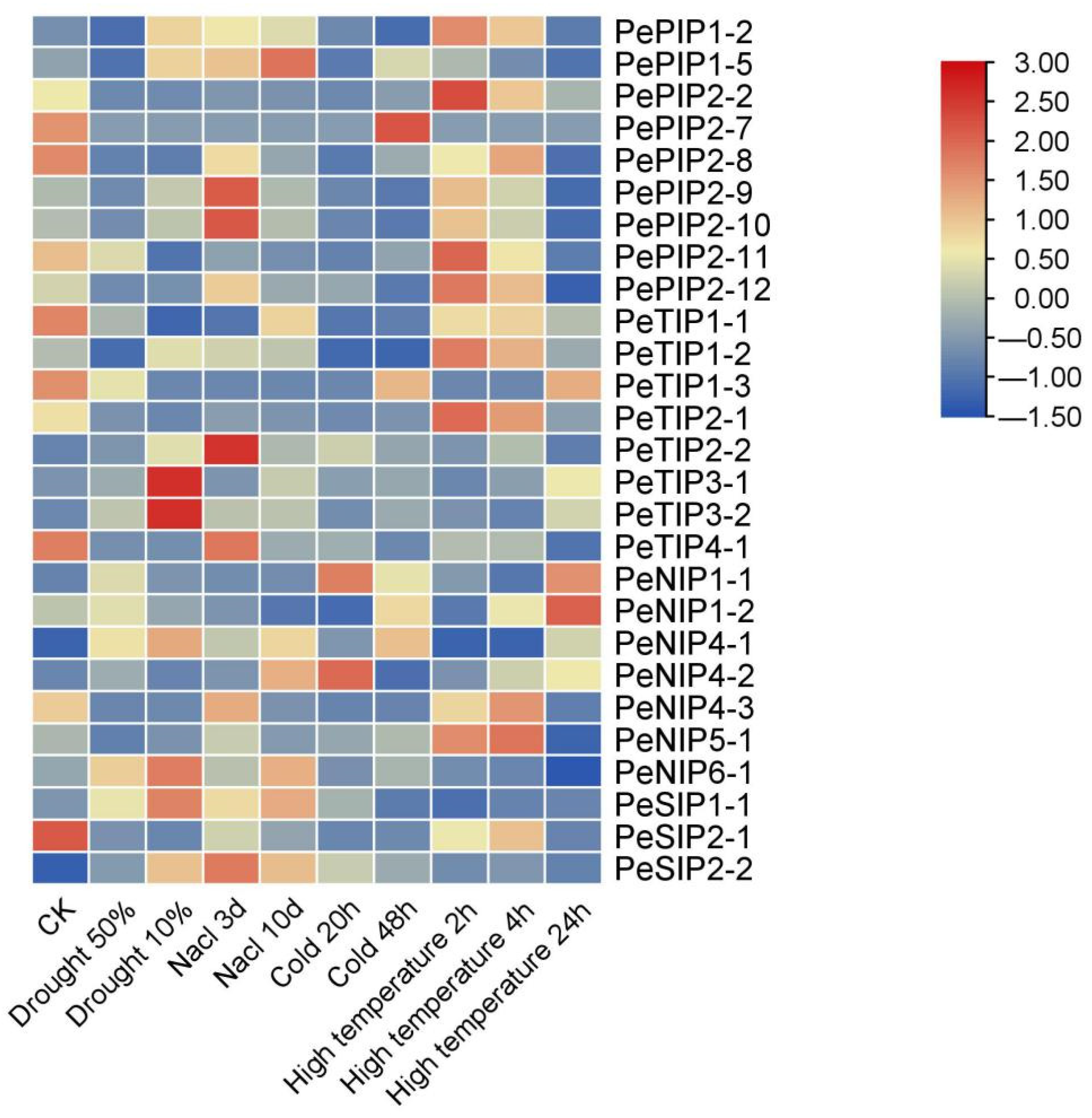
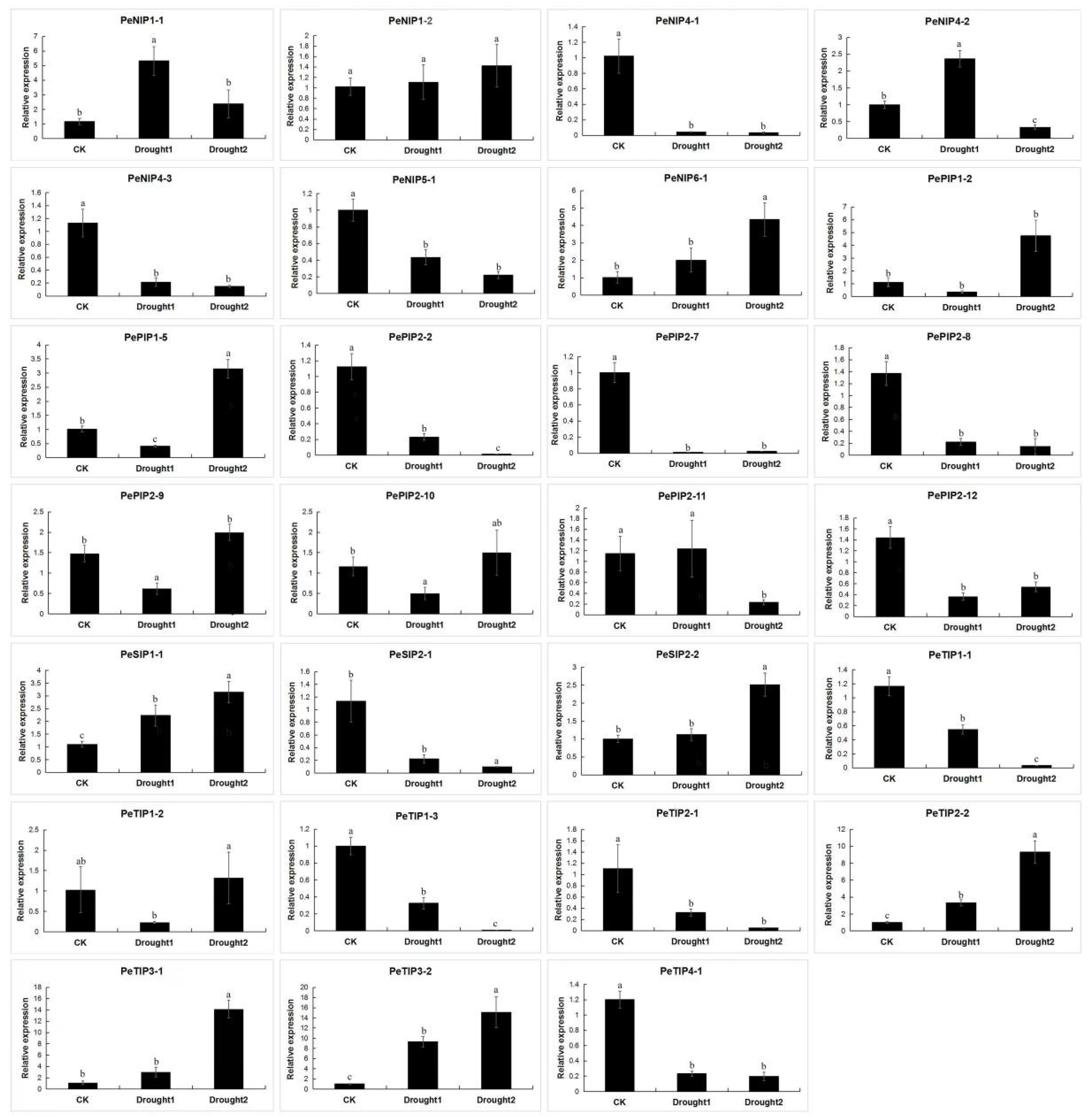
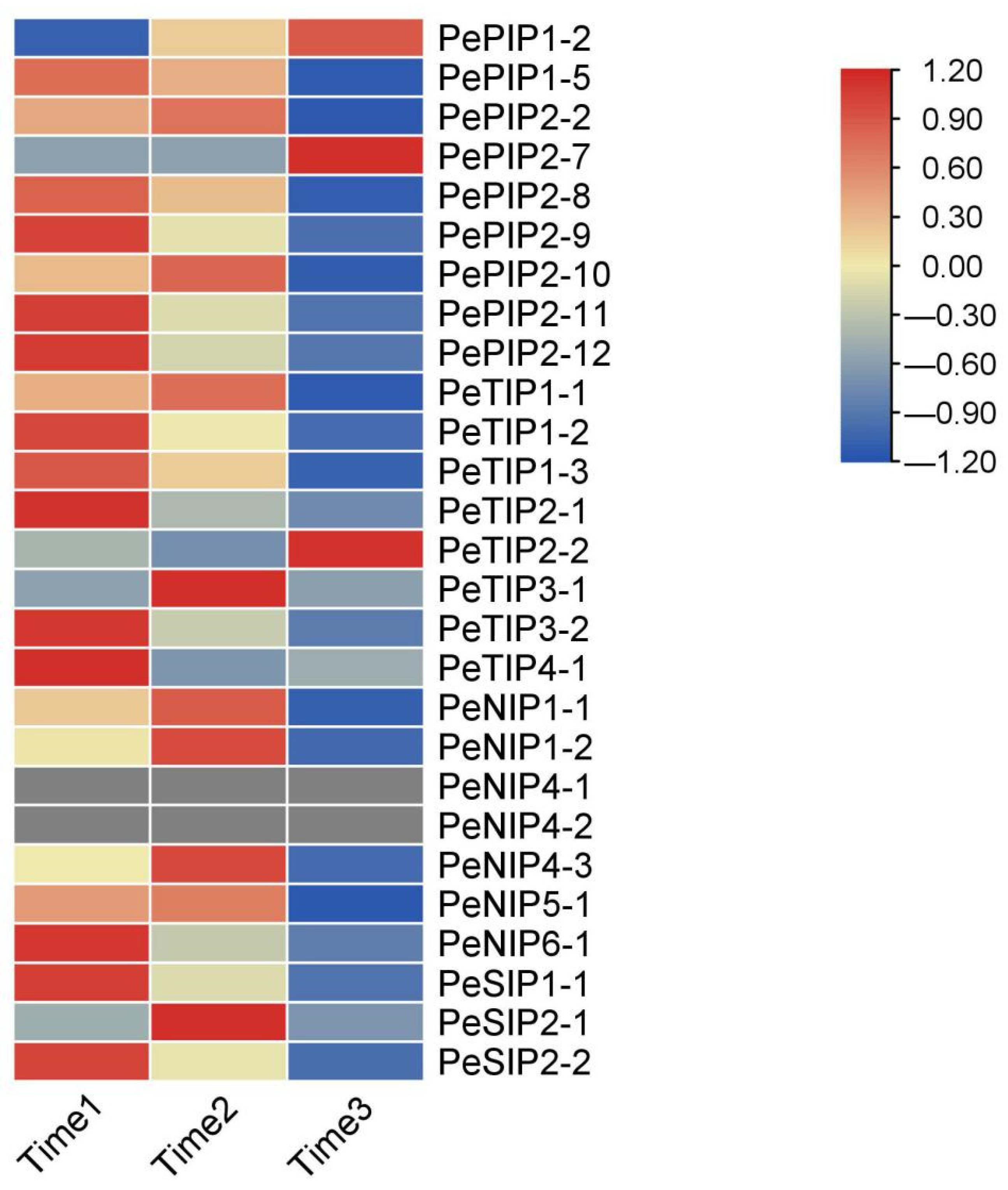
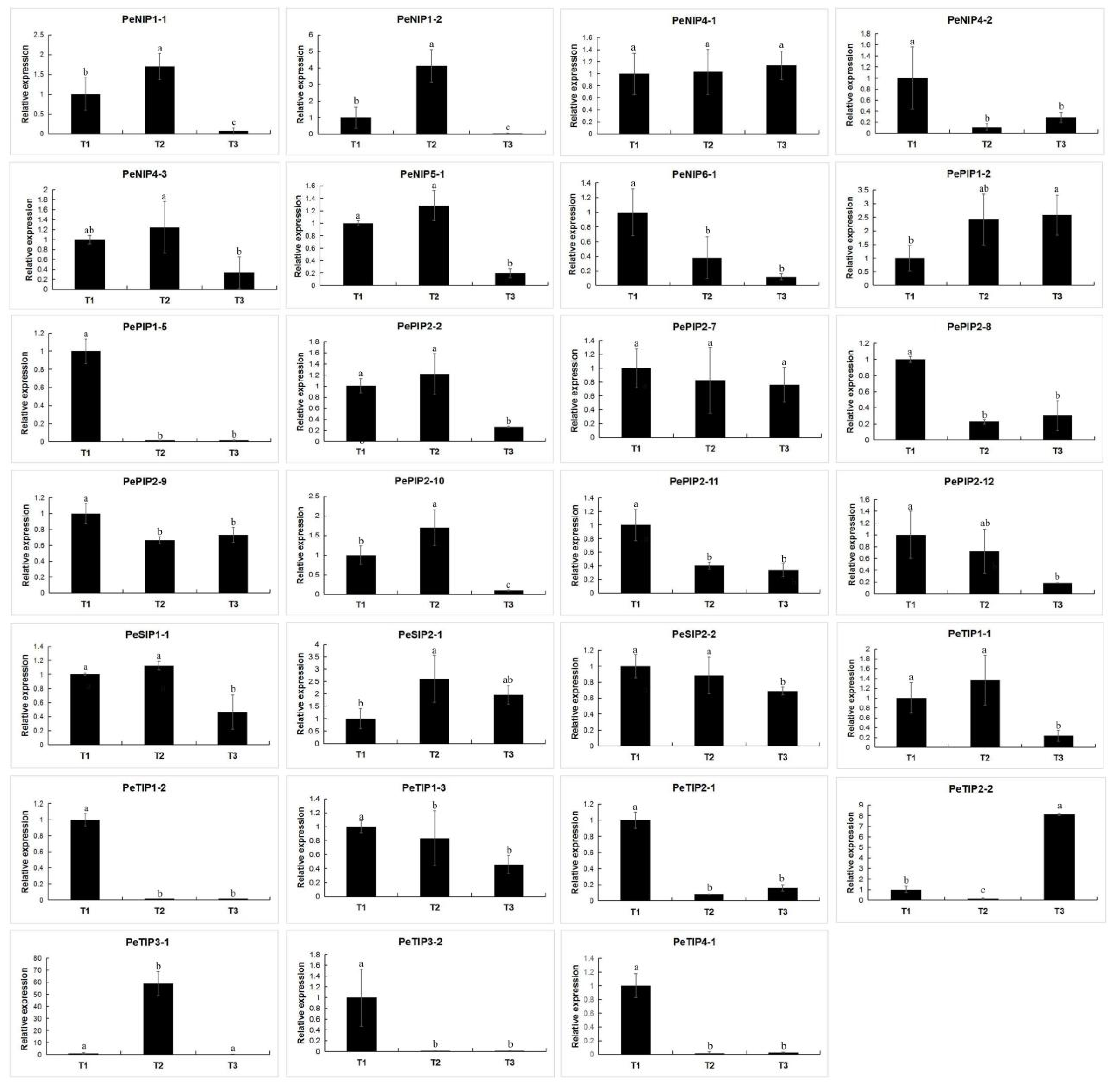
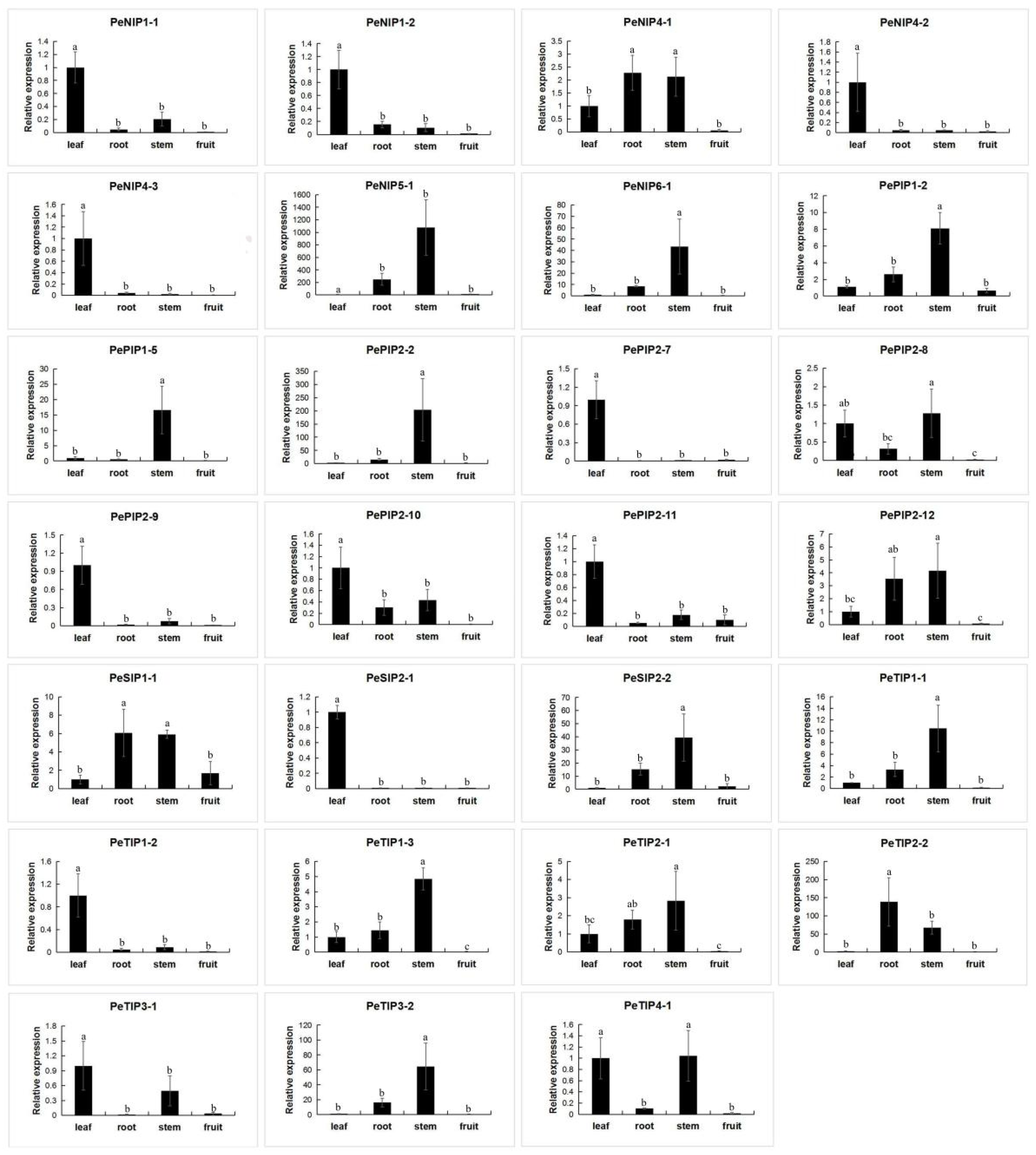
2.9. Expression Pattern of AQPs in Passion Fruit
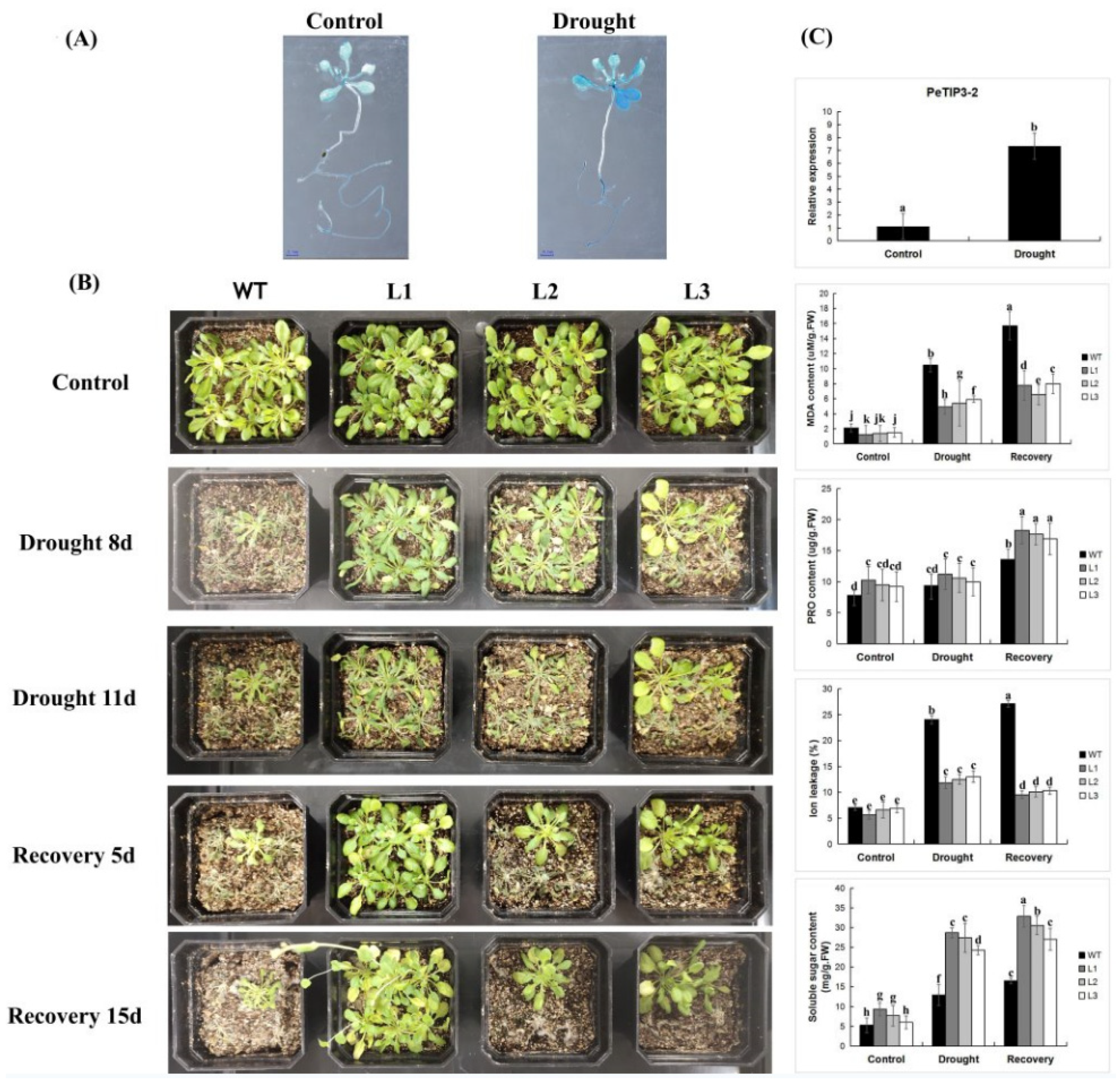
2.10. Overexpression of PeTIP3-2 Enhances Transgenic Arabidopsis Tolerance to Drought Stress
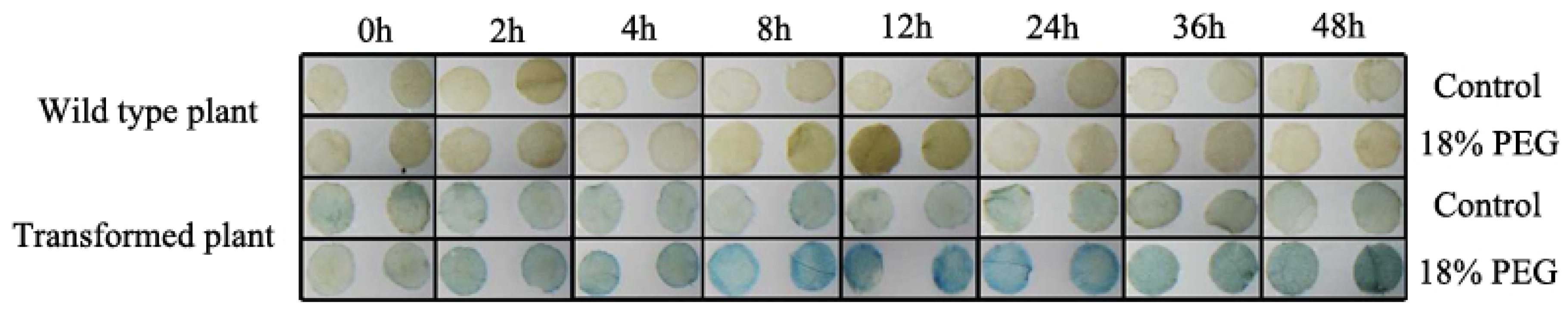
2.11. Osmotic Stress-Induced Activity Analysis in Transgenic Tobacco
3. Discussion
3.1. The Evolutionary Characteristics of the AQP Family in Plants
3.2. The Function of AQP Promoters in Plants
3.3. The Functional Diversity of AQPs in Plants
4. Materials and Methods
4.1. Identification of AQP Genes in Passion Fruit
4.2. Gene Identification, Gene Structure, and Chromosomal Locations
4.3. Analysis of cis-Acting Elements of PeAQP Genes
4.4. Homology Modeling of PeAQP 3D Structure
4.5. Analysis of Gene Protein Interaction Regulatory Network
4.6. Gene Collinearity Analysis
4.7. Plant Materials and Growth Conditions
4.8. RNA Extraction, Transcriptome Sequencing, and qRT-PCR
4.9. Heat Map
4.10. Cloning and Vector Construction of PeTIP3-2
4.11. Plant Transformation and Generation of Transgenic Plants
4.12. Drought and PEG Stress Treatment in WT and Transgenic Plants
4.13. GUS Activity Detection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, D.; Xu, Y.; Wu, B.; Ma, F.N.; Song, S. Comparative analysis of basic quality of passion fruits (Passiflora edulis sims) in Guangxi, Guizhou and Fujian, China. Bangladesh J. Bot. 2019, 48, 901–906. [Google Scholar]
- Costa, J.L.; Jesus, O.N.D.; Oliverira, G.A.F.; Oliverira, E.J.D. Effect of selection on genetic variability in yellow passion fruit. Crop Breed. Appl. Biotechnol. 2012, 12, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Kapilan, R.; Maryam Vaziri, R.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Hu, W.; Liu, J.; Song, S.; Hou, X.; Jia, C.; Li, J.; Miao, H.; Wang, Z.; Tie, W.W.; et al. An aquaporin gene MaPIP2-7 is involved in tolerance to drought, cold and salt stresses in transgenic banana (Musa acuminata L.). Plant Physiol. Biochem. 