
The Role of miRNAs in Dexmedetomidine’s Neuroprotective Effects against Brain Disorders

 , , , and
, , , and
Abstract
:1. Introduction
2. Pharmacological Features of Dexmedetomidine
3. Evidence of Dexmedetomidine as a Neuroprotective Agent
4. Short Overview of miRNAs as Therapeutic and Diagnostic Tools in Brain Disorders
5. Regulation Production of miRNAs after Dexmedetomidine Treatment
5.1. miRNA Biogenesis
5.2. DEX-Dependent Transcription Controlling Systems of the miRNAs
6. Translation Relevance of miRNA in Dexmedetomidine-Mediated Neuroprotection
6.1. Neuroprotective Profile of Dexmedetomidine in Brain Disorders
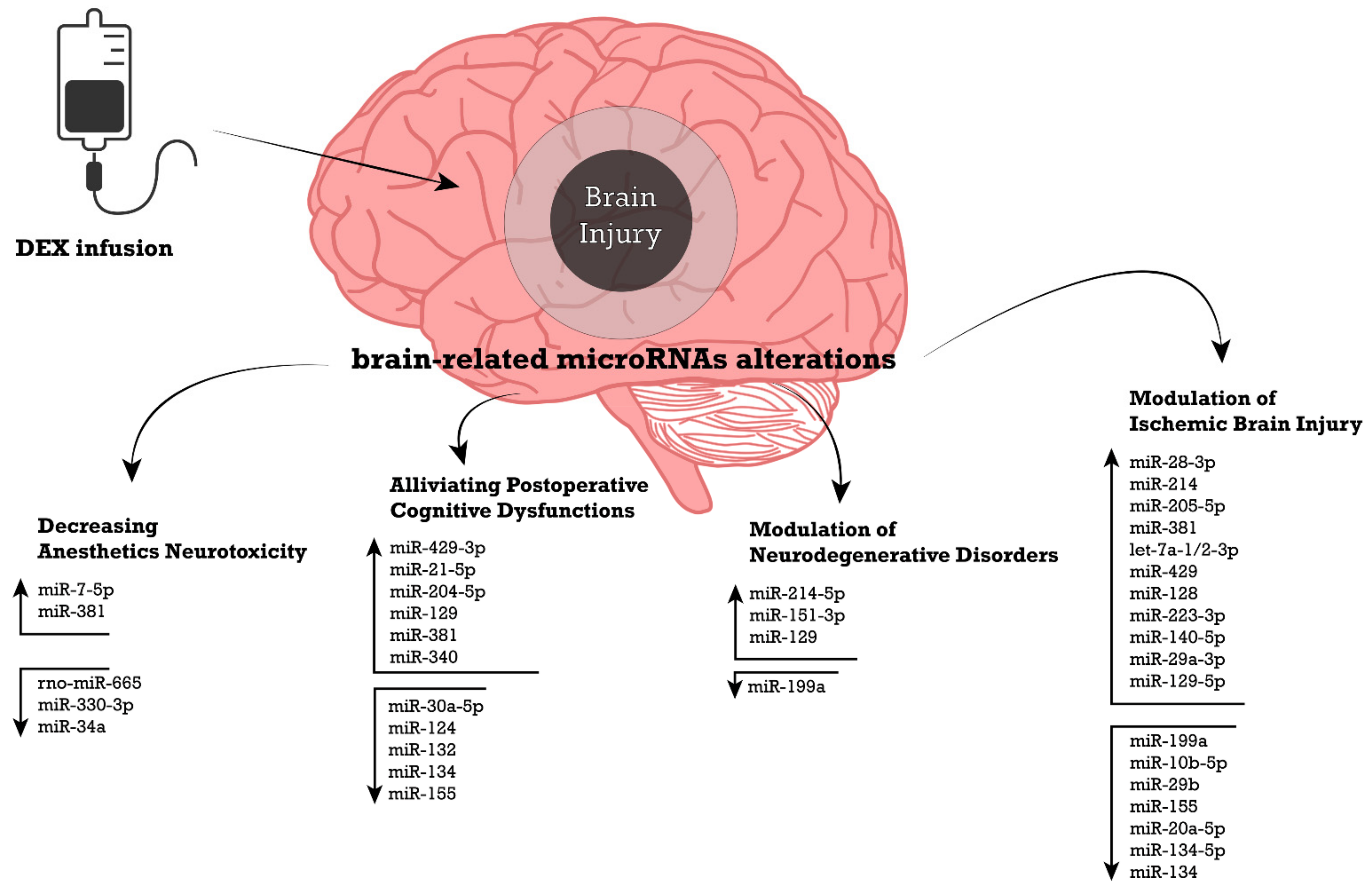
6.2. Modulation of Ischemic Brain Injury
6.3. Decreasing the Neurotoxicity of Anesthetics
6.4. DEX-Mediated Neuroprotection against Postoperative Cognitive Dysfunctions
6.5. Modulation of Neurodegenerative Disorders
7. MiRNAs in Pharmacological Response of Dexmedetomidine and Drug Response Variability
7.1. Involvement of MiRNAs in Drug Response Variability
7.2. MiRNAs in Dexmedetomidine’s Pharmacological Profiles
8. Potential Clinical Applications and Therapeutic Targets
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farrell, D.; Bendo, A.A. Perioperative Management of Severe Traumatic Brain Injury: What Is New? Curr. Anesthesiol. Rep. 2018, 8, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkkinen, M.G.; Kim, M.-O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallin, M.T.; Culpepper, W.J.; Nichols, E.; Bhutta, Z.A.; Gebrehiwot, T.T.; Hay, S.I.; Khalil, I.A.; Krohn, K.J.; Liang, X.; Naghavi, M.; et al. Global, Regional, and National Burden of Multiple Sclerosis 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, J.; Nutma, E.; van der Valk, P.; Amor, S. Inflammation in CNS Neurodegenerative Diseases. Immunology 2018, 154, 204–219. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.-S.; Knapp, D.J.; Crews, F.T. Systemic LPS Causes Chronic Neuroinflammation and Progressive Neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Fidalgo, A.R.; Cibelli, M.; White, J.P.M.; Nagy, I.; Maze, M.; Ma, D. Systemic Inflammation Enhances Surgery-Induced Cognitive Dysfunction in Mice. Neurosci. Lett. 2011, 498, 63–66. [Google Scholar] [CrossRef]
- He, W.; Zhang, Z.; Sha, X. Nanoparticles-Mediated Emerging Approaches for Effective Treatment of Ischemic Stroke. Biomaterials 2021, 277, 121111. [Google Scholar] [CrossRef]
- Hong, H.-Y.; Choi, J.S.; Kim, Y.J.; Lee, H.Y.; Kwak, W.; Yoo, J.; Lee, J.-T.; Kwon, T.-H.; Kim, I.-S.; Han, H.-S.; et al. Detection of Apoptosis in a Rat Model of Focal Cerebral Ischemia Using a Homing Peptide Selected from in Vivo Phage Display. J. Control. Release Off. J. Control. Release Soc. 2008, 131, 167–172. [Google Scholar] [CrossRef]
- Paudyal, A.; Ghinea, F.S.; Driga, M.P.; Fang, W.-H.; Alessandri, G.; Combes, L.; Degens, H.; Slevin, M.; Hermann, D.M.; Popa-Wagner, A. P5 Peptide-Loaded Human Adipose-Derived Mesenchymal Stem Cells Promote Neurological Recovery After Focal Cerebral Ischemia in a Rat Model. Transl. Stroke Res. 2021, 12, 125–135. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An Update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef]
- Chamorro, Á.; Lo, E.H.; Renú, A.; van Leyen, K.; Lyden, P.D. The Future of Neuroprotection in Stroke. J. Neurol. Neurosurg. Psychiatry 2021, 92, 129–135. [Google Scholar] [CrossRef]
- Bozorgi, H.; Zamani, M.; Motaghi, E.; Eslami, M. Dexmedetomidine as an Analgesic Agent with Neuroprotective Properties: Experimental and Clinical Aspects. J. Pain Palliat. Care Pharmacother. 2021, 35, 215–225. [Google Scholar] [CrossRef]
- Liaquat, Z.; Xu, X.; Zilundu, P.L.M.; Fu, R.; Zhou, L. The Current Role of Dexmedetomidine as Neuroprotective Agent: An Updated Review. Brain Sci. 2021, 11, 846. [Google Scholar] [CrossRef]
- Wang, Y.; Han, R.; Zuo, Z. Dexmedetomidine-Induced Neuroprotection: Is It Translational? Transl. Perioper. Pain Med. 2016, 1, 15–19. [Google Scholar]
- Hoffman, W.E.; Kochs, E.; Werner, C.; Thomas, C.; Albrecht, R.F. Dexmedetomidine Improves Neurologic Outcome from Incomplete Ischemia in the Rat. Reversal by the Alpha 2-Adrenergic Antagonist Atipamezole. Anesthesiology 1991, 75, 328–332. [Google Scholar] [CrossRef]
- Cheng, J.; Zhu, P.; Qin, H.; Li, X.; Yu, H.; Yu, H.; Peng, X. Dexmedetomidine Attenuates Cerebral Ischemia/Reperfusion Injury in Neonatal Rats by Inhibiting TLR4 Signaling. J. Int. Med. Res. 2018, 46, 2925–2932. [Google Scholar] [CrossRef]
- Zhu, Y.-M.; Wang, C.-C.; Chen, L.; Qian, L.-B.; Ma, L.-L.; Yu, J.; Zhu, M.-H.; Wen, C.-Y.; Yu, L.-N.; Yan, M. Both PI3K/Akt and ERK1/2 Pathways Participate in the Protection by Dexmedetomidine against Transient Focal Cerebral Ischemia/Reperfusion Injury in Rats. Brain Res. 2013, 1494, 1–8. [Google Scholar] [CrossRef]
- Luo, C.; Ouyang, M.-W.; Fang, Y.-Y.; Li, S.-J.; Zhou, Q.; Fan, J.; Qin, Z.-S.; Tao, T. Dexmedetomidine Protects Mouse Brain from Ischemia-Reperfusion Injury via Inhibiting Neuronal Autophagy through Up-Regulating HIF-1α. Front. Cell. Neurosci. 2017, 11, 197. [Google Scholar] [CrossRef] [Green Version]
- Lankadeva, Y.R.; Ma, S.; Iguchi, N.; Evans, R.G.; Hood, S.G.; Farmer, D.G.S.; Bailey, S.R.; Bellomo, R.; May, C.N. Dexmedetomidine Reduces Norepinephrine Requirements and Preserves Renal Oxygenation and Function in Ovine Septic Acute Kidney Injury. Kidney Int. 2019, 96, 1150–1161. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Tüfekci, K.U.; Oner, M.G.; Meuwissen, R.L.J.; Genç, S. The Role of MicroRNAs in Human Diseases. Methods Mol. Biol. 2014, 1107, 33–50. [Google Scholar] [CrossRef]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar] [PubMed]
- Chen, Y.-M.; He, X.-Z.; Wang, S.-M.; Xia, Y. δ-Opioid Receptors, MicroRNAs, and Neuroinflammation in Cerebral Ischemia/Hypoxia. Front. Immunol. 2020, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Hoss, A.G.; Labadorf, A.; Beach, T.G.; Latourelle, J.C.; Myers, R.H. MicroRNA Profiles in Parkinson’s Disease Prefrontal Cortex. Front. Aging Neurosci. 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuzziello, N.; Vilardo, L.; Pelucchi, P.; Consiglio, A.; Liuni, S.; Trojano, M.; Liguori, M. Investigating the Role of MicroRNA and Transcription Factor Co-Regulatory Networks in Multiple Sclerosis Pathogenesis. Int. J. Mol. Sci. 2018, 19, 3652. [Google Scholar] [CrossRef] [Green Version]
