
Modulations in Chlorophyll a Fluorescence Based on Intensity and Spectral Variations of Light


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. General Properties of Light
3. Light and Photosynthesis
3.1. Light as the Source of Energy in Photosynthesis
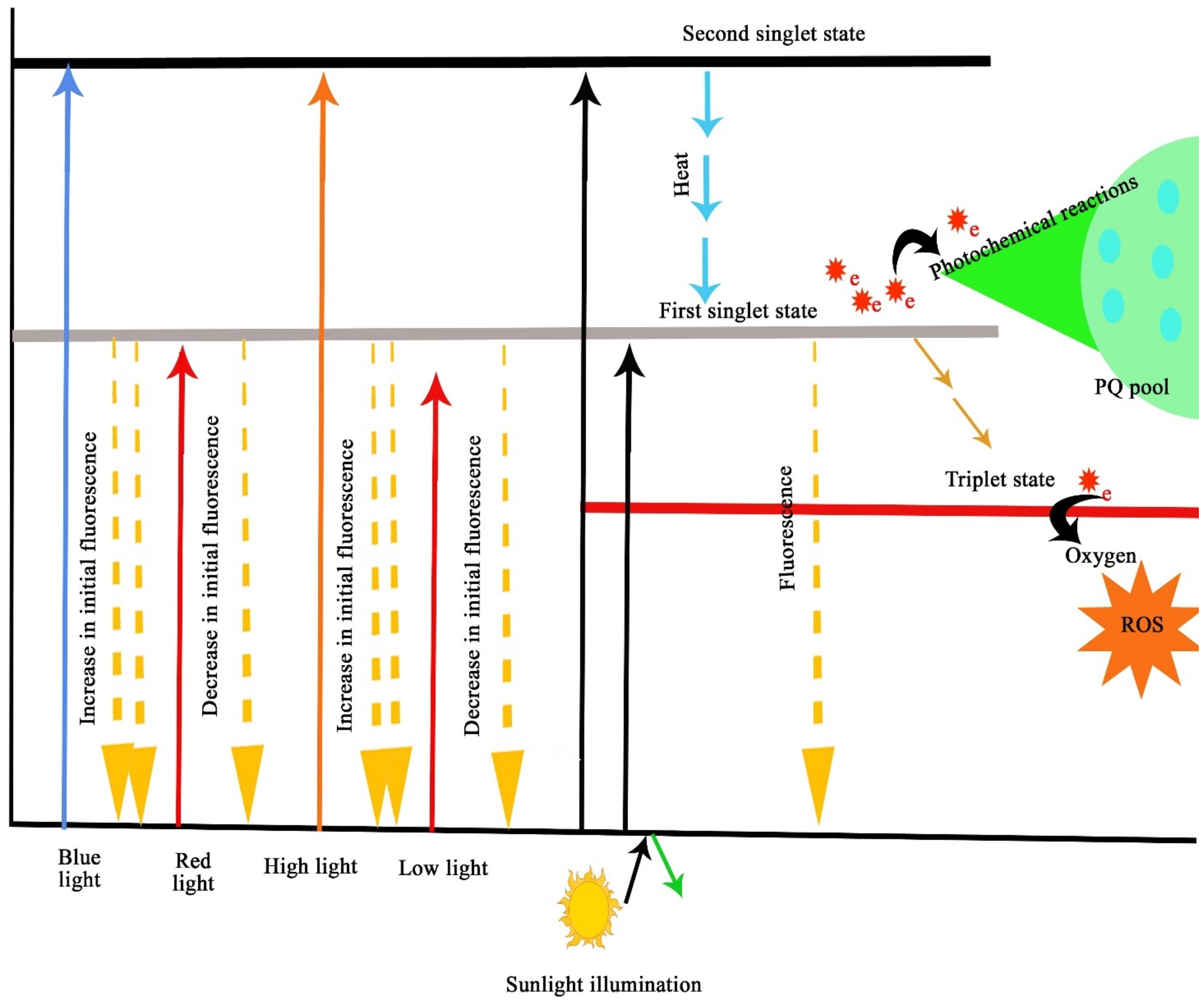
3.2. The Mechanisms of Light Interactions in Photosynthetic Efficiency of Plants
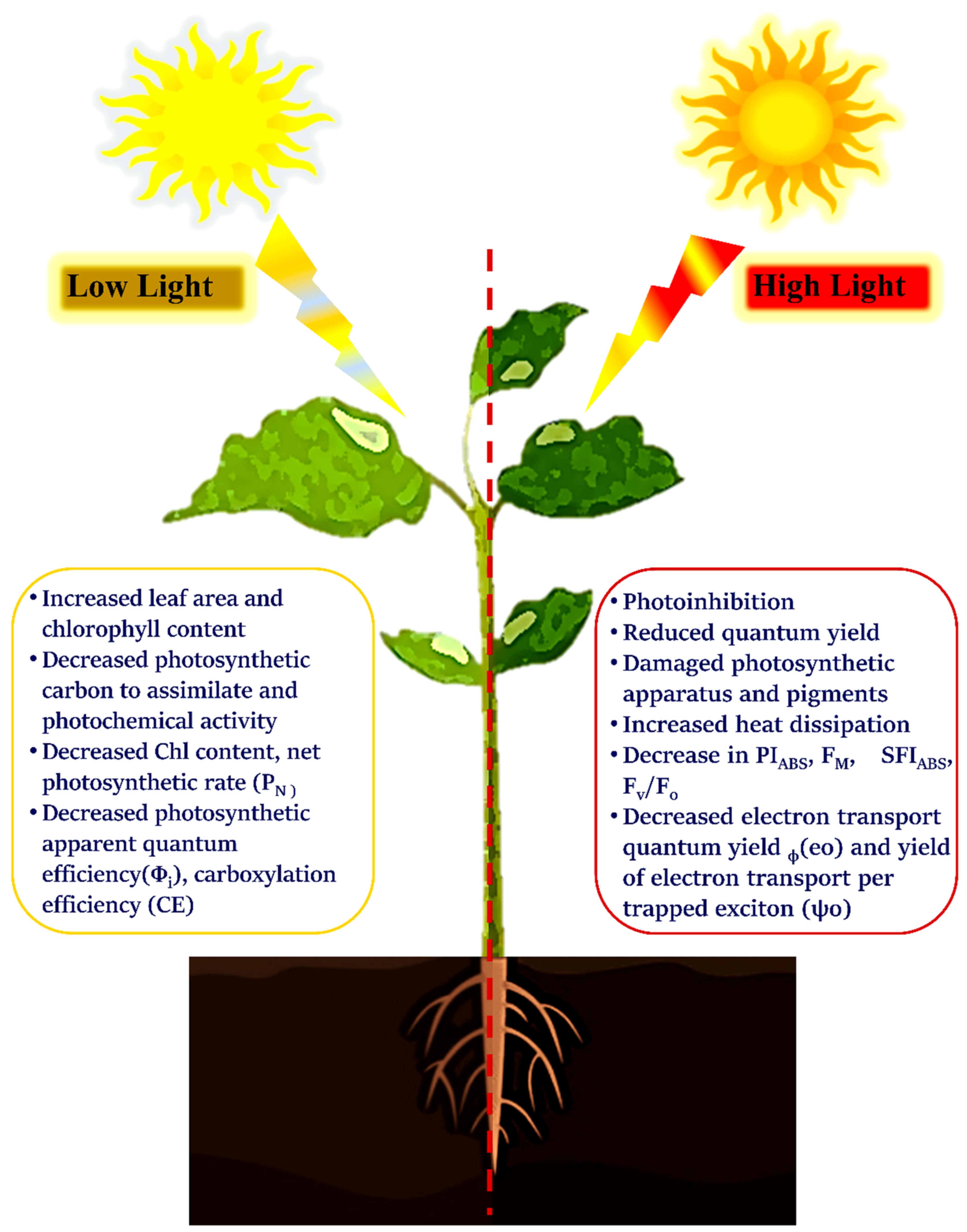
4. Impact of Light Intensity on Photosynthesis
4.1. High Light
4.2. Low Light
5. Quality of Light
5.1. UV Radiation
5.2. Ionising Radiations
5.3. White Light Exposure
5.4. Red and Far-red Light Exposure
5.5. Blue and Green Light
5.6. Combinations of Different Quality of Light and its Imprints on Photosynthetic Performance
6. Photosynthetic Responses Related to Duration of Light
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
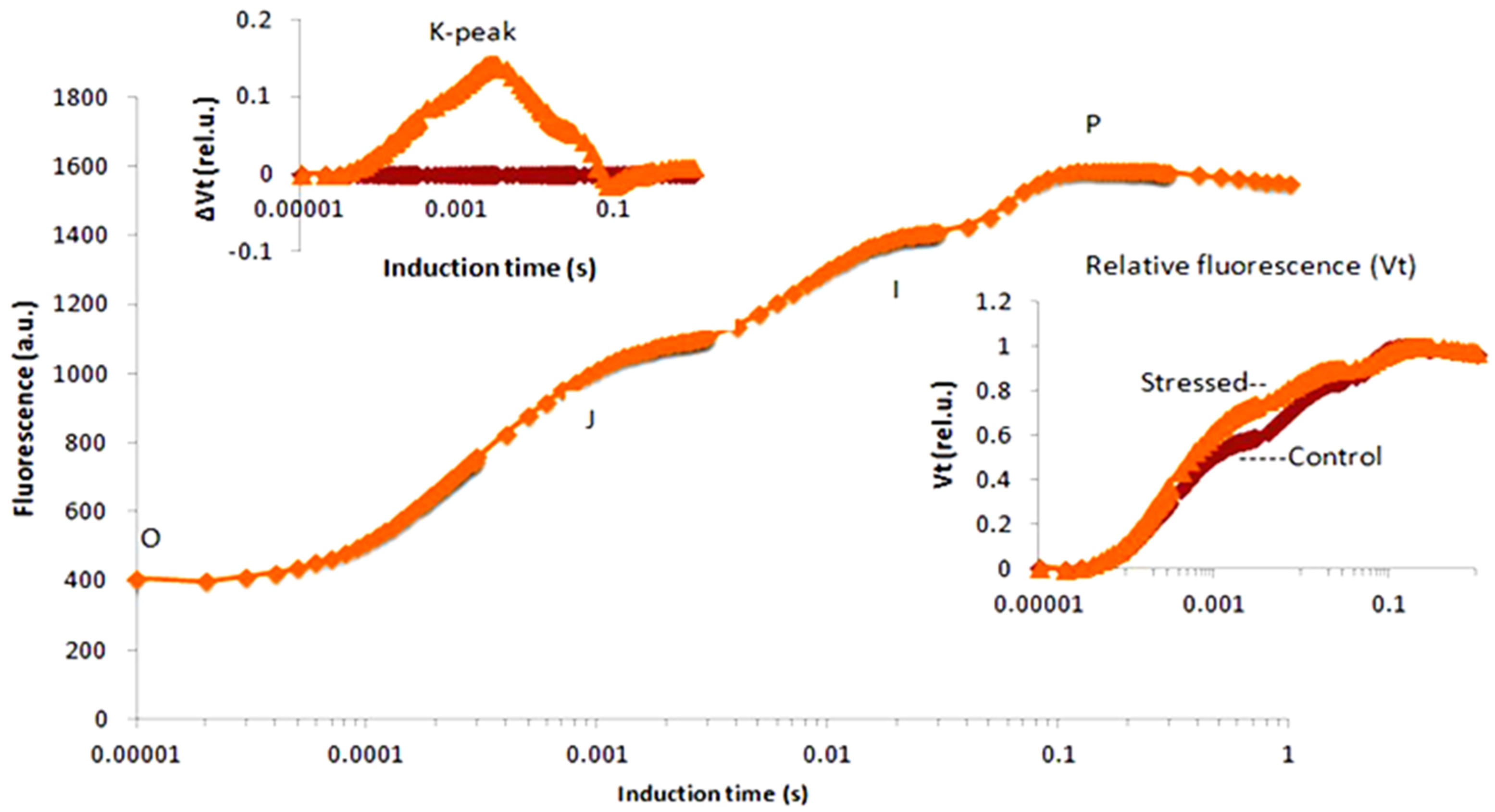
| Phases in Induction Curve | |
| O = FO | Minimal fluorescence/First step of chl a fluorescence transient |
| J = FJ | Intermediate step in the chl a fluorescence transient at 2 ms |
| I = FI | Intermediate step in the chl a fluorescence transient at 30 ms |
| P = FP = FM | Maximal fluorescence level/Final step of chl a fluorescence transient |
| K | Intermediate step in the chl a fluorescence transient at 0.3 ms |
| OJ-phase | It represents the reduction in the acceptor side of PS II |
| JI-phase | It represents the reduction in the PQ (Plastoquinone) |
| IP-phase | It represents the reduction in the acceptor side of PS I |
| Area | Area above the fluorescence induction curve |
| tFM | Time taken to reach FM |
| Other JIP Parameters | |
| FV = FM − F0 | Maximal variable fluorescence (FM-FO) |
| FV/FM | It represents the maximum quantum yield of PSII |
| FV/FO | It represents the maximum efficiency of water-splitting complex |
| SM = Area/(Fm − Fo) | It represents the multiple turnovers of QA reductions |
| SM/tFM | It represents the average redox state of QA in the time span from 0 to tFM |
| N = Sm/ΔV/Δt0 *(1/VJ) | Turnover number of QA indicates the number of times QA was reduced from 0 to tFM |
| SFI ABS | An indicator of PS II structure and functioning |
| VJ | Relative variable fluorescence at phase J of the fluorescence induction curve |
| VI | Relative variable fluorescence at phase I of the fluorescence induction curve |
| PI ABS = γRC/(1 − γRC) * φPo/(1 − φPo) × ψo/(1 − ψo) | Performance index of PS II on absorption basis |
| PI TOTAL = PIABS × δRo/(1 − δRo) | Performance index of electron flux to the final PS I electron acceptors |
| 10RC/abs | Absorption per RC |
| RC/CSM | Density of active PS II reaction centres per cross-section |
| DFABS = log (PIABS) | PSII-relative driving force index on an absorption basis |
| Kn | Non-photochemical de-excitation rate constant |
| Kp | Photochemical de-excitation rate constant |
| Yield Parameters | |
| φPo | Maximum quantum yield of primary PSII photochemistry (at t = 0) |
| φ (Do) | Quantum yield of energy dissipation |
| φ(Eo) | Quantum yield (at t = 0) for electron transport from QA− to plastoquinone |
| ψo | Probability (at t = 0) that a trapped exciton moves an electron into the electron transport chain beyond QA |
| ϒRC | Probability that PSII chlorophyll molecule functions as RC |
| δ(Ro) = (1 − VJ)/(1 − VI) | Efficiency/probability (at t = 0) with which an electron from the intersystem carriers moves to reduce end electron acceptors at the PSI acceptor side |
| Specific Energy Flux | |
| ABS/RC = (1 − γRC)/γRC | Absorption flux per RC corresponding directly to its apparent antenna size |
| TRo/RC = ΔV/Δt0 × (1/Vj) | Trapping flux leading to QA reduction per RC at t = 0 |
| ETo/RC = ΔV/Δt0 × (1/Vj) ψ0 | Electron transport flux from QA− to plastoquinone per RC at t = 0 |
| DIo/RC = (ABS/RC − TR0/RC) | Dissipated energy flux per RC at the initial moment of the measurement, i.e., at t = 0 |