2020, 147, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Tanveer, R.; Mustafa, G.; Farooq, M.; Amin, I.; Mansoor, S. Comparative phylogenetic analysis of aquaporins provides insight into the gene family expansion and evolution in plants and their role in drought tolerant and sus-ceptible chickpea cultivars. Genomics 2020, 112, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, J.; Song, S.; Liu, J.; Hou, X.; Li, Y.; Wang, A.; Huang, D.; Wang, J.; Xu, Z.; et al. A novel aquaporin gene MaSIP2-1 confers tolerance to drought and cold stresses in transgenic banana. Mol. Breed. 2020, 40, 62. [Google Scholar] [CrossRef]
- Feng, Z.J.; Xu, S.C.; Liu, N.; Zhang, G.W.; Hu, Q.Z.; Xu, Z.S.; Gong, Y.M. Identification of the AQP members involved in abiotic stress responses from Arabidopsis. Gene 2018, 646, 64–73. [Google Scholar] [CrossRef]
- Li, D.D.; Ruan, X.M.; Zhang, J.; Wu, Y.J.; Wang, X.L.; Li, X.B. Cotton plasma membrane intrinsic protein 2s (PIP2s) selectively interact to regulate their water channel activities and are required for fibre development. New Phytol. 2013, 199, 695–707. [Google Scholar] [CrossRef]
- Alleva, K.; Marquez, M.; Villarreal, N.; Mut, P.; Bustamante, C.; Bellati, J.; Martínez, G.; Civello, M.; Amodeo, G. Cloning, functional characterization, and co-expression studies of a novel aquaporin (FaPIP2;1) of strawberry fruit. J. Exp. Bot. 2010, 61, 3935–3945. [Google Scholar] [CrossRef] [Green Version]
- Picaud, S.; Becq, F.; Dédaldéchamp, F.; Ageorges, A.; Delrot, S. Cloning and expression of two plasma membrane aquaporins expressed during the ripening of grape berry. Funct. Plant Biol. 2003, 30, 621–630. [Google Scholar] [CrossRef]
- Reuscher, S.; Akiyama1, M.; Mori1, C.; Aoki, K.; Shibata, D.; Shiratake, K. Genome-Wide Identification and Expression Analysis of Aquaporins in Tomato. PLoS ONE 2013, 8, e79052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Liu, J.; Jia, C.; Hu, W.; Song, S.; Xu, B.; Jin, Z. Overexpression of an banana aquaporin gene MaPIP1;1 enhances tolerance to multiple abiotic stresses in transgenic banana andanalysis of its interacting transcription factors. Front. Plant Sci. 2021, 12, 780544. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.L.; Yu, X.; Ye, Q.; Ding, X.D.; Kitagawa, Y.; Kwak, S.; Su, W.A.; Tang, Z.C. The role of aquaporin RWC3 in drought avoidance in rice. Plant Cell Physiol. 