- Petrescu, G.E.D.; Sabo, A.A.; Torsin, L.I.; Calin, G.A.; Dragomir, M.P. MicroRNA Based Theranostics for Brain Cancer: Basic Principles. J. Exp. Clin. Cancer Res. 2019, 38, 231. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Zhu, Y.; He, G.; Ni, H.; Liu, C.; Ma, L.; Zhang, L.; Shi, D. Dexmedetomidine Attenuates Neuroinflammation In LPS-Stimulated BV2 Microglia Cells Through Upregulation Of MiR-340. Drug Des. Devel. Ther. 2019, 13, 3465–3475. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wu, Y.; Li, N.; Wang, Z. Up-Regulation of MiRNA-151-3p Enhanced the Neuroprotective Effect of Dexmedetomidine against β-Amyloid by Targeting DAPK-1 and TP53. Exp. Mol. Pathol. 2021, 118, 104587. [Google Scholar] [CrossRef]
- Yang, J.-J.; Zhao, Y.-H.; Yin, K.-W.; Zhang, X.-Q.; Liu, J. Dexmedetomidine Inhibits Inflammatory Response and Oxidative Stress through Regulating MiR-205-5p by Targeting HMGB1 in Cerebral Ischemic/Reperfusion. Immunopharmacol. Immunotoxicol. 2021, 43, 478–486. [Google Scholar] [CrossRef]
- Xue, Y.; Xu, T.; Jiang, W. Dexmedetomidine Protects PC12 Cells from Ropivacaine Injury through MiR-381/LRRC4 /SDF-1/CXCR4 Signaling Pathway. Regen. Ther. 2020, 14, 322–329. [Google Scholar] [CrossRef]
- Ciccacci, C.; Rufini, S.; Politi, C.; Novelli, G.; Forte, V.; Borgiani, P. Could MicroRNA Polymorphisms Influence Warfarin Dosing? A Pharmacogenetics Study on Mir133 Genes. Thromb. Res. 2015, 136, 367–370. [Google Scholar] [CrossRef]
- Cai, X.; Li, B.; Wei, W.; Guan, Y.; Bai, X.; Huang, M.; Huang, Y.; Rong, L.; Song, X.; Zhong, G. Circulating MicroRNA-30a-5p, MicroRNA-101-3p, MicroRNA-140-3p and MicroRNA-141-3p as Potential Biomarkers for Dexmedetomidine Response in Pediatric Patients. Eur. J. Clin. Pharmacol. 2021, 77, 1853–1859. [Google Scholar] [CrossRef]
- Scott-Warren, V.L.; Sebastian, J. Dexmedetomidine: Its Use in Intensive Care Medicine and Anaesthesia. BJA Educ. 2016, 16, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Yuen, V.M.Y.; Goulay-Dufay, S.; Kwok, P.C.L. Pharmacokinetics and Pharmacodynamics of Dexmedetomidine. Drug Dev. Ind. Pharm. 2016, 42, 1917–1927. [Google Scholar] [CrossRef]
- Shehabi, Y.; Howe, B.D.; Bellomo, R.; Arabi, Y.M.; Bailey, M.; Bass, F.E.; Bin Kadiman, S.; McArthur, C.J.; Murray, L.; Reade, M.C.; et al. Early Sedation with Dexmedetomidine in Critically Ill Patients. N. Engl. J. Med. 2019, 380, 2506–2517. [Google Scholar] [CrossRef]
- Kim, K.H. Safe Sedation and Hypnosis Using Dexmedetomidine for Minimally Invasive Spine Surgery in a Prone Position. Korean J. Pain 2014, 27, 313–320. [Google Scholar] [CrossRef]
- Yuan, D.; Liu, Z.; Kaindl, J.; Maeda, S.; Zhao, J.; Sun, X.; Xu, J.; Gmeiner, P.; Wang, H.-W.; Kobilka, B.K. Activation of the A2B Adrenoceptor by the Sedative Sympatholytic Dexmedetomidine. Nat. Chem. Biol. 2020, 16, 507–512. [Google Scholar] [CrossRef]
- Weerink, M.A.S.; Struys, M.M.R.F.; Hannivoort, L.N.; Barends, C.R.M.; Absalom, A.R.; Colin, P. Clinical Pharmacokinetics and Pharmacodynamics of Dexmedetomidine. Clin. Pharmacokinet. 2017, 56, 893–913. [Google Scholar] [CrossRef] [Green Version]
- De Wolf, A.M.; Fragen, R.J.; Avram, M.J.; Fitzgerald, P.C.; Rahimi-Danesh, F. The Pharmacokinetics of Dexmedetomidine in Volunteers with Severe Renal Impairment. Anesth. Analg. 2001, 93, 1205–1209. [Google Scholar] [CrossRef]
- Zhong, W.; Zhang, Y.; Zhang, M.-Z.; Huang, X.-H.; Li, Y.; Li, R.; Liu, Q.-W. Pharmacokinetics of Dexmedetomidine Administered to Patients with End-Stage Renal Failure and Secondary Hyperparathyroidism Undergoing General Anaesthesia. J. Clin. Pharm. Ther. 2018, 43, 414–421. [Google Scholar] [CrossRef]
- Ahsan, M.Z.; Khan, F.U.; Zhao, M.-J.; Wang, Y.-X. Synergistic Interaction between Butorphanol and Dexmedetomidine in Antinociception. Eur. J. Pharm. Sci. 2020, 149, 105322. [Google Scholar] [CrossRef]
- Saccomanni, G.; Giorgi, M.; Del Carlo, S.; Manera, C.; Saba, A.; Macchia, M. Simultaneous Detection and Quantification of Parecoxib and Valdecoxib in Canine Plasma by HPLC with Spectrofluorimetric Detection: Development and Validation of a New Methodology. Anal. Bioanal. Chem. 2011, 401, 1677–1684. [Google Scholar] [CrossRef]
- Fenton, C.; Keating, G.M.; Wagstaff, A.J. Valdecoxib: A Review of Its Use in the Management of Osteoarthritis, Rheumatoid Arthritis, Dysmenorrhoea and Acute Pain. Drugs 2004, 64, 1231–1261. [Google Scholar] [CrossRef]
- Du, X.; Song, F.; Zhang, X.; Ma, S. Protective Efficacy of Combined Use of Parecoxib and Dexmedetomidine on Postoperative Hyperalgesia and Early Cognitive Dysfunction after Laparoscopic Cholecystectomy for Elderly Patients. Acta Cir. Bras. 2019, 34, e201900905. [Google Scholar] [CrossRef]
- Hu, J.; Lv, B.; Guo, W.; Wang, B.; Miao, D.; Qiu, X.; Chen, X. Effects of Dexmedetomidine on the Pharmacokinetics of Parecoxib and Its Metabolite Valdecoxib in Beagles by UPLC-MS/MS. BioMed Res. Int. 2020, 2020, e1563874. [Google Scholar] [CrossRef]
- Kaye, A.D.; Chernobylsky, D.J.; Thakur, P.; Siddaiah, H.; Kaye, R.J.; Eng, L.K.; Harbell, M.W.; Lajaunie, J.; Cornett, E.M. Dexmedetomidine in Enhanced Recovery After Surgery (ERAS) Protocols for Postoperative Pain. Curr. Pain Headache Rep. 2020, 24, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Q.; Wang, P.; Jin, W.; Zhong, C.; Ge, Z.; Xu, K. The Effect of Dexmedetomidine as a Sedative Agent for Mechanically Ventilated Patients With Sepsis: A Systematic Review and Meta-Analysis. Front. Med. 2021, 8, 2444. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Hu, B.; Li, Z.; Li, J. Dexmedetomidine Controls Systemic Cytokine Levels through the Cholinergic Anti-Inflammatory Pathway. Inflammation 2014, 37, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, U.; Sarkar, M.E.; Kumar, A.; Gupta, A.; Agarwal, A.; Singh, T.K.; Badada, V.; Dwivedi, Y. Comparison of Clonidine and Dexmedetomidine for Short-Term Sedation of Intensive Care Unit Patients. Indian J. Crit. Care Med. 2014, 18, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Reddy, V.S.; Shaik, N.A.; Donthu, B.; Reddy Sannala, V.K.; Jangam, V. Intravenous Dexmedetomidine versus Clonidine for Prolongation of Bupivacaine Spinal Anesthesia and Analgesia: A Randomized Double-Blind Study. J. Anaesthesiol. Clin. Pharmacol. 2013, 29, 342–347. [Google Scholar] [CrossRef]
- Abdallah, F.W.; Brull, R. Facilitatory Effects of Perineural Dexmedetomidine on Neuraxial and Peripheral Nerve Block: A Systematic Review and Meta-Analysis. Br. J. Anaesth. 2013, 110, 915–925. [Google Scholar] [CrossRef] [Green Version]
- Keating, G.M. Dexmedetomidine: A Review of Its Use for Sedation in the Intensive Care Setting. Drugs 2015, 75, 1119–1130. [Google Scholar] [CrossRef]