| Phenomenological Energy Flux | |
| ABS/CSm | Absorption of energy per excited cross-section (CS) approximated by FM |
| TRo/CSm | Excitation energy flux trapped by PSII of a photosynthesizing sample cross-section (CS) approximated by FM |
| ETo/CSm | Electron flux transported by PSII of a photosynthesizing sample cross-section (CS) approximated by FM |
| DIo/CSm | Heat dissipation of excitation energy by PSII of a photosynthesizing sample cross-section (CS) approximated by FM |
References
- Yousef, A.F.; Ali, M.M.; Rizwan, H.M.; Tadda, S.A.; Kalaji, H.M.; Yang, H.; Ahmed, M.A.A.; Wróbel, J.; Xu, Y.; Chen, F. Photosynthetic apparatus performance of tomato seedlings grown under various combinations of LED illumination. PLoS ONE 2021, 16, e0249373. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Bin Khalid, M.H.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The Influence of Light Intensity and Leaf Movement on Photosynthesis Characteristics and Carbon Balance of Soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, K.A.; Larsson, P.; Gauslaa, Y. Light screening in lichen cortices can be quantified by chlorophyll fluorescence techniques for both reflecting and absorbing pigments. Planta 2010, 231, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Trouwborst, G.; Hogewoning, S.W.; Van Kooten, O.; Harbinson, J.; Van Ieperen, W. Plasticity of photosynthesis after the ‘red light syndrome’ in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Faseela, P.; Puthur, J.T. The imprints of the high light and UV-B stresses in Oryza sativa L. ‘Kanchana’ seedlings are differentially modulated. J. Photochem. Photobiol. B Biol. 2018, 178, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.T.D.; Puthur, J.T. UV-B priming enhances specific secondary metabolites in Oryza sativa (L.) empowering to encounter diverse abiotic stresses. Plant Growth Regul. 2020, 92, 169–180. [Google Scholar]
- Xu, Y.; Yang, M.; Cheng, F.; Liu, S.; Liang, Y. Effects of LED photoperiods and light qualities on in vitro growth and chlorophyll fluorescence of Cunninghamia lanceolata. BMC Plant Biol. 2020, 20, 269. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef] [Green Version]
- Ziegelberger, G. ICNIRP guidelines on limits of exposure to incoherent visible and infrared radiation. Health Phys. 2013, 105, 74–96. [Google Scholar]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary me-tabolism: A review. Hort Sci. 2015, 50, 1128–1135. [Google Scholar]
- Ducat, D.C.; Silver, P.A. Improving carbon fixation pathways. Curr. Opin. Chem. Biol. 2012, 16, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Narzi, D.; Capone, M.; Bovi, D.; Guidoni, L. Evolution from S3 to S4 States of the Oxygen-Evolving Complex in Photosystem II Monitored by Quantum Mechanics/Molecular Mechanics (QM/MM) Dynamics. Chem. A Eur. J. 2018, 24, 10820–10828. [Google Scholar] [CrossRef]
- Sirohiwal, A.; Neese, F.; Pantazis, D.A. Protein matrix control of reaction center excitation in photosystem II. J. Am. Chem. Soc. 2020, 142, 18174–18190. [Google Scholar] [CrossRef]
- Huner, N.; Öquist, G.; Melis, A. Photostasis in plants, green algae and cyanobacteria: The role of light harvesting antenna complexes. In Light-Harvesting Antennas in Photosynthesis; Springer: Dordrecht, The Netherlands, 2003; pp. 401–421. [Google Scholar]
- Balevičius, V.; Fox, K.F.; Bricker, W.P.; Jurinovich, S.; Prandi, I.G.; Mennucci, B.; Duffy, C.D. Fine control of chlo-rophyll-carotenoid interactions defines the functionality of light-harvesting proteins in plants. Sci. Rep. 2017, 7, 13956. [Google Scholar] [CrossRef] [Green Version]