2004, 45, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.J.; Liu, N.; Zhang, G.W.; Niu, F.G.; Xu, S.C.; Gong, Y.M. Investigation of the AQP family in soybean and the promoter activity of TIP2;6 in heat stress and hormone responses. Int. J. Mol. Sci. 2019, 20, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, D.; Zhu, G.; Mi, X.; Guo, W. Combining genome-wide and transcriptome-wide analyses reveal the evolutionary conservation and functional diversity of aquaporins in cotton. BMC Genom. 2019, 20, 538. [Google Scholar] [CrossRef] [PubMed]
- Madrid-Espinoza, J.; Brunel-Saldias, N.; Guerra, F.P.; Gutiérrez, A.; Pozo, A.D. Genome-Wide identification and transcriptional regulation of aquaporin genes in bread wheat (Triticum aestivum L.) under water stress. Genes 2018, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Sade, N.; Gebretsadik, M.; Seligmann, R.; Schwartz, A.; Wallach, R.; Moshelion, M. The role of tobacco Aquaporin1 in improving water use efficiency, hydraulic conductivity, and yield production under salt stress. Plant Physiol. 2010, 152, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; He, X.; Zhao, B.; Zhou, C.; Liang, Y.; Ge, R.; Shen, Y.; Huang, Z. Overexpressing a putative aquaporin gene from wheat, TaNIP, enhances salt tolerance in transgenic Arabidopsis. Plant Cell Physiol. 2010, 51, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Liu, X.; Zhu, J.; Fan, W.; Zhang, Z. An aquaporin gene from halophyte Sesuvium portulacastrum, SpAQP1, increases salt tolerance in transgenic tobacco. Plant Cell Rep. 2016, 35, 385–395. [Google Scholar] [CrossRef]
- Xu, C.H.; Wang, M.; Zhou, L. Heterologous Expression of the Wheat Aquaporin Gene TaTIP2;2 Compromises the Abiotic Stress Tolerance of Arabidopsis thaliana. PLoS ONE 2013, 8, e79618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Hu, W.; Liu, J.H.; Zhang, J.B.; Jia, C.H.; Miao, H.X.; Xu, B.Y.; Jin, Z.Q. A banana aquaporin gene, MaPIP1;1, is involved in tolerance to drought and salt stresses. BMC Plant Biol. 2014, 14, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, P.; Sade, N.; Arzani, A.; Wilhelmi, M.D.M.R.; Coe, K.M.; Li, B.; Blumwald, E. Effects of abiotic stress on physiological plasticity and water use of Setaria viridis (L.). Plant Sci. 2016, 251, 128–138. [Google Scholar] [CrossRef]
- Obaid, A.Y.; Sabir, J.S.; Atef, A.; Liu, X.; Edris, S.; El-Domyati, F.M.; Mutwakil, M.Z.; Gadalla, N.O.; Hajrah, N.H.; Al-Kordy, M.A. Analysis of transcriptional response to heat stress in Rhazya stricta. BMC Plant Biol. 2016, 16, 252. [Google Scholar] [CrossRef] [Green Version]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between drought, heat and air humidity responses revealed by transcriptome-metabolome co-analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Johanson, U.; Karlsson, M.; Johansson, I.; Gustavsson, S.; Sjovall, S.; Fraysse, L.; Weig, A.R.; Kjellbom, P. The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol. 2001, 126, 1358–1369. [Google Scholar] [CrossRef] [Green Version]
- Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M.J.; Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol. 2001, 125, 1206–1215. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Hou, X.; Huang, C.; Yan, Y.; Tie, W.; Ding, Z.; Wei, Y.; Liu, J.; Miao, H.; Lu, Z.; et al. Genome-wide identification and expression analyses of aquaporin gene family during development and abiotic stress in banana. Int. J. Mol. Sci. 2015, 16, 19728–19751. [Google Scholar] [CrossRef] [Green Version]
- Tao, P.; Zhong, X.; Li, B.; Wang, W.; Yue, Z.; Lei, J.; Guo, W.; Huang, X. Genome-wide identification and characterization of aquaporin genes (AQPs) in Chinese cabbage (Brassica rapa ssp. pekinensis). Mol. Genet Genom. 2014, 289, 1131–1145. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Huang, D.; Zhang, S.; Wang, W.; Ma, F.; Wu, B.; Xu, Y.; Xu, B.; Chen, D.; Zou, M.; et al. Chromosome-scale genome assembly provides insights into the evolution and flflavor synthesis of passion fruit (Passiflflora edulis Sims). Hortic. Res. 2021, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.X.; Moon, S.; Jung, K.H. Genome–wide expression analysis of rice aquaporin genes and development of a functional gene network mediated by aquaporin expression in roots. Planta 2013, 238, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Hove, R.M.; Ziemann, M.; Bhave, M. Identification and expression analysis of the barley (Hordeum vulgare L.) aquaporin gene family. PLoS ONE 2015, 10, e0128025. [Google Scholar] [CrossRef] [PubMed]
- Hove, R.M.; Bhave, M. Plant aquaporins with non-aqua functions: Deciphering the signature sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar] [CrossRef]
- de Paula Santos Martins, C.; Pedrosa, A.M.; Du, D.; Gonçalves, L.P.; Yu, Q.; Gmitter, F.G., Jr. Genome-wide characterization and expression analysis of major intrinsic proteins during abiotic and biotic stresses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0138786. [Google Scholar]
- Maurel, C.; Verdoucq, L.; Luu, D.-T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [Green Version]
- Wudick, M.M.; Luu, D.T.; Maurel, C. A look inside. Localization patterns and functions of intracellular plant aquaporins. New Phytol. 2009, 184, 289–302. [Google Scholar] [CrossRef]
- Lopez-Zaplana, A.; Nicolas-Espinosa, J.; Carvajal, M.; Bárzana, G. Genome-wide analysis of the aquaporin genes in melon (Cucumis melo L.). Sci. Rep. 2020, 10, 22240. [Google Scholar] [CrossRef]
- Chaumont, F.; Barrieu, F.; Jung, R.; Chrispeels, M.J. Plasma membrane intrinsic proteins from maize cluster in two sequence subgroups with difffferential aquaporin activity. Plant Physiol. 2000, 122, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Uehlein, N.; Otto, B.; Hanson, D.T.; Fischer, M.; McDowell, N.; Kaldenhoff, R. Function of Nicotiana tabacum aquaporins as chloroplast gas pores challenges the concept of membrane CO2 permeability. Plant Cell 2008, 20, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattolin, S.; Sorieul, M.; Frigerio, L. Mapping of tonoplast intrinsic proteins in maturing and germinating Arabidopsis seeds reveals dual localization of embryonic TIPs to the tonoplast and plasma membrane. Mol. Plant 2011, 4, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, D.T.; Martiniére, A.; Sorieul, M.; Runions, J.; Maurel, C. Fluorescence recovery after photobleaching reveals high cycling dynamics of plasma membrane aquaporins in Arabidopsis roots under salt stress. Plant J. 2012, 69, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Deokar, A.A.; Tar’an, B. Genome-Wide Analysis of the Aquaporin Gene Family in Chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F. Identifification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhara, I.; Ugaki, M.; Hirochika, H.; Ohshima, M.; Murakami, T.; Gotoh, Y. Efficient promoter cassettes for enhanced expression of foregin genes in dicotyledonous and monocotyledonous plants. Plant Cell Physiol. 1996, 37, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Potenza, L.; Cucchiarini, L.; Piatti, E.; Angelini, U.; Dacha, M. Effects of high static magnetic field exposure on different DNAs. Bioelectromagnetics 2004, 25, 352–355. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Petrillo, E.; Godoy Herz, M.A.; Barta, A.; Kalyna, M.; Kornblihtt, A.R. Let there be light: Regulation of gene expression in plants. RNA Biol. 2014, 11, 1215–1220. [Google Scholar] [CrossRef] [Green Version]
- Baaziz, K.B.; Lopez, D.; Rabot, A.; Combes, D.; Gousset, A.; Bouzid, S. Light-mediated K(leaf) induction and contribution of both the PIP1s and PIP2s aquaporins in fifive tree species: Walnut (Juglans regia) case study. Tree Physiol. 2012, 32, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Xu, Y.; Huang, D.; Miao, H.; Liu, J.; Jia, C.; Hu, W.; Valarezo, A.V.; Xu, B.; Jin, Z. Identifification of a novel promoter from banana aquaporin family gene (MaTIP1;2) which responses to drought and salt-stress in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2018, 128, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Hu, W.; Deng, X.; Ma, Z.; Chen, L.; Huang, C.; Wang, C.; Wang, J.; He, Y.; Yang, G. Overexpression of the wheat aquaporin gene, TaAQP7, enhances drought tolerance in transgenic tobacco. PLoS ONE 2012, 7, e52439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreedharan, S.; Shekhawat, U.K.; Ganapathi, T.R. Constitutive and stress-inducible overexpression of a native aquaporin gene (MusaPIP2;6) in transgenic banana plants signals its pivotal role in salt tolerance. Plant Mol. Biol. 2015, 88, 41–52. [Google Scholar] [CrossRef]
- Choat, B.; Gambetta, G.A.; Shackel, K.A.; Matthews, M.A. Vascular function in grape berries across development and its relevance to apparent hydraulic isolation. Am. Soc. Plant Physiol. 2009, 151, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4, molecular evolutionary genetics analysis MEGA software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1199–1202. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, J.L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo, C.T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatilegene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | CDS Length (bp) | Protein Length (aa) | Molecular Formula | MW (Da) | pI | Number of Phosphate Sites | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| PePIP1-2 | P_edulia060015940.g | 864 | 287 | C1421H2155N359O380S11 | 30,700.58 | 8.22 | Ser:11 Thr:8 Tyr:5 | Cell membrane |
| PePIP1-5 | P_edulia020006715.g | 858 | 285 | C1416H2163N357O377S9 | 30,508.46 | 8.21 | Ser:9 Thr:6 Tyr:5 | Cell membrane |
| PePIP2-2 | P_edulia010001259.g | 771 | 256 | C1276H1975N329O332S5 | 27,397.02 | 7.11 | Ser:4 Thr:3 Tyr:3 | Cell membrane |
| PePIP2-7 | P_edulia020006513.g | 597 | 198 | C979H1497N257O255S6 | 21,139.58 | 9.71 | Ser:10 Thr:4 Tyr:3 | Cell membrane |
| PePIP2-8 | P_edulia010005210.g | 741 | 246 | C1203H1854N292O319S7 | 25,736.14 | 5.57 | Ser:7 Thr:5 Tyr:2 | Cell membrane |
| PePIP2-9 | P_edulia020006819.g | 858 | 285 | C1416H2163N357O377S9 | 30,508.46 | 8.21 | Ser:9 Thr:6 Tyr:5 | Cell membrane |
| PePIP2-10 | P_edulia080019927.g | 791 | 252 | C1201H1826N294O325S5 | 25,743.78 | 5.55 | Ser:5 Thr:5 Tyr:1 | Cell membrane |
| PePIP2-11 | P_edulia080019415.g | 744 | 247 | C1177H1803N285O320S5 | 25,226.28 | 5.86 | Ser:9 Thr:4 Tyr:4 | Cell membrane |
| PePIP2-12 | P_edulia010001027.g | 753 | 250 | C1175H1800N278O322S3 | 25,069.07 | 4.93 | Ser:12 Thr:1 Tyr:3 | Cell membrane |
| PeTIP1-1 | P_edulia010000969.g | 758 | 252 | C1210H1834N296O326S5 | 25,903.96 | 5.71 | Ser:8 Thr:5 Tyr:1 | Cell membrane |
| PeTIP1-2 | P_edulia010001331.g | 477 | 158 | C826H1244N196O201S6 | 17,328.52 | 9.3 | Ser:2 Thr:4 Tyr:2 | Cell membrane |
| PeTIP1-3 | P_edulia050012817.g | 1794 | 597 | C2924H4612N802O845S29 | 65,451.39 | 7.3 | Ser:29 Thr:3 Tyr:0 | Cell membrane |
| PeTIP2-1 | P_edulia090020609.g | 711 | 236 | C1195H1882N304O305S9 | 25,676.48 | 10.09 | Ser:11 Thr:5 Tyr:0 | Cell membrane |
| PeTIP2-2 | P_edulia050011915.g | 900 | 299 | C1427H2243N371O399S10 | 31,301.35 | 7.69 | Ser:13 Thr:8 Tyr:3 | Cell membrane |
| PeTIP3-1 | P_edulia060014309.g | 858 | 285 | C1384H2188N348O386S12 | 30,263.42 | 6.43 | Ser:13 Thr:6 Tyr:6 | Cell membrane |
| PeTIP3-2 | P_edulia060014363.g | 831 | 276 | C1333H2091N339O381S12 | 29,347.03 | 8.24 | Ser:15 Thr:8 Tyr:3 | Cell membrane |
| PeTIP4-1 | P_edulia090020949.g | 756 | 251 | C1195H1815N293O329S9 | 25,838.86 | 4.97 | Ser:9 Thr:4 Tyr:1 | Cell membrane |
| PeNIP1-1 | P_edulia080019109.g | 861 | 286 | C1417H2156N360O373S8 | 30,459.37 | 8.84 | Ser:13 Thr:7 Tyr:2 | Cell membrane |
| PeNIP1-2 | P_edulia010004282.g | 858 | 278 | C1370H2103N347O365S10 | 29,595.47 | 8.61 | Ser:11 Thr:5 Tyr:4 | Cell membrane |
| PeNIP4-1 | P_edulia010004322.g | 837 | 285 | C1408H2164N358O379S9 | 30,459.38 | 8.22 | Ser:10 Thr:8 Tyr:4 | Cell membrane |
| PeNIP4-2 | P_edulia030009116.g | 909 | 302 | C1444H2270N370O410S14 | 31,822.98 | 7.77 | Ser:22 Thr:9 Tyr:2 | Cell membrane |
| PeNIP4-3 | P_edulia090022062.g | 234 | 77 | C351H569N103O107S4 | 8072.24 | 9.69 | Ser:3 Thr:7 Tyr:0 | Nucleus |
| PeNIP5-1 | P_edulia050011768.g | 861 | 286 | C1421H2186N360O384S9 | 30,745.71 | 7.67 | Ser:6 Thr:10 Tyr:4 | Cell membrane |
| PeNIP6-1 | P_eduliaContig210022875.g | 864 | 273 | C1365H2091N341O361S9 | 29,343.23 | 8.99 | Ser:8 Thr:10 Tyr:4 | Cell membrane |
| PeSIP1-1 | P_edulia030007640.g | 858 | 285 | C1383H2188N346O386S12 | 30,223.39 | 6.36 | Ser:13 Thr:6 Tyr:6 | Cell membrane |
| PeSIP2-1 | P_edulia090022232.g | 773 | 257 | C1259H1968N312O351S11 | 27,444.02 | 6.82 | Ser:17 Thr:6 Tyr:2 | Cell membrane |
| PeSIP2-2 | P_edulia090022235.g | 771 | 256 | C1275H1973N329O332S5 | 27,383 | 7.11 | Ser:4 Thr:3 Tyr:2 | Cell membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Zhang, D.; Ma, F.; Xing, W.; Huang, D.; Wu, B.; Chen, J.; Chen, D.; Xu, B.; Xu, Y. Genome-Wide Identification and Expression Analyses of the Aquaporin Gene Family in Passion Fruit (Passiflora edulis), Revealing PeTIP3-2 to Be Involved in Drought Stress. Int. J. Mol. Sci. 2022, 23, 5720. https://doi.org/10.3390/ijms23105720
Song S, Zhang D, Ma F, Xing W, Huang D, Wu B, Chen J, Chen D, Xu B, Xu Y. Genome-Wide Identification and Expression Analyses of the Aquaporin Gene Family in Passion Fruit (Passiflora edulis), Revealing PeTIP3-2 to Be Involved in Drought Stress. International Journal of Molecular Sciences. 2022; 23(10):5720. https://doi.org/10.3390/ijms23105720
Chicago/Turabian StyleSong, Shun, Dahui Zhang, Funing Ma, Wenting Xing, Dongmei Huang, Bin Wu, Jian Chen, Di Chen, Binqiang Xu, and Yi Xu. 2022. "Genome-Wide Identification and Expression Analyses of the Aquaporin Gene Family in Passion Fruit (Passiflora edulis), Revealing PeTIP3-2 to Be Involved in Drought Stress" International Journal of Molecular Sciences 23, no. 10: 5720. https://doi.org/10.3390/ijms23105720