- Giovannitti, J.A.; Thoms, S.M.; Crawford, J.J. Alpha-2 Adrenergic Receptor Agonists: A Review of Current Clinical Applications. Anesth. Prog. 2015, 62, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Jeong, J.S.; Park, H.; Sung, K.-S.; Choi, S.J.; Gwak, M.S.; Kim, G.S.; Hahm, T.S.; Ko, J.S. Postoperative Pain Control after the Use of Dexmedetomidine and Propofol to Sedate Patients Undergoing Ankle Surgery under Spinal Anesthesia: A Randomized Controlled Trial. J. Pain Res. 2019, 12, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Yuki, K. The Immunomodulatory Mechanism of Dexmedetomidine. Int. Immunopharmacol. 2021, 97, 107709. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.; Tian, S.; Wang, H.; Wu, H.; Zhang, A.; Gao, C. Anti-Inflammatory Effects of Perioperative Dexmedetomidine Administered as an Adjunct to General Anesthesia: A Meta-Analysis. Sci. Rep. 2015, 5, 12342. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xiao, F.; Zhang, J.; Wang, X.; Ying, J.; Wei, G.; Chen, S.; Huang, X.; Yu, W.; Liu, X.; et al. Dexmedetomidine Mitigated NLRP3-Mediated Neuroinflammation via the Ubiquitin-Autophagy Pathway to Improve Perioperative Neurocognitive Disorder in Mice. Front. Pharmacol. 2021, 12, 1143. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Guan, W.; Zhao, Y.; Wang, C.; Song, M.; Yao, Y.; Yang, T.; Fan, H. Dexmedetomidine Ameliorates Lipopolysaccharide-Induced Acute Kidney Injury in Rats by Inhibiting Inflammation and Oxidative Stress via the GSK-3β/Nrf2 Signaling Pathway. J. Cell. Physiol. 2019, 234, 18994–19009. [Google Scholar] [CrossRef]
- Tas, N.; Altinbas, A.; Noyan, T.; Kokturk, S.; Ayhan, S.; Canakci, E. Acute Acetaminophene-Induced Hepatotoxicity and Nephrotoxicity; Therapeutic Effect of Dexmedetomidine. Bratisl. Lek. Listy 2019, 120, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.; Sanfilippo, F.; Arnemann, P.; Hessler, M.; Kampmeier, T.G.; D’Egidio, A.; Orecchioni, A.; Santonocito, C.; Frati, G.; Greco, E.; et al. The Effect of Propofol and Dexmedetomidine Sedation on Norepinephrine Requirements in Septic Shock Patients: A Crossover Trial. Crit. Care Med. 2019, 47, e89. [Google Scholar] [CrossRef] [PubMed]
- Bafna, U.; Sharma, P.; Singhal, R.K.; Gurjar, S.S.; Bhargava, S.K. Comparison of Hypotensive Properties of Dexmedetomidine versus Clonidine for Induced Hypotension during Functional Endoscopic Sinus Surgery: A Randomised, Double-Blind Interventional Study. Indian J. Anaesth. 2021, 65, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Cioccari, L.; Luethi, N.; Bailey, M.; Shehabi, Y.; Howe, B.; Messmer, A.S.; Proimos, H.K.; Peck, L.; Young, H.; Eastwood, G.M.; et al. The Effect of Dexmedetomidine on Vasopressor Requirements in Patients with Septic Shock: A Subgroup Analysis of the Sedation Practice in Intensive Care Evaluation [SPICE III] Trial. Crit. Care 2020, 24, 441. [Google Scholar] [CrossRef]
- Aouad, M.T.; Zeeni, C.; Al Nawwar, R.; Siddik-Sayyid, S.M.; Barakat, H.B.; Elias, S.; Yazbeck Karam, V.G. Dexmedetomidine for Improved Quality of Emergence From General Anesthesia: A Dose-Finding Study. Anesth. Analg. 2019, 129, 1504–1511. [Google Scholar] [CrossRef]
- Li, N.; Ouyang, B.S.; Liu, L.; Lin, C.S.; Xing, D.D.; Liu, J. Dexmedetomidine Protected COPD-Induced Lung Injury by Regulating MiRNA-146a. Bratisl. Lek. Listy 2016, 117, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Li, J.; Fan, Q.; Zhao, X.; Chen, K. Dexmedetomidine Alleviates Myocardial Ischemia/Reperfusion-Induced Injury and Ca2+ Overload via the MicroRNA-346-3p/CaMKIId Axis. Int. J. Cardiol. 2021, 338, 185–195. [Google Scholar] [CrossRef]
- Ma, X.-G.; Liu, Y.; Xue, M.-X. [Dexmedetomidine alleviates hepatic ischemia-reperfusion injury by regulating MALAT1/miR-126-5p/HMGB1 axis]. Sheng Li Xue Bao 2021, 73, 253–262. [Google Scholar]
- Zheng, T.; Zheng, C.; Gao, F.; Huang, F.; Hu, B.; Zheng, X. Correction to: Dexmedetomidine Suppresses Bupivacaine-Induced Parthanatos in Human SH-SY5Y Cells via the MiR-7-5p/PARP1 Axis-Mediated ROS. Naunyn. Schmiedebergs Arch. Pharmacol. 2021, 394, 1055–1056. [Google Scholar] [CrossRef]
- Wei, W.; Sun, Z.; He, S.; Zhang, W.; Chen, S. Protective Role of Dexmedetomidine against Sevoflurane-Induced Postoperative Cognitive Dysfunction via the MicroRNA-129/TLR4 Axis. J. Clin. Neurosci. 2021, 92, 89–97. [Google Scholar] [CrossRef]
- Hu, G.; Shi, Z.; Shao, W.; Xu, B. MicroRNA-214-5p Involves in the Protection Effect of Dexmedetomidine against Neurological Injury in Alzheimer’s Disease via Targeting the Suppressor of Zest 12. Brain Res. Bull. 2022, 178, 164–172. [Google Scholar] [CrossRef]
- Huang, W.; Xiao, F.; Huang, W.; Wei, Q.; Li, X. MicroRNA-29a-3p Strengthens the Effect of Dexmedetomidine on Improving Neurologic Damage in Newborn Rats with Hypoxic-Ischemic Brain Damage by Inhibiting HDAC4. Brain Res. Bull. 2021, 167, 71–79. [Google Scholar] [CrossRef]
- Fang, H.; Li, H.-F.; Yan, J.-Y.; Yang, M.; Zhang, J.-P. Dexmedetomidine-up-Regulated MicroRNA-381 Exerts Anti-Inflammatory Effects in Rats with Cerebral Ischaemic Injury via the Transcriptional Factor IRF4. J. Cell. Mol. Med. 2021, 25, 2098–2109. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Zhang, Y.; Cai, D.-S. Dexmedetomidine Ameliorates Postoperative Cognitive Dysfunction via the MicroRNA-381-Mediated EGR1/P53 Axis. Mol. Neurobiol. 2021, 58, 5052–5066. [Google Scholar] [CrossRef]
- Kho, W.; von Haefen, C.; Paeschke, N.; Nasser, F.; Endesfelder, S.; Sifringer, M.; González-López, A.; Lanzke, N.; Spies, C.D. Dexmedetomidine Restores Autophagic Flux, Modulates Associated MicroRNAs and the Cholinergic Anti-Inflammatory Pathway upon LPS-Treatment in Rats. J. Neuroimmune Pharmacol. 2021. [Google Scholar] [CrossRef]
- Paeschke, N.; Von Haefen, C.; Endesfelder, S.; Sifringer, M.; Spies, C.D. Dexmedetomidine Prevents Lipopolysaccharide-Induced MicroRNA Expression in the Adult Rat Brain. Int. J. Mol. Sci. 2017, 18, 1830. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhu, X.; Yu, H.; Wang, W.; Wang, Y.; Li, F.; Teng, J. Regulatory Role of MicroRNA-320 during off-Pump Coronary Artery Bypass Grafting with Dexmedetomidine Adjunct Anesthesia. Exp. Ther. Med. 2021, 22, 1201. [Google Scholar] [CrossRef]
- Sun, L.; Zhou, J.; Sun, C. MicroRNA-211-5p Enhances Analgesic Effect of Dexmedetomidine on Inflammatory Visceral Pain in Rats by Suppressing ERK Signaling. J. Mol. Neurosci. 2019, 68, 19–28. [Google Scholar] [CrossRef]
- Wang, L.; Tang, S.; Wang, Z.; Chen, H.; Rajcha, S.S.; Qian, J. The Administration of Dexmedetomidine Changes MicroRNA Expression Profiling of Rat Hearts. Biomed. Pharmacother. 2019, 120, 109463. [Google Scholar] [CrossRef]
- Pardo, P.S.; Hajira, A.; Boriek, A.M.; Mohamed, J.S. MicroRNA-434-3p Regulates Age-Related Apoptosis through EIF5A1 in the Skeletal Muscle. Aging 2017, 9, 1012–1029. [Google Scholar] [CrossRef] [Green Version]
- Wong, L.L.; Rademaker, M.T.; Saw, E.L.; Lew, K.S.; Ellmers, L.J.; Charles, C.J.; Richards, A.M.; Wang, P. Identification of Novel MicroRNAs in the Sheep Heart and Their Regulation in Heart Failure. Sci. Rep. 2017, 7, 8250. [Google Scholar] [CrossRef]
- Wang, J.; Dai, E.; Su, Z. Dexmedetomidine Attenuates Lipopolysaccharide-Induced Acute Lung Injury by Regulation of MicroRNA-223-3p and HDAC4; Social Science Research Network: Rochester, NY, USA, 2020. [Google Scholar]
- Fang, H.; Li, H.-F.; Yang, M.; Wang, R.-R.; Wang, Q.-Y.; Zheng, P.-C.; Zhang, F.-X.; Zhang, J.-P. MicroRNA-128 Enhances Neuroprotective Effects of Dexmedetomidine on Neonatal Mice with Hypoxic-Ischemic Brain Damage by Targeting WNT1. Biomed. Pharmacother. 2019, 113, 108671. [Google Scholar] [CrossRef]
- Bao, N.; Tang, B. Organ-Protective Effects and the Underlying Mechanism of Dexmedetomidine. Mediators Inflamm. 2020, 2020, 6136105. [Google Scholar] [CrossRef]
- Tanabe, K.; Matsushima-Nishiwaki, R.; Kozawa, O.; Iida, H. Dexmedetomidine Suppresses Interleukin-1β-Induced Interleukin-6 Synthesis in Rat Glial Cells. Int. J. Mol. Med. 2014, 34, 1032–1038. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, J.; Qian, W.; Zhao, J.; Sun, L.; Qian, Y.; Xiao, H. Dexmedetomidine Inhibits Tumor Necrosis Factor-Alpha and Interleukin 6 in Lipopolysaccharide-Stimulated Astrocytes by Suppression of c-Jun N-Terminal Kinases. Inflammation 2014, 37, 942–949. [Google Scholar] [CrossRef]
- Zhu, Y.-J.; Peng, K.; Meng, X.-W.; Ji, F.-H. Attenuation of Neuroinflammation by Dexmedetomidine Is Associated with Activation of a Cholinergic Anti-Inflammatory Pathway in a Rat Tibial Fracture Model. Brain Res. 2016, 1644, 1–8. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.; Zhang, L.; Wang, G.; Zhang, M.; Yu, Y. Neuroprotection of Dexmedetomidine against Cerebral Ischemia-Reperfusion Injury in Rats: Involved in Inhibition of NF-ΚB and Inflammation Response. Biomol. Ther. 2017, 25, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Chiu, K.-M.; Lin, T.-Y.; Lu, C.-W.; Wang, S.-J. Inhibitory Effect of Glutamate Release from Rat Cerebrocortical Nerve Terminals by A2 Adrenoceptor Agonist Dexmedetomidine. Eur. J. Pharmacol. 2011, 670, 137–147. [Google Scholar] [CrossRef]
- Wang, X.; Shan, Y.; Tang, Z.; Gao, L.; Liu, H. Neuroprotective Effects of Dexmedetomidine against Isoflurane-Induced Neuronal Injury via Glutamate Regulation in Neonatal Rats. Drug Des. Devel. Ther. 2018, 13, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Ling, X.; Song, R.; Gao, X.; Liang, Z.; Fang, F.; Cang, J. Upregulation of GLT-1 via PI3K/Akt Pathway Contributes to Neuroprotection Induced by Dexmedetomidine. Front. Neurol. 2019, 10, 1041. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-H.; Kim, D.-O.; Yi, J.-W.; Park, S.-W.; Kang, W.-J.; Choi, Y.-K.; Kim, S.-H.; Ko, I.-G.; Jin, J.-J.; Kim, S.-E.; et al. Dexmedetomidine, α2 -Adrenoceptor Agonist, Does Not Induce Apoptosis in the Brachial Plexus of Rats. Anim. Cells Syst. 2014, 18, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Ding, T.; Yu, L.; Zhong, Y.; Dai, H.; Yan, M. Dexmedetomidine Protects against Oxygen–Glucose Deprivation-Induced Injury through the I2 Imidazoline Receptor-PI3K/AKT Pathway in Rat C6 Glioma Cells. J. Pharm. Pharmacol. 2011, 64, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, L.; Gao, M.; Guo, W.; Ma, Y. Dexmedetomidine Reduces Postoperative Delirium after Joint Replacement in Elderly Patients with Mild Cognitive Impairment. Aging Clin. Exp. Res. 2016, 28, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Meng, Z.-T.; Wu, X.-H.; Cui, F.; Li, H.-L.; Wang, D.-X.; Zhu, X.; Zhu, S.-N.; Maze, M.; Ma, D. Dexmedetomidine for Prevention of Delirium in Elderly Patients after Non-Cardiac Surgery: A Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2016, 388, 1893–1902. [Google Scholar] [CrossRef] [Green Version]
- Pandharipande, P.P.; Sanders, R.D.; Girard, T.D.; McGrane, S.; Thompson, J.L.; Shintani, A.K.; Herr, D.L.; Maze, M.; Ely, E.W. MENDS investigators Effect of Dexmedetomidine versus Lorazepam on Outcome in Patients with Sepsis: An a Priori-Designed Analysis of the MENDS Randomized Controlled Trial. Crit. Care 2010, 14, R38. [Google Scholar] [CrossRef] [Green Version]
- Finnerty, M.; Marczynski, T.J.; Amirault, H.J.; Urbancic, M.; Andersen, B.R. Benzodiazepines Inhibit Neutrophil Chemotaxis and Superoxide Production in a Stimulus Dependent Manner; PK-11195 Antagonizes These Effects. Immunopharmacology 1991, 22, 185–193. [Google Scholar] [CrossRef]
- Miles, B.A.; Lafuse, W.P.; Zwilling, B.S. Binding of Alpha-Adrenergic Receptors Stimulates the Anti-Mycobacterial Activity of Murine Peritoneal Macrophages. J. Neuroimmunol. 1996, 71, 19–24. [Google Scholar] [CrossRef]
- Shokri, H.; Ali, I. A Randomized Control Trial Comparing Prophylactic Dexmedetomidine versus Clonidine on Rates and Duration of Delirium in Older Adult Patients Undergoing Coronary Artery Bypass Grafting. J. Clin. Anesth. 2020, 61, 109622. [Google Scholar] [CrossRef]
- Chen, P.-H.; Tsuang, F.-Y.; Lee, C.-T.; Yeh, Y.-C.; Cheng, H.-L.; Lee, T.-S.; Chang, Y.-W.; Cheng, Y.-J.; Wu, C.-Y. Neuroprotective Effects of Intraoperative Dexmedetomidine versus Saline Infusion Combined with Goal-Directed Haemodynamic Therapy for Patients Undergoing Cranial Surgery: A Randomised Controlled Trial. Eur. J. Anaesthesiol. 2021, 38, 1262–1271. [Google Scholar] [CrossRef]
- Ge, Y.; Li, Q.; Nie, Y.; Gao, J.; Luo, K.; Fang, X.; Wang, C. Dexmedetomidine Improves Cognition after Carotid Endarterectomy by Inhibiting Cerebral Inflammation and Enhancing Brain-Derived Neurotrophic Factor Expression. J. Int. Med. Res. 2019, 47, 2471–2482. [Google Scholar] [CrossRef]
- Luo, X.; Zheng, X.; Huang, H. Protective Effects of Dexmedetomidine on Brain Function of Glioma Patients Undergoing Craniotomy Resection and Its Underlying Mechanism. Clin. Neurol. Neurosurg. 2016, 146, 105–108. [Google Scholar] [CrossRef]
- Calin, G.A.; Liu, C.-G.; Sevignani, C.; Ferracin, M.; Felli, N.; Dumitru, C.D.; Shimizu, M.; Cimmino, A.; Zupo, S.; Dono, M.; et al. MicroRNA Profiling Reveals Distinct Signatures in B Cell Chronic Lymphocytic Leukemias. Proc. Natl. Acad. Sci. USA 2004, 101, 11755–11760. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of MicroRNAs in Serum: A Novel Class of Biomarkers for Diagnosis of Cancer and Other Diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Huang, W. MicroRNAs: Biomarkers, Diagnostics, and Therapeutics. Methods Mol. Biol. 2017, 1617, 57–67. [Google Scholar] [CrossRef]
- Basak, I.; Patil, K.S.; Alves, G.; Larsen, J.P.; Møller, S.G. MicroRNAs as Neuroregulators, Biomarkers and Therapeutic Agents in Neurodegenerative Diseases. Cell. Mol. Life Sci. CMLS 2016, 73, 811–827. [Google Scholar] [CrossRef]
- Alawneh, K.Z.; Raffee, L.A.; Alshehabat, M.A.M.; Haddad, H.; Jaradat, S.A. Characterizing and Profiling MicroRNAs in Dogs Undergoing Induced Ischemic Brain Stroke After Middle Cerebral Artery Occlusion Under Fluoroscopic Guidance. Vasc. Health Risk Manag. 2021, 17, 543–550. [Google Scholar] [CrossRef]
- Liu, D.-Z.; Tian, Y.; Ander, B.P.; Xu, H.; Stamova, B.S.; Zhan, X.; Turner, R.J.; Jickling, G.; Sharp, F.R. Brain and Blood MicroRNA Expression Profiling of Ischemic Stroke, Intracerebral Hemorrhage, and Kainate Seizures. J. Cereb. Blood Flow Metab. 2010, 30, 92–101. [Google Scholar] [CrossRef]
- Zeng, L.; Liu, J.; Wang, Y.; Wang, L.; Weng, S.; Tang, Y.; Zheng, C.; Cheng, Q.; Chen, S.; Yang, G.-Y. MicroRNA-210 as a Novel Blood Biomarker in Acute Cerebral Ischemia. Front. Biosci. Elite Ed. 2011, 3, 1265–1272. [Google Scholar] [CrossRef]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating MiR-125a-5p, MiR-125b-5p, and MiR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef]
- Ponnusamy, V.; Yip, P.K. The Role of MicroRNAs in Newborn Brain Development and Hypoxic Ischaemic Encephalopathy. Neuropharmacology 2019, 149, 55–65. [Google Scholar] [CrossRef]
- Ma, F.; Zhang, X.; Yin, K.-J. MicroRNAs in Central Nervous System Diseases: A Prospective Role in Regulating Blood-Brain Barrier Integrity. Exp. Neurol. 2020, 323, 113094. [Google Scholar] [CrossRef]
- Saugstad, J.A. MicroRNAs as Effectors of Brain Function with Roles in Ischemia and Injury, Neuroprotection, and Neurodegeneration. J. Cereb. Blood Flow Metab. 2010, 30, 1564–1576. [Google Scholar] [CrossRef]
- Tan, K.S.; Armugam, A.; Sepramaniam, S.; Lim, K.Y.; Setyowati, K.D.; Wang, C.W.; Jeyaseelan, K. Expression Profile of MicroRNAs in Young Stroke Patients. PLoS ONE 2009, 4, e7689. [Google Scholar] [CrossRef] [Green Version]
- Jeyaseelan, K.; Lim, K.Y.; Armugam, A. MicroRNA Expression in the Blood and Brain of Rats Subjected to Transient Focal Ischemia by Middle Cerebral Artery Occlusion. Stroke 2008, 39, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voelz, C.; Ebrahimy, N.; Zhao, W.; Habib, P.; Zendedel, A.; Pufe, T.; Beyer, C.; Slowik, A. Transient Focal Cerebral Ischemia Leads to MiRNA Alterations in Different Brain Regions, Blood Serum, Liver, and Spleen. Int. J. Mol. Sci. 2021, 23, 161. [Google Scholar] [CrossRef] [PubMed]
- Bertero, T.; Rezzonico, R.; Pottier, N.; Mari, B. Impact of MicroRNAs in the Cellular Response to Hypoxia. Int. Rev. Cell Mol. Biol. 2017, 333, 91–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, J.; Duan, F.; Cong, L.; Qi, X. The Role of the MicroRNA Regulatory Network in Alzheimer’s Disease: A Bioinformatics Analysis. Arch. Med. Sci. 2021, 18, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Kashif, H.; Shah, D.; Sukumari-Ramesh, S. Dysregulation of MicroRNA and Intracerebral Hemorrhage: Roles in Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 8115. [Google Scholar] [CrossRef]
- Wang, H.-W.; Noland, C.; Siridechadilok, B.; Taylor, D.W.; Ma, E.; Felderer, K.; Doudna, J.A.; Nogales, E. Structural Insights into RNA Processing by the Human RISC-Loading Complex. Nat. Struct. Mol. Biol. 2009, 16, 1148–1153. [Google Scholar] [CrossRef] [Green Version]
- Doxakis, E. Post-Transcriptional Regulation of α-Synuclein Expression by Mir-7 and Mir-153. J. Biol. Chem. 2010, 285, 12726–12734. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Liu, G.; Jin, S.M.; Parisiadou, L.; Xie, C.; Yu, J.; Sun, L.; Ma, B.; Ding, J.; Vancraenenbroeck, R.; et al. MicroRNA-205 Regulates the Expression of Parkinson’s Disease-Related Leucine-Rich Repeat Kinase 2 Protein. Hum. Mol. Genet. 2013, 22, 608–620. [Google Scholar] [CrossRef] [Green Version]
- Abbasi-Habashi, S.; Jickling, G.C.; Winship, I.R. Immune Modulation as a Key Mechanism for the Protective Effects of Remote Ischemic Conditioning After Stroke. Front. Neurol. 2021, 12, 746486. [Google Scholar] [CrossRef]
- Vinciguerra, A.; Formisano, L.; Cerullo, P.; Guida, N.; Cuomo, O.; Esposito, A.; Di Renzo, G.; Annunziato, L.; Pignataro, G. MicroRNA-103-1 Selectively Downregulates Brain NCX1 and Its Inhibition by Anti-MiRNA Ameliorates Stroke Damage and Neurological Deficits. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 1829–1838. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.-J.; Ouyang, Y.-B.; Xiong, X.; Stary, C.M.; Giffard, R.G. Post-Stroke Treatment with MiR-181 Antagomir Reduces Injury and Improves Long-Term Behavioral Recovery in Mice after Focal Cerebral Ischemia. Exp. Neurol. 2015, 264, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Y.; Chen, W.; Li, W.; Huang, Y.; Duan, D.; Ge, L.; He, J.; Liu, J.; Hu, Z.; Lu, M. Ischemic-Hypoxic Preconditioning Enhances the Mitochondrial Function Recovery of Transplanted Olfactory Mucosa Mesenchymal Stem Cells via MiR-181a Signaling in Ischemic Stroke. Aging 2021, 13, 11234–11256. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The Nuclear RNase III Drosha Initiates MicroRNA Processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Lytle, J.R.; Yario, T.A.; Steitz, J.A. Target MRNAs Are Repressed as Efficiently by MicroRNA-Binding Sites in the 5’ UTR as in the 3’ UTR. Proc. Natl. Acad. Sci. USA 2007, 104, 9667–9672. [Google Scholar] [CrossRef] [Green Version]
- Kiriakidou, M.; Tan, G.S.; Lamprinaki, S.; De Planell-Saguer, M.; Nelson, P.T.; Mourelatos, Z. An MRNA M7G Cap Binding-like Motif within Human Ago2 Represses Translation. Cell 2007, 129, 1141–1151. [Google Scholar] [CrossRef] [Green Version]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of MicroRNA Biogenesis and Its Crosstalk with Other Cellular Pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Forman, J.J.; Legesse-Miller, A.; Coller, H.A. A Search for Conserved Sequences in Coding Regions Reveals That the Let-7 MicroRNA Targets Dicer within Its Coding Sequence. Proc. Natl. Acad. Sci. USA 2008, 105, 14879–14884. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, W.; Liu, Y.; Liu, T.; Li, C.; Wang, L. Oncogenic Role of MicroRNA-532-5p in Human Colorectal Cancer via Targeting of the 5’UTR of RUNX3. Oncol. Lett. 2018, 15, 7215–7220. [Google Scholar] [CrossRef] [Green Version]
- Nallamshetty, S.; Chan, S.Y.; Loscalzo, J. Hypoxia: A Master Regulator of MicroRNA Biogenesis and Activity. Free Radic. Biol. Med. 2013, 64, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Minhas, G.; Mathur, D.; Ragavendrasamy, B.; Sharma, N.K.; Paanu, V.; Anand, A. Hypoxia in CNS Pathologies: Emerging Role of MiRNA-Based Neurotherapeutics and Yoga Based Alternative Therapies. Front. Neurosci. 2017, 11, 386. [Google Scholar] [CrossRef]
- Morimoto, K.; Eguchi, R.; Kitano, T.; Otsuguro, K.-I. Alpha and Beta Adrenoceptors Activate Interleukin-6 Transcription through Different Pathways in Cultured Astrocytes from Rat Spinal Cord. Cytokine 2021, 142, 155497. [Google Scholar] [CrossRef]
- Morimoto, K.; Kitano, T.; Eguchi, R.; Otsuguro, K.-I. Bidirectional Modulation of TNF-α Transcription via α- and β-Adrenoceptors in Cultured Astrocytes from Rat Spinal Cord. Biochem. Biophys. Res. Commun. 2020, 528, 78–84. [Google Scholar] [CrossRef]
- Nohata, N.; Goto, Y.; Gutkind, J.S. Onco-GPCR Signaling and Dysregulated Expression of MicroRNAs in Human Cancer. J. Hum. Genet. 2017, 62, 87–96. [Google Scholar] [CrossRef]
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic Gper Signaling Regulates Mir144 Expression in Cancer Cells and Cancer-Associated Fibroblasts (Cafs). Oncotarget 2015, 6, 16573–16587. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; He, H.; Chen, Q. Estradiol Induces HOTAIR Levels via GPER-Mediated MiR-148a Inhibition in Breast Cancer. J. Transl. Med. 2015, 13, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Tang, H.; Li, H.; Zhao, R.; Huang, Q.; Liu, J. Recent Advances in the Development of Neuroprotective Agents and Therapeutic Targets in the Treatment of Cerebral Ischemia. Eur. J. Med. Chem. 2019, 162, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, Y.; Li, F.; Liu, X.; Zhang, H.; Zhu, Z.; Huang, H.; Xu, X. Circ-Camk4 Involved in Cerebral Ischemia/Reperfusion Induced Neuronal Injury. Sci. Rep. 2020, 10, 7012. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, H.; Shang, H.; Chen, X.; Yang, S.; Qu, Y.; Ding, J.; Li, X. Circular RNA Circ_0000950 Promotes Neuron Apoptosis, Suppresses Neurite Outgrowth and Elevates Inflammatory Cytokines Levels via Directly Sponging MiR-103 in Alzheimer’s Disease. Cell Cycle Georget. Tex 2019, 18, 2197–2214. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, L.; Wang, S.; Hong, Q. Circular RNA CircDLGAP4 Exerts Neuroprotective Effects via Modulating MiR-134-5p/CREB Pathway in Parkinson’s Disease. Biochem. Biophys. Res. Commun. 2020, 522, 388–394. [Google Scholar] [CrossRef]
- Chen, H.; Gu, B.; Zhao, X.; Zhao, Y.; Huo, S.; Liu, X.; Lu, H. Circular RNA Hsa_circ_0007364 Increases Cervical Cancer Progression through Activating Methionine Adenosyltransferase II Alpha (MAT2A) Expression by Restraining MicroRNA-101-5p. Bioengineered 2020, 11, 1269–1279. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y. Circular RNA Cerebellar Degeneration-Related Protein 1 Antisense RNA (Circ-CDR1as) Downregulation Induced by Dexmedetomidine Treatment Protects Hippocampal Neurons against Hypoxia/Reoxygenation Injury through the MicroRNA-28-3p (MiR-28-3p)/Tumor Necrosis Factor Receptor-Associated Factor-3 (TRAF3) Axis. Bioengineered 2021, 12, 10512–10524. [Google Scholar] [CrossRef]
- Liu, W.; Shao, C.; Zang, C.; Sun, J.; Xu, M.; Wang, Y. Protective Effects of Dexmedetomidine on Cerebral Ischemia/Reperfusion Injury via the MicroRNA-214/ROCK1/NF-ΚB Axis. BMC Anesthesiol. 2021, 21, 203. [Google Scholar] [CrossRef]
- Sifringer, M.; von Haefen, C.; Krain, M.; Paeschke, N.; Bendix, I.; Bührer, C.; Spies, C.D.; Endesfelder, S. Neuroprotective Effect of Dexmedetomidine on Hyperoxia-Induced Toxicity in the Neonatal Rat Brain. Oxid. Med. Cell. Longev. 2015, 2015, 530371. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-J.; Zhang, D.-Z.; Xi, Y.; Wu, C.-A. Protective Effect of Dexmedetomidine on Neuronal Hypoxic Injury through Inhibition of MiR-134. Hum. Exp. Toxicol. 2021, 40, 2145–2155. [Google Scholar] [CrossRef]
- Chen, Y.; Fan, Z.; Wu, Q. Dexmedetomidine Improves Oxygen-Glucose Deprivation/Reoxygenation (OGD/R)-Induced Neurological Injury through Regulating SNHG11/MiR-324-3p/VEGFA Axis. Bioengineered 2021, 12, 4794–4804. [Google Scholar] [CrossRef]
- Shi, X.; Yan, C.; Liu, B.; Yang, C.; Nie, X.; Wang, X.; Zheng, J.; Wang, Y.; Zhu, Y. MiR-381 Regulates Neural Stem Cell Proliferation and Differentiation via Regulating Hes1 Expression. PLoS ONE 2015, 10, e0138973. [Google Scholar] [CrossRef]
- Mamun, A.A.; Chauhan, A.; Qi, S.; Ngwa, C.; Xu, Y.; Sharmeen, R.; Hazen, A.L.; Li, J.; Aronowski, J.A.; McCullough, L.D.; et al. Microglial IRF5-IRF4 Regulatory Axis Regulates Neuroinflammation after Cerebral Ischemia and Impacts Stroke Outcomes. Proc. Natl. Acad. Sci. USA 2020, 117, 1742–1752. [Google Scholar] [CrossRef]
- Degos, V.; Charpentier, T.L.; Chhor, V.; Brissaud, O.; Lebon, S.; Schwendimann, L.; Bednareck, N.; Passemard, S.; Mantz, J.; Gressens, P. Neuroprotective Effects of Dexmedetomidine against Glutamate Agonist-Induced Neuronal Cell Death Are Related to Increased Astrocyte Brain-Derived Neurotrophic Factor Expression. Anesthesiology 2013, 118, 1123–1132. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Wang, L.; Liu, W.; Zhang, Y.; Hu, Z.; Guo, H.; Lv, J.; Du, H. Dexmedetomidine Had Neuroprotective Effects on Hippocampal Neuronal Cells via Targeting LncRNA SHNG16 Mediated MicroRNA-10b-5p/BDNF Axis. Mol. Cell. Biochem. 2020, 469, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Yang, F.; Tang, Z.; Bi, C.; Sun, S.; Zhang, Y.; Liu, H. Dexmedetomidine Ameliorates the Neurotoxicity of Sevoflurane on the Immature Brain Through the BMP/SMAD Signaling Pathway. Front. Neurosci. 2018, 12, 964. [Google Scholar] [CrossRef]
- Fang, H.; Li, H.-F.; He, M.-H.; Yan, J.-Y.; Yang, M.; Zhang, F.-X.; Wang, R.-R.; Wang, Q.-Y.; Zhang, J.-P. Long Non-Coding RNA MALAT1 Sponges MicroRNA-429 to Regulate Apoptosis of Hippocampal Neurons in Hypoxic-Ischemic Brain Damage by Regulating WNT1. Brain Res. Bull. 2019, 152, 1–10. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, G.; Zeng, Q.; Gao, R.; Zhang, S.; Wang, L.; Liu, B.; Yu, Y.; Zhao, A.; Li, R.; et al. MiR-29b Expression Is Associated with a Dexmedetomidine-Mediated Protective Effect against Oxygen-Glucose Deprivation-Induced Injury to SK-N-SH Cells in Vitro. Cell Biol. Int. 2018, 42, 344–352. [Google Scholar] [CrossRef]
- Wu, L.; Xi, Y.; Kong, Q. Dexmedetomidine Protects PC12 Cells from Oxidative Damage through Regulation of MiR-199a/HIF-1α. Artif. Cells Nanomed. Biotechnol. 2020, 48, 506–514. [Google Scholar] [CrossRef]
- Dahmani, S.; Rouelle, D.; Gressens, P.; Mantz, J. Effects of Dexmedetomidine on Hippocampal Focal Adhesion Kinase Tyrosine Phosphorylation in Physiologic and Ischemic Conditions. Anesthesiology 2005, 103, 969–977. [Google Scholar] [CrossRef]
- Reyland, M.E.; Barzen, K.A.; Anderson, S.M.; Quissell, D.O.; Matassa, A.A. Activation of PKC Is Sufficient to Induce an Apoptotic Program in Salivary Gland Acinar Cells. Cell Death Differ. 2000, 7, 1200–1209. [Google Scholar] [CrossRef]
- Zhu, Y.-S.; Liu, Z.; Min, J.; Xiong, Y.-F. Dexmedetomidine Reduces the Apoptosis of Rat Hippocampal Neurons via Mediating ERK1/2 Signal Pathway by Targeting MiR-155. Acta Histochem. 2021, 123, 151734. [Google Scholar] [CrossRef]
- Mielcarek, M.; Zielonka, D.; Carnemolla, A.; Marcinkowski, J.T.; Guidez, F. HDAC4 as a Potential Therapeutic Target in Neurodegenerative Diseases: A Summary of Recent Achievements. Front. Cell. Neurosci. 2015, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.-M.; Liu, J.; Wang, Y.; Zhang, M.-H. Silencing of Long Noncoding RNA MEG3 Enhances Cerebral Protection of Dexmedetomidine against Hypoxic-Ischemic Brain Damage in Neonatal Mice by Binding to MiR-129-5p. J. Cell. Biochem. 2019, 120, 7978–7988. [Google Scholar] [CrossRef]
- Rayasam, A.; Hsu, M.; Kijak, J.A.; Kissel, L.; Hernandez, G.; Sandor, M.; Fabry, Z. Immune Responses in Stroke: How the Immune System Contributes to Damage and Healing after Stroke and How This Knowledge Could Be Translated to Better Cures? Immunology 2018, 154, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Li, P.; Guo, Y.; Wang, H.; Leak, R.K.; Chen, S.; Gao, Y.; Chen, J. Microglia/Macrophage Polarization Dynamics Reveal Novel Mechanism of Injury Expansion after Focal Cerebral Ischemia. Stroke 2012, 43, 3063–3070. [Google Scholar] [CrossRef] [Green Version]
- Ta Na, H.S.; Nuo, M.; Meng, Q.-T.; Xia, Z.-Y. The Pathway of Let-7a-1/2-3p and HMGB1 Mediated Dexmedetomidine Inhibiting Microglia Activation in Spinal Cord Ischemia-Reperfusion Injury Mice. J. Mol. Neurosci. 2019, 69, 106–114. [Google Scholar] [CrossRef]
- Perez-Zoghbi, J.F.; Zhu, W.; Grafe, M.R.; Brambrink, A.M. Dexmedetomidine-Mediated Neuroprotection against Sevoflurane-Induced Neurotoxicity Extends to Several Brain Regions in Neonatal Rats. Br. J. Anaesth. 2017, 119, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Sanders, R.D.; Sun, P.; Patel, S.; Li, M.; Maze, M.; Ma, D. Dexmedetomidine Provides Cortical Neuroprotection: Impact on Anaesthetic-Induced Neuroapoptosis in the Rat Developing Brain. Acta Anaesthesiol. Scand. 2010, 54, 710–716. [Google Scholar] [CrossRef]
- Duan, X.; Li, Y.; Zhou, C.; Huang, L.; Dong, Z. Dexmedetomidine Provides Neuroprotection: Impact on Ketamine-Induced Neuroapoptosis in the Developing Rat Brain. Acta Anaesthesiol. Scand. 2014, 58, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Twaroski, D.; Bosnjak, Z.J.; Bai, X. MicroRNAs: New Players in Anesthetic-Induced Developmental Neurotoxicity. Pharm. Anal. Acta 2015, 6, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, B.; Lim, C.S.; Kim, Y.-H.; Lee, J.U.; Kim, Y.M.; Jung, C.; Jo, Y. Comparison of Topical Ropivacaine with and without Ketamine on Post-Surgical Pain in Children Undergoing Tonsillectomy: A Randomized Controlled Double-Blind Study. J. Anesth. 2017, 31, 559–564. [Google Scholar] [CrossRef] [PubMed]
- David, K.K.; Andrabi, S.A.; Dawson, T.M.; Dawson, V.L. Parthanatos, a Messenger of Death. Front. Biosci. Landmark Ed. 2009, 14, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.-H.; Yang, Y.-L.; Cheng, X.; Liu, M.; Zhang, S.-S.; Wang, Y.-H.; Du, G.-H. Baicalein Attenuates Caspase-Independent Cells Death via Inhibiting PARP-1 Activation and AIF Nuclear Translocation in Cerebral Ischemia/Reperfusion Rats. Apoptosis Int. J. Program. Cell Death 2020, 25, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Lu, B.; Feng, C.; Wang, C.; Wang, Y.; Luo, T.; Feng, J.; Jia, H.; Chi, G.; Luo, Y.; et al. Deoxypodophyllotoxin Triggers Parthanatos in Glioma Cells via Induction of Excessive ROS. Cancer Lett. 2016, 371, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Gao, R.; Chen, L. Dexmedetomidine Regulates Sevoflurane-Induced Neurotoxicity through the MiR-330-3p/ULK1 Axis. J. Biochem. Mol. Toxicol. 2021, 35, e22919. [Google Scholar] [CrossRef] [PubMed]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.R.; Ver Donck, L.; et al. Systemic Immune Activation Leads to Neuroinflammation and Sickness Behavior in Mice. Mediators Inflamm. 2013, 2013, 271359. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, Y.K.; Yuk, D.Y.; Choi, D.Y.; Ban, S.B.; Oh, K.W.; Hong, J.T. Neuro-Inflammation Induced by Lipopolysaccharide Causes Cognitive Impairment through Enhancement of Beta-Amyloid Generation. J. Neuroinflamm. 2008, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Ramirez, M.A.; Wu, D.; Pryce, G.; Simpson, J.E.; Reijerkerk, A.; King-Robson, J.; Kay, O.; de Vries, H.E.; Hirst, M.C.; Sharrack, B.; et al. MicroRNA-155 Negatively Affects Blood-Brain Barrier Function during Neuroinflammation. FASEB J. 2014, 28, 2551–2565. [Google Scholar] [CrossRef]
- Ning, Q.; Liu, Z.; Wang, X.; Zhang, R.; Zhang, J.; Yang, M.; Sun, H.; Han, F.; Zhao, W.; Zhang, X. Neurodegenerative Changes and Neuroapoptosis Induced by Systemic Lipopolysaccharide Administration Are Reversed by Dexmedetomidine Treatment in Mice. Neurol. Res. 2017, 39, 357–366. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Wyss-Coray, T.; Mucke, L. Inflammation in Neurodegenerative Disease—A Double-Edged Sword. Neuron 2002, 35, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Hu, S.; Li, Y.; Xue, D.; Wu, X. Dexmedetomidine, an Alpha 2a Adrenergic Receptor Agonist, Mitigates Experimental Autoimmune Encephalomyelitis by Desensitization of CXCR7 in Microglia. Biochemistry 2018, 57, 4197–4205. [Google Scholar] [CrossRef]
- Freeman, L.C.; Ting, J.P.-Y. The Pathogenic Role of the Inflammasome in Neurodegenerative Diseases. J. Neurochem. 2016, 136 (Suppl. 1), 29–38. [Google Scholar] [CrossRef]
- Schweneker, K.; Gorka, O.; Schweneker, M.; Poeck, H.; Tschopp, J.; Peschel, C.; Ruland, J.; Gross, O. The Mycobacterial Cord Factor Adjuvant Analogue Trehalose-6,6′-Dibehenate (TDB) Activates the Nlrp3 Inflammasome. Immunobiology 2013, 218, 664–673. [Google Scholar] [CrossRef]
- Meihe, L.; Shan, G.; Minchao, K.; Xiaoling, W.; Peng, A.; Xili, W.; Jin, Z.; Huimin, D. The Ferroptosis-NLRP1 Inflammasome: The Vicious Cycle of an Adverse Pregnancy. Front. Cell Dev. Biol. 2021, 9, 2137. [Google Scholar] [CrossRef]
- Kaushal, V.; Dye, R.; Pakavathkumar, P.; Foveau, B.; Flores, J.; Hyman, B.; Ghetti, B.; Koller, B.H.; LeBlanc, A.C. Neuronal NLRP1 Inflammasome Activation of Caspase-1 Coordinately Regulates Inflammatory Interleukin-1-Beta Production and Axonal Degeneration-Associated Caspase-6 Activation. Cell Death Differ. 2015, 22, 1676–1686. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.G.; Dash, P.; Aldridge, J.R.; Ellebedy, A.H.; Reynolds, C.; Funk, A.J.; Martin, W.J.; Lamkanfi, M.; Webby, R.J.; Boyd, K.L.; et al. The Intracellular Sensor NLRP3 Mediates Key Innate and Healing Responses to Influenza A Virus via the Regulation of Caspase-1. Immunity 2009, 30, 566–575. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.-R.; Chen, Y.; Chen, Y.-N.; Qiu, G.-L.; Wen, J.-G.; Zheng, Y.; Li, X.-F.; Cheng, H.; Li, Y.-H.; Li, J. Dexmedetomidine Inhibits the Invasion, Migration, and Inflammation of Rheumatoid Arthritis Fibroblast-like Synoviocytes by Reducing the Expression of NLRC5. Int. Immunopharmacol. 2020, 82, 106374. [Google Scholar] [CrossRef]
- Petralia, M.C.; Battaglia, G.; Bruno, V.; Pennisi, M.; Mangano, K.; Lombardo, S.D.; Fagone, P.; Cavalli, E.; Saraceno, A.; Nicoletti, F.; et al. The Role of Macrophage Migration Inhibitory Factor in Alzheimer’s Disease: Conventionally Pathogenetic or Unconventionally Protective? Molcules 2020, 25, 291. [Google Scholar] [CrossRef] [Green Version]
- Tilstam, P.V.; Qi, D.; Leng, L.; Young, L.; Bucala, R. MIF Family Cytokines in Cardiovascular Diseases and Prospects for Precision-Based Therapeutics. Expert Opin. Ther. Targets 2017, 21, 671–683. [Google Scholar] [CrossRef]
- Stosic-Grujicic, S.; Stojanovic, I.; Nicoletti, F. MIF in Autoimmunity and Novel Therapeutic Approaches. Autoimmun. Rev. 2009, 8, 244–249. [Google Scholar] [CrossRef]
- Günther, S.; Fagone, P.; Jalce, G.; Atanasov, A.G.; Guignabert, C.; Nicoletti, F. Role of MIF and D-DT in Immune-Inflammatory, Autoimmune, and Chronic Respiratory Diseases: From Pathogenic Factors to Therapeutic Targets. Drug Discov. Today 2019, 24, 428–439. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Wong, D.W.L.; Bucala, R.; Djudjaj, S.; Boor, P. Evolving Complexity of MIF Signaling. Cell. Signal. 2019, 57, 76–88. [Google Scholar] [CrossRef]
- Li, S.; Nie, K.; Zhang, Q.; Guo, M.; Qiu, Y.; Li, Y.; Gao, Y.; Wang, L. Macrophage Migration Inhibitory Factor Mediates Neuroprotective Effects by Regulating Inflammation, Apoptosis and Autophagy in Parkinson’s Disease. Neuroscience 2019, 416, 50–62. [Google Scholar] [CrossRef]
- Benedek, G.; Meza-Romero, R.; Jordan, K.; Zhang, Y.; Nguyen, H.; Kent, G.; Li, J.; Siu, E.; Frazer, J.; Piecychna, M.; et al. MIF and D-DT Are Potential Disease Severity Modifiers in Male MS Subjects. Proc. Natl. Acad. Sci. USA 2017, 114, E8421–E8429. [Google Scholar] [CrossRef] [Green Version]
- Macrophage Migration Inhibitory Factor Potentiates Autoimmune-Mediated Neuroinflammation—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/23797673/ (accessed on 29 March 2022).
- Petralia, M.C.; Mazzon, E.; Fagone, P.; Basile, M.S.; Lenzo, V.; Quattropani, M.C.; Bendtzen, K.; Nicoletti, F. Pathogenic Contribution of the Macrophage Migration Inhibitory Factor Family to Major Depressive Disorder and Emerging Tailored Therapeutic Approaches. J. Affect. Disord. 2020, 263, 15–24. [Google Scholar] [CrossRef]
- Lee, S.-T.; Chu, K.; Im, W.-S.; Yoon, H.-J.; Im, J.-Y.; Park, J.-E.; Park, K.-H.; Jung, K.-H.; Lee, S.K.; Kim, M.; et al. Altered MicroRNA Regulation in Huntington’s Disease Models. Exp. Neurol. 2011, 227, 172–179. [Google Scholar] [CrossRef]
- Figueroa-Romero, C.; Hur, J.; Lunn, J.S.; Paez-Colasante, X.; Bender, D.E.; Yung, R.; Sakowski, S.A.; Feldman, E.L. Expression of MicroRNAs in Human Post-Mortem Amyotrophic Lateral Sclerosis Spinal Cords Provides Insight into Disease Mechanisms. Mol. Cell. Neurosci. 2016, 71, 34–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putteeraj, M.; Fairuz, Y.M.; Teoh, S.L. MicroRNA Dysregulation in Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Li, T.; Zhang, W.; Ren, J.; Li, Z.; Wu, G. MiR-199a-3p Inhibits Cell Proliferation and Induces Apoptosis by Targeting YAP1, Suppressing Jagged1-Notch Signaling in Human Hepatocellular Carcinoma. J. Biomed. Sci. 2016, 23, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Zhang, D.-F.; Luo, R.; Wu, Y.; Zhou, H.; Kong, L.-L.; Bi, R.; Yao, Y.-G. A Systematic Integrated Analysis of Brain Expression Profiles Reveals YAP1 and Other Prioritized Hub Genes as Important Upstream Regulators in Alzheimer’s Disease. Alzheimers Dement. 2018, 14, 215–229. [Google Scholar] [CrossRef]
- Guo, H.; Tian, J.; Zhu, J.; Li, L.; Sun, K.; Shao, S.; Cui, G. Electroacupuncture Suppressed Neuronal Apoptosis and Improved Cognitive Impairment in the AD Model Rats Possibly via Downregulation of Notch Signaling Pathway. Evid. Based Complement. Alternat. Med. 2015, 2015, e393569. [Google Scholar] [CrossRef]
- Sun, W.; Zhao, J.; Li, C. Dexmedetomidine Provides Protection Against Hippocampal Neuron Apoptosis and Cognitive Impairment in Mice with Alzheimer’s Disease by Mediating the MiR-129/YAP1/JAG1 Axis. Mol. Neurobiol. 2020, 57, 5044–5055. [Google Scholar] [CrossRef]
- Latini, A.; Borgiani, P.; Novelli, G.; Ciccacci, C. MiRNAs in Drug Response Variability: Potential Utility as Biomarkers for Personalized Medicine. Pharmacogenomics 2019, 20, 1049–1059. [Google Scholar] [CrossRef]
- Thummel, K.E.; Lin, Y.S. Sources of Interindividual Variability. Methods Mol. Biol. 2014, 1113, 363–415. [Google Scholar] [CrossRef]
- Turner, R.M.; Park, B.K.; Pirmohamed, M. Parsing Interindividual Drug Variability: An Emerging Role for Systems Pharmacology. Wiley Interdiscip. Rev. Syst. Biol. Med. 2015, 7, 221–241. [Google Scholar] [CrossRef]
- Baer-Dubowska, W.; Majchrzak-Celińska, A.; Cichocki, M. Pharmocoepigenetics: A New Approach to Predicting Individual Drug Responses and Targeting New Drugs. Pharmacol. Rep. 2011, 63, 293–304. [Google Scholar] [CrossRef]
- Li, M.-P.; Hu, Y.-D.; Hu, X.-L.; Zhang, Y.-J.; Yang, Y.-L.; Jiang, C.; Tang, J.; Chen, X.-P. MiRNAs and MiRNA Polymorphisms Modify Drug Response. Int. J. Environ. Res. Public Health 2016, 13, 1096. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Green, B.; Marrone, A.; Guo, Y.; Kadlubar, S.; Lin, D.; Fuscoe, J.; Pogribny, I.; Ning, B. Suppression of CYP2C9 by MicroRNA Hsa-MiR-128-3p in Human Liver Cells and Association with Hepatocellular Carcinoma. Sci. Rep. 2015, 5, 8534. [Google Scholar] [CrossRef]
- Xu, K.; Liang, X.; Shen, K.; Cui, D.; Zheng, Y.; Xu, J.; Fan, Z.; Qiu, Y.; Li, Q.; Ni, L.; et al. MiR-297 Modulates Multidrug Resistance in Human Colorectal Carcinoma by down-Regulating MRP-2. Biochem. J. 2012, 446, 291–300. [Google Scholar] [CrossRef]
- Kuang, W.-H.; Dong, Z.-Q.; Tian, L.-T.; Li, J. MicroRNA-451a, MicroRNA-34a-5p, and MicroRNA-221-3p as Predictors of Response to Antidepressant Treatment. Braz. J. Med. Biol. Res. 2018, 51, e7212. [Google Scholar] [CrossRef] [Green Version]
- Graziano, F.; Canestrari, E.; Loupakis, F.; Ruzzo, A.; Galluccio, N.; Santini, D.; Rocchi, M.; Vincenzi, B.; Salvatore, L.; Cremolini, C.; et al. Genetic Modulation of the Let-7 MicroRNA Binding to KRAS 3’-Untranslated Region and Survival of Metastatic Colorectal Cancer Patients Treated with Salvage Cetuximab-Irinotecan. Pharm. J. 2010, 10, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Boni, V.; Zarate, R.; Villa, J.C.; Bandrés, E.; Gomez, M.A.; Maiello, E.; Garcia-Foncillas, J.; Aranda, E. Role of Primary MiRNA Polymorphic Variants in Metastatic Colon Cancer Patients Treated with 5-Fluorouracil and