- Emerson, R.; Rabinowitch, E. Red drop and role of auxiliary pigments in photosynthesis. Plant Physiol. 1960, 35, 477. [Google Scholar] [CrossRef] [Green Version]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of Phosphorus Deficiency on the Photosynthetic Electron Transport Chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Neupane, P.; Bhuju, S.; Thapa, N.; Bhattarai, H.K. ATP Synthase: Structure, Function and Inhibition. Biomol. Concepts 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Storti, M.; Alboresi, A.; Gerotto, C.; Aro, E.M.; Finazzi, G.; Morosinotto, T. Role of cyclic and pseudo-cyclic electron transport in response to dynamic light changes in Physcomitrella patens. Plant Cell Environ. 2019, 42, 1590–1602. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Santabarbara, S.; Agostini, G.; Casazza, A.P.; Syme, C.D.; Heathcote, P.; Böhles, F.; Evans, M.C.W.; Jennings, R.C.; Carbonera, D. Chlorophyll triplet states associated with Photosystem I and Photosystem II in thylakoids of the green alga Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 88–105. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2017, 132, 13–66. [Google Scholar] [CrossRef] [Green Version]
- Oliwa, J.; Skoczowski, A. Different response of photosynthetic apparatus to high-light stress in sporotrophophyll and nest leaves of Platyceriumbifurcatum. Photosynthetica 2019, 57, 147–159. [Google Scholar] [CrossRef]
- Lu, T.; Yu, H.; Li, Q.; Chai, L.; Jiang, W. Improving Plant Growth and Alleviating Photosynthetic Inhibition and Oxidative Stress From Low-Light Stress With Exogenous GR24 in Tomato (Solanum lycopersicum L.) Seedlings. Front. Plant Sci. 2019, 10, 490. [Google Scholar] [CrossRef]
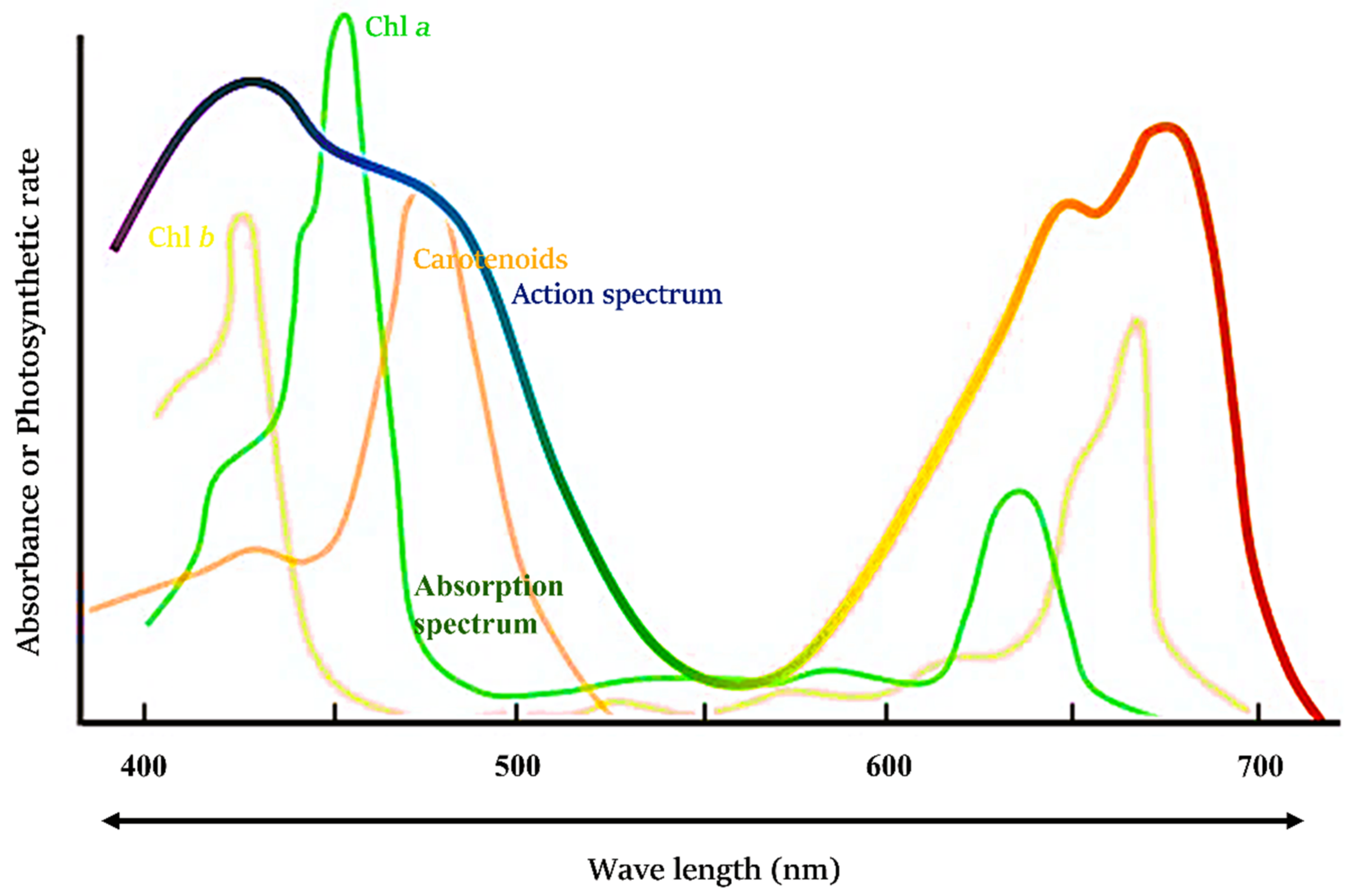
- McCree, K. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1972, 9, 191–216. [Google Scholar] [CrossRef]
- Zhen, S.; Haidekker, M.; Van Iersel, M.W. Far-red light enhances photochemical efficiency in a wavelength-dependent manner. Physiol. Plant. 2019, 167, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nishio, J.N.; Vogelmann, T.C. Green Light Drives CO2 Fixation Deep within Leaves. Plant Cell Physiol. 1998, 39, 1020–1026. [Google Scholar] [CrossRef]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 23, 539–548. [Google Scholar] [CrossRef]
- Murchie, E.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 328. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2021, 41, 742–780. [Google Scholar] [CrossRef]
- Thomas, B. Encyclopedia of Applied Plant Sciences; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Torres, R.; Romero, J.M.; Lagorio, M.G. Effects of sub-optimal illumination in plants. Comprehensive chlorophyll fluorescence analysis. J. Photochem. Photobiol. B Biol. 2021, 218, 112182. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 1984, 35, 15–44. [Google Scholar] [CrossRef]
- Osmond, C.B.; Grace, S.C. Perspectives on photoinhibition and photorespiration in the field: Quintessential ineffi-ciencies of the light and dark reactions of photosynthesis? J. Exp. Bot. 1995, 46, 1351–1362. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibi-tion. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- Messant, M.; Krieger-Liszkay, A.; Shimakawa, G. Dynamic Changes in Protein-Membrane Association for Regulating Photosynthetic Electron Transport. Cells 2021, 10, 1216. [Google Scholar] [CrossRef]
- Guo, Y.-P.; Zhou, H.-F.; Zhang, L.-C. Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci. Hortic. 2006, 108, 260–267. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 Response of Stomata and Its Dependence on Environmental Factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef] [Green Version]
- Zivcak, M.; Brestic, M.; Kunderlikova, K.; Sytar, O.; Allakhverdiev, S.I. Repetitive light pulse-induced photoinhibition of photosystem I severely affects CO2 assimilation and photoprotection in wheat leaves. Photosynth. Res. 2015, 126, 449–463. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava ATsimilli-Michael, M. Screening the vitality and photosynthetic activity of plants by fluorescent transient. In Crop Improvement for Food Security; SSARM: Hisar, India, 1999; pp. 72–115. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Faseela, P.; Puthur, J.T. Chlorophyll a fluorescence changes in response to short and long term high light stress in rice seedlings. Indian J. Plant Physiol. 2016, 22, 30–33. [Google Scholar] [CrossRef]
- Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A.; Nicola, S. Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiol. Biochem. 2016, 106, 141–148. [Google Scholar] [CrossRef]
- Zhu, S.-Q.; Zhao, H.; Liang, J.-S.; Ji, B.-H.; Jiao, D.-M. Relationships between Phosphatidylglycerol Molecular Species of Thylakoid Membrane Lipids and Sensitivities to Chilling-induced Photoinhibition in Rice. J. Integr. Plant Biol. 2008, 50, 194–202. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence parameters as early indicators of light stress in barley. J. Photochem. Photobiol. B Biol. 2012, 112, 1–6. [Google Scholar] [CrossRef]
- Lei, L.; Xiang-Yi, L.; Xin-Wen, X.; Li-Sha, L.; Fan-Jiang, Z.; Feng-Li, C. Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possesses a different adaptation mechanism to shade. Plant Physiol. Biochem. 2014, 74, 239–245. [Google Scholar] [CrossRef]
- Yangen, F.; Xiuxiu, Z.; Hanyue, W.; Yueyue, T.; Qinzeng, X.; Lixia, Z. Effects of light intensity on metabolism of light-harvesting pigment and photosynthetic system in Camellia sinensis L. cultivar ‘Huangjinya’. Environ. Exp. Bot. 2019, 166, 103796. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M. PSII Fluorescence Techniques for Measurement of Drought and High Temperature Stress Signal in Crop Plants: Protocols and Applications. In Molecular Stress Physiology of Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 87–131. ISBN 9788132208075. [Google Scholar]
- Kono, M.; Kawaguchi, H.; Mizusawa, N.; Yamori, W.; Suzuki, Y.; Terashima, I. Far-red light accelerates photosyn-thesis in the low-light phases of fluctuating light. Plant Cell Physiol. 2020, 61, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhong, H.; Wang, J.; Sui, X.; Xu, N. Adaptive changes in chlorophyll content and photosynthetic features to low light in Physocarpus amurensis Maxim and Physocarpus opulifolius “Diabolo”. PeerJ 2016, 4, e2125. [Google Scholar] [CrossRef] [Green Version]
- Sui, X.-L.; Mao, S.-L.; Wang, L.-H.; Zhang, B.-X.; Zhang, Z.-X. Effect of Low Light on the Characteristics of Photosynthesis and Chlorophyll a Fluorescence During Leaf Development of Sweet Pepper. J. Integr. Agric. 2012, 11, 1633–1643. [Google Scholar] [CrossRef]
- Ryan, K.; Tay, M.; Martin, A.; McMinn, A.; Davy, S. Chlorophyll fluorescence imaging analysis of the responses of Antarctic bottom-ice algae to light and salinity during melting. J. Exp. Mar. Biol. Ecol. 2011, 399, 156–161. [Google Scholar] [CrossRef]
- Szabó, M.; Parker, K.; Guruprasad, S.; Kuzhiumparambil, U.; Lilley, R.M.; Tamburic, B.; Schliep, M.; Larkum, A.W.; Schreiber, U.; Raven, J.A.; et al. Photosynthetic acclimation of Nannochloropsis oculata investigated by multi-wavelength chlorophyll fluorescence analysis. Bioresour. Technol. 2014, 167, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Tarakanov, I.G.; Tovstyko, D.A.; Lomakin, M.P.; Shmakov, A.S.; Sleptsov, N.N.; Shmarev, A.N.; Litvinskiy, V.A.; Ivlev, A.A. Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants. Plants 2022, 11, 441. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.I. The UV-B Photoreceptor UVR8: From Structure to Physiology. Plant Cell 2014, 26, 21–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellmann, E. Specific Ultraviolet Effects In Plant Morphogenesis. Photochem. Photobiol. 1976, 24, 659–660. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.T.; Puthur, J.T. Amplification of abiotic stress tolerance potential in rice seedlings with a low dose of UV-B seed priming. Funct. Plant Biol. 2019, 46, 455. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Kacharava, N.; Chanishvili, S.; Badridze, G.; Chkhubianishvili, E.; Janukashvili, N. Effect of seed irradiation on the content of antioxidants in leaves of Kidney bean, Cabbage and Beet cultivars. Aust. J. Crop Sci. 2009, 3, 137. [Google Scholar]
- Lai, J.; Lim, P.; Wong, C.; Phang, S.; Beardall, J. Photosynthetic response and DNA mutation of tropical, temperate and polar Chlorella under short-term UVR stress. Polar Sci. 2018, 20, 35–44. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Li, Y.; Ma, L.; Bu, N.; Ma, C. Changes in photosynthesis, antioxidant enzymes and lipid peroxidation in soybean seedlings exposed to UV-B radiation and/or Cd. Plant Soil 2011, 352, 377–387. [Google Scholar] [CrossRef]
- Thomas, T.D.; Dinakar, C.; Puthur, J.T. Effect of UV-B priming on the abiotic stress tolerance of stress-sensitive rice seedlings: Priming imprints and cross-tolerance. Plant Physiol. Biochem. 2019, 147, 21–30. [Google Scholar] [CrossRef]
- Faseela, P.; Puthur, J.T. Intraspecific variation in sensitivity of high yielding rice varieties towards UV-B radiation. Physiol. Mol. Biol. Plants 2019, 25, 727–740. [Google Scholar] [CrossRef]
- Ek, N.; Kalaji, H.M.; EkmekÇİ, Y. Probing the photosynthetic efficiency of some European and Anatolian Scots pine populations under UV-B radiation using polyphasic chlorophyll a fluorescence transient. Photosynthetica 2020, 58, 468–478. [Google Scholar]
- Quan, J.; Song, S.; Abdulrashid, K.; Chai, Y.; Yue, M.; Liu, X. Separate and Combined Response to UV-B Radiation and Jasmonic Acid on Photosynthesis and Growth Characteristics of Scutellaria baicalensis. Int. J. Mol. Sci. 2018, 19, 1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, H.; Niu, G.; Gu, M. Pre-Harvest UV-B Radiation and Photosynthetic Photon Flux Density Interactively Affect Plant Photosynthesis, Growth, and Secondary Metabolites Accumulation in Basil (Ocimum basilicum) Plants. Agronomy 2019, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Kreslavski, V.D.; Strokina, V.V.; Khudyakova, A.Y.; Shirshikova, G.N.; Kosobryukhov, A.A.; Pashkovskiy, P.P.; Alwasel, S.; Allakhverdiev, S.I. Effect of high-intensity light and UV-B on photosynthetic activity and the expression of certain light-responsive genes in A. thaliana phyA and phyB mutants. Biochim. Biophys. Acta 2021, 1862, 148445. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, K.; Piwowarczyk, B.; Wysocka, A.; Wójtowicz, T.; Makowski, W.; Golemiec, E. Response of grass pea (Lathyrus sativus L.) photosynthetic apparatus to short-term intensive UV-A: Red radiation. Acta Physiol. Plant. 2019, 41, 168. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Hao, Z.; Anken, R.H.; Lu, J.; Liu, Y. Effects of UV-B radiation on photosynthesis activity of Wolffia arrhiza as probed by chlorophyll fluorescence transients. Adv. Space Res. 2010, 45, 839–845. [Google Scholar] [CrossRef]
- Vass, I.; Sass, L.; Spetea, C.; Bakou, A.; Ghanotakis, D.F.; Petrouleas, V. UV-B-Induced Inhibition of Photosystem II Electron Transport Studied by EPR and Chlorophyll Fluorescence. Impairment of Donor and Acceptor Side Components. Biochemistry 1996, 35, 8964–8973. [Google Scholar] [CrossRef]
- Vass, I.; Kirilovsky, D.; Etienne, A.-L. UV-B Radiation-Induced Donor- and Acceptor-Side Modifications of Photosystem II in the Cyanobacterium Synechocystis sp. PCC 6803. Biochemistry 1999, 38, 12786–12794. [Google Scholar] [CrossRef]
- Larkum, A.W.; Karge, M.; Reifarth, F.; Eckert, H.J.; Post, A.; Renger, G. Effect of monochromatic UV-B radiation on electron transfer reactions of Photosystem II. Photosynth. Res. 2001, 68, 49–60. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Grinberg, M.A.; Sukhov, V.; Vodeneev, V. Effect of ionizing radiation on physiological and molecular processes in plants. J. Environ. Radioact. 2019, 202, 8–24. [Google Scholar] [CrossRef]
- Chandorkar, K.; Clark, G. Physiological and morphological responses of Pinus strobus L. and Pinus sylvestris L. seedlings subjected to low-level continuous gamma irradiation at a radioactive waste disposal area. Environ. Exp. Bot. 1986, 26, 259–270. [Google Scholar] [CrossRef]
- Kim, J.H.; Moon, Y.R.; Lee, M.H.; Kim, J.H.; Wi, S.G.; Park, B.J.; Kim, C.S.; Chung, B.Y. Photosynthetic capacity of Arabidopsis plants at the reproductive stage tolerates gamma irradiation. J. Radiat. Res. 2011, 52, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Gomes, T.; Xie, L.; Brede, D.; Lind, O.-C.; Solhaug, K.A.; Salbu, B.; Tollefsen, K.E. Sensitivity of the green algae Chlamydomonas reinhardtii to gamma radiation: Photosynthetic performance and ROS formation. Aquat. Toxicol. 2017, 183, 1–10. [Google Scholar] [CrossRef]
- Vanhoudt, N.; Horemans, N.; Wannijn, J.; Nauts, R.; Van Hees, M.; Vandenhove, H. Primary stress responses in Ara-bidopsis thaliana exposed to gamma radiation. J. Environ. Radioact. 2014, 129, 1–6. [Google Scholar] [CrossRef]
- Sato, R.; Ito, H.; Tanaka, A. Chlorophyll b degradation by chlorophyll b reductase under high-light conditions. Photosynth. Res. 2015, 126, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Shi, M.; Huang, J.-Z.; Xu, J.; Wang, Z.-D.; Guo, D.-P. Regulation of photosynthetic performance and antioxidant capacity by 60Co γ-irradiation in Zizania latifolia plants. J. Environ. Radioact. 2013, 129, 33–42. [Google Scholar] [CrossRef]
- Lee, H.Y.; Baek, M.H.; Park, S.C.; Park, Y.I.; Kim, J.S. Effects of low dose γ-radiation on photosynthesis of red pepper (Capsicum annuum L.) and the reduction of photoinhibition. Korean J. Environ. Agric. 2002, 21, 83–89. [Google Scholar]
- Sobczak, A.; Sujkowska-Rybkowska, M.; Gajc-Wolska, J.; Kowalczyk, W.; Borucki, W.; Kalaji, H.M.; Kowalczyk, K. Photosynthetic Efficiency and Anatomical Structure of Pepper Leaf (Capsicum annuum L.) Transplants Grown under High-Pressure Sodium (HPS) and Light-Emitting Diode (LED) Supplementary Lighting Systems. Plants 2021, 10, 1975. [Google Scholar] [CrossRef]
- Darko, É.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, ply052. [Google Scholar] [CrossRef]
- Maiza, R.; Kurnia, D. The Influence of Light Wavelengths Toward the Growth of Brassica rapa L. J. Physics Conf. Ser. 2019, 1245, 012089. [Google Scholar] [CrossRef]
- Yang, B.; Zhou, X.; Xu, R.; Wang, J.; Lin, Y.; Pang, J.; Wu, S.; Zhong, F. Comprehensive analysis of photosynthetic characteristics and quality improvement of purple cabbage under different combinations of monochromatic light. Front. Plant Sci. 2016, 7, 1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf Morphology, Photosynthetic Performance, Chlorophyll Fluorescence, Stomatal Development of Lettuce (Lactuca sativa L.) Exposed to Different Ratios of Red Light to Blue Light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Evans, J. The Dependence of Quantum Yield on Wavelength and Growth Irradiance. Funct. Plant Biol. 1987, 14, 69–79. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [Green Version]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic perfor-mance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar]
- Kim, S.-J.; Hahn, E.-J.; Heo, J.-W.; Paek, K.-Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2003, 101, 143–151. [Google Scholar] [CrossRef]
- Zhen, S.; Van Iersel, M.W. Far-red light is needed for efficient photochemistry and photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-J.; Park, S.-Y.; Oh, M.-M. Growth and cell division of lettuce plants under various ratios of red to far-red light-emitting diodes. Hortic. Environ. Biotechnol. 2015, 56, 186–194. [Google Scholar] [CrossRef]
- Cao, K.; Yu, J.; Xu, D.; Ai, K.; Bao, E.; Zou, Z. Exposure to lower red to far-red light ratios improve tomato tolerance to salt stress. BMC Plant Biol. 2018, 18, 92. [Google Scholar] [CrossRef]
- Hogewoning, S.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the performance of cucumber (Cucumis sativus L.) seedlings by LED illumination. Sci. Rep. 2022, 12, 852. [Google Scholar] [CrossRef]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa × hybridabut does not affect time to flower opening. Physiol. Plant. 2012, 148, 146–159. [Google Scholar] [CrossRef]
- Wu, H. Effect of different light qualities on growth, pigment content, chlorophyll fluorescence, and antioxidant enzyme activity in the red alga Pyropia haitanensis (Bangiales, Rhodophyta). BioMed Res. Int. 2016, 2016, 7383918. [Google Scholar] [CrossRef] [Green Version]
- Vesk, M.; Jeffrey, S.W. Effect of blue-green light on photosynthetic pigments and chloroplast structure in unicellular marine algae from six classes 1. J. Phycol. 1977, 13, 280–288. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.N.; Yoshihara, T. Effect of green light wavelength and intensity on pho-tomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Wang, C.-J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.-D.; Liu, H.-X.; Wang, Y.-P.; Guo, J.-H. Induction of Drought Tolerance in Cucumber Plants by a Consortium of Three Plant Growth-Promoting Rhizobacterium Strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Qian, S.; Liu, Q.; Jin, C.; Zhou, X. The Response of Bio-Component Production of Nannochloris oculata to the Combinations of Monochromatic Light. J. Ocean Univ. China 2022, 21, 243–251. [Google Scholar] [CrossRef]
- Lili, M.E.N.G.; Kai, C.A.O.; Qian, S.U.N.; Zongchun, B.A.I.; Yi, Z.H.A.N.G. Effects of Different Ratios of Red Light and Far Red Light on Growth, Photosynthetic Characteristics and Quality of Mesembryanthemum crystallinum L. J. Nucl. Agric. Sci. 2022, 36, 226. [Google Scholar]
- Modarelli, G.C.; Paradiso, R.; Arena, C.; De Pascale, S.; Van Labeke, M.-C. High Light Intensity from Blue-Red LEDs Enhance Photosynthetic Performance, Plant Growth, and Optical Properties of Red Lettuce in Controlled Environment. Horticulturae 2022, 8, 114. [Google Scholar] [CrossRef]
- Escobar-Bravo, R.; Klinkhamer, P.G.L.; Leiss, K.A. Interactive Effects of UV-B Light with Abiotic Factors on Plant Growth and Chemistry, and Their Consequences for Defense against Arthropod Herbivores. Front. Plant Sci. 2017, 8, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, M.-Y.; Zhang, Y.-J.; Liu, L.-A.; Shi, L.; Ma, Q.-H.; Chow, W.S.; Jiang, C.-D. Do rapid photosynthetic responses protect maize leaves against photoinhibition under fluctuating light? Photosynth. Res. 2020, 149, 57–68. [Google Scholar] [CrossRef]
- Charles, F.; Nilprapruck, P.; Roux, D.; Sallanon, H. Visible light as a new tool to maintain fresh-cut lettuce post-harvest quality. Postharvest Biol. Technol. 2018, 135, 51–56. [Google Scholar] [CrossRef]
- Li, Y.-T.; Yang, C.; Zhang, Z.-S.; Zhao, S.-J.; Gao, H.-Y. Photosynthetic acclimation strategies in response to intermittent exposure to high light intensity in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2020, 181, 104275. [Google Scholar] [CrossRef]
- Ayalon, I.; De Barros Marangoni, L.F.; Benichou, J.I.; Avisar, D.; Levy, O. Red Sea corals under Artificial Light Pol-lution at Night (ALAN) undergo oxidative stress and photosynthetic impairment. Glob. Change Biol. 2019, 25, 4194–4207. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Burkart, S. Photosynthesis and high light stress. Bulg. J. Plant Physiol 1999, 25, 3–16. [Google Scholar]
- Alvino, A.; Sorrentino, G. Highlight Effects on Chlorophyll Fluorescence and Light Interception in Lilium. Adv. Hortic. Sci. 1996, 10, 70–78. [Google Scholar] [CrossRef]
- Kandler, O.; Sironval, C. Photoxidation processes in normal green Chlorella cells II. Effects on metabolism. Biochim. Biophys. Acta 1959, 33, 207–215. [Google Scholar] [CrossRef]
- Carvalho, L.; Coito, J.L.; Gonçalves, E.M.F.; Chaves, M.M.; Amâncio, S. Differential physiological response of the grapevine varieties Touriga Nacional and Trincadeira to combined heat, drought and light stresses. Plant Biol. 2015, 18, 101–111. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
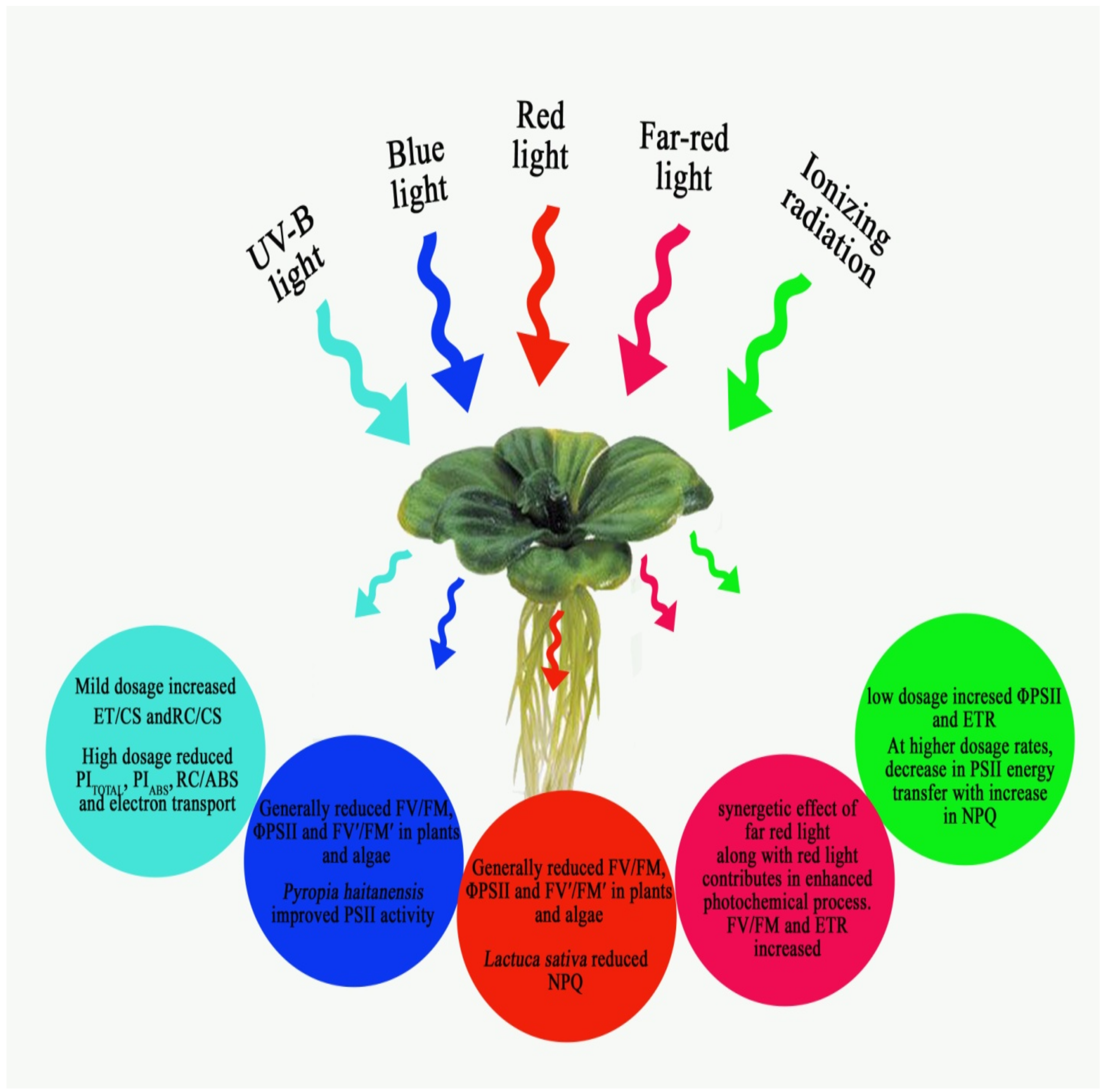{kind=link}
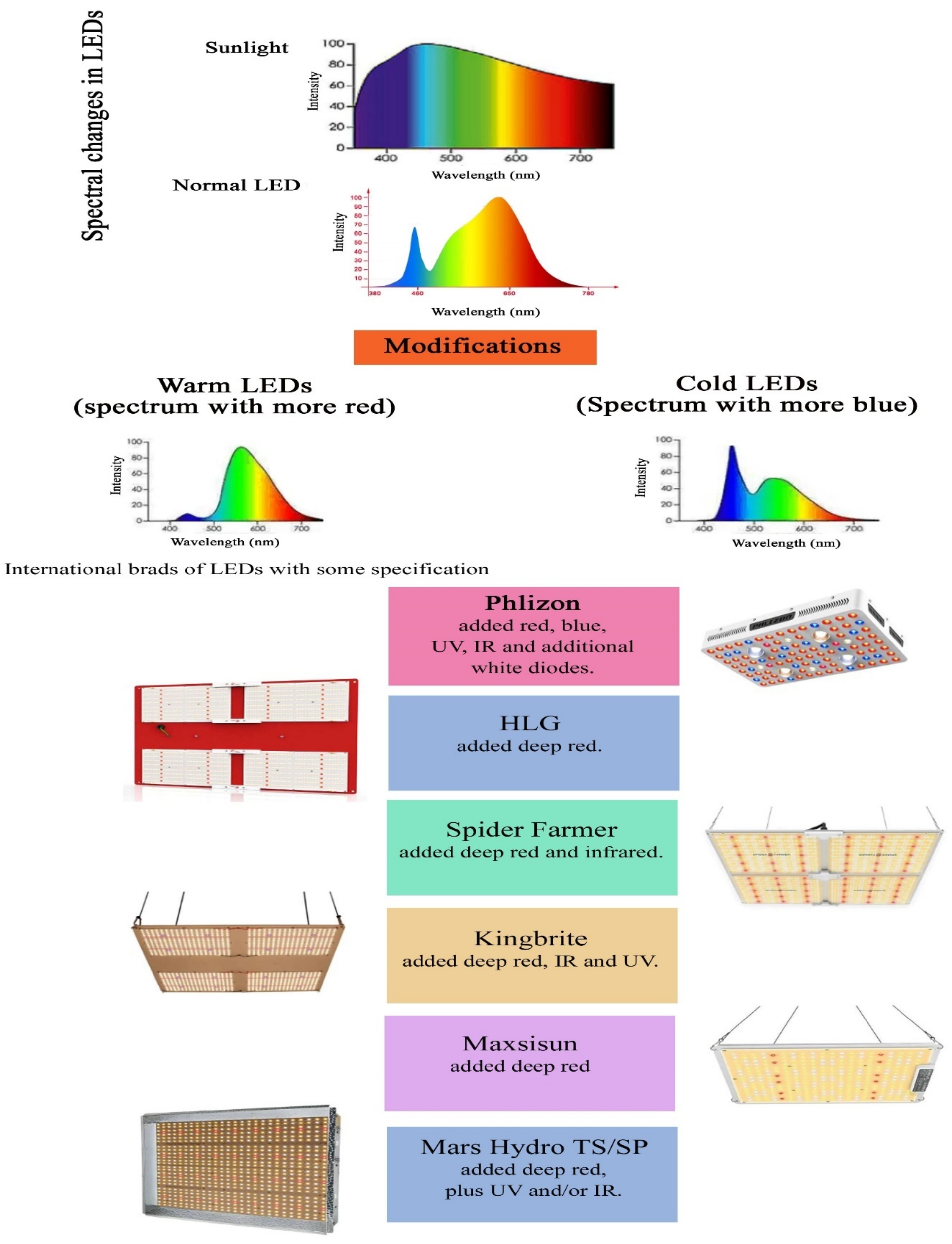{kind=link}
| Plant | Dose | Effect | Reference |
|---|---|---|---|
| Oryza sativa L. | 2000 µmol m−2 s−1 | Increased ABS and DIo/RC | [5] |
| Chlamydomonas reinhardtti | 2000 µmol m−2 s−1 | Decreased photosynthetic efficiency and increased photo-oxidative damage | [36] |
| Dunellia salina | 1000 µmol m−2 s−1 | Induced photo-damage to cells and increased free hydroxyl radicals | [40] |
| Glycine max (L.) Merr. | 2000 µmol m−2 s−1 | Decline in photo-chemical quenching coefficient and FV/FM | [52] |
| Oryza sativa L. | 2000 µmol m−2 s−1 | Reduction in photo-chemical efficiency accompanied by high emission of dissipated energy leading to PS I and PS II damage | [44] |
| Lillium sp. | 2000 µmol m−2 s−1 | Energy dissipation level increased DI and level of excess light absorbed by FV/FM | [53] |
| Chlorella sp. | 2000 µmol m−2 s−1 | Inhibited photosynthesis and ligh-dependent oxygen bleaching | [54] |
| Grapevine | 2000 µmol m−2 s−1 | Increased DIo/RC, reduced photosynthesis | [55] |
| Arabidopsis thaliana (L.) Heynh. | 1000 µmol m−2 s−1 | Degradation of chl b by an isozyme chl –b reductase | [56] |
| Hordeum vulgare L. | 1800 µmol m−2 s−1 | Severe damaging effect on FV/FM and PIabs | [48] |
| Plant | Intensity of Light | Response in Important JIP parameter | References |
|---|---|---|---|
| Oryza sativa | UV-B (4–6 kJ m−2 d−1) | RC:CS, ABS:CS, ETo:CSM, TR:CS, ET:CS increased | [70] |
| O. sativa | UV-B (28 kJ m−2 d−1) | ABS/RC and ETo/RC decreased; DIo/RC increased | [5] |
| Pinus sylvestris L. | UV-B (2.8 kJ m−2 d−1 for 2 days) | PITotal, PIABS, RC/ABS, REo/RC decreased; ABS/RC, ETo/RC, DIo/RC increased | [72] |
| Scutellaria baicalensis | UV-B (10.30 kJ m−2 d−1 for 15 days) | Pmax and ETRmax decreased; FV/FM no change | [73] |
| Microalgae Chlorella sp. | UV-A (8.54 wm−2) + UV-B (1.17 wm−2) | FV/FM and rETRm decreased | [68] |
| Glycine max | UV-A (7.1 kJ m−2 d−1) | FV/FM and Fv/Fo no change | [69] |
| Wolffia arrhiza | UV-B (4 wm−2) | φpo, φEo, φo decreased; RC/ABS and RC/CS increased | [77] |
| Spinacia oleracea | UV-B (50 µEm−2 s−1) | FV/FM decreased | [78] |
| Cyanobacterium Synechocystis sp. | UV-B (2.4 wm−2) | Fmax increased | [79] |
| Spinacia oleracea | UV-B (5 wm−2) | F(t)/Fo decreased | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janeeshma, E.; Johnson, R.; Amritha, M.S.; Noble, L.; Aswathi, K.P.R.; Telesiński, A.; Kalaji, H.M.; Auriga, A.; Puthur, J.T. Modulations in Chlorophyll a Fluorescence Based on Intensity and Spectral Variations of Light. Int. J. Mol. Sci. 2022, 23, 5599. https://doi.org/10.3390/ijms23105599
Janeeshma E, Johnson R, Amritha MS, Noble L, Aswathi KPR, Telesiński A, Kalaji HM, Auriga A, Puthur JT. Modulations in Chlorophyll a Fluorescence Based on Intensity and Spectral Variations of Light. International Journal of Molecular Sciences. 2022; 23(10):5599. https://doi.org/10.3390/ijms23105599
Chicago/Turabian StyleJaneeshma, Edappayil, Riya Johnson, M. S. Amritha, Louis Noble, K. P. Raj Aswathi, Arkadiusz Telesiński, Hazem M. Kalaji, Alicja Auriga, and Jos T. Puthur. 2022. "Modulations in Chlorophyll a Fluorescence Based on Intensity and Spectral Variations of Light" International Journal of Molecular Sciences 23, no. 10: 5599. https://doi.org/10.3390/ijms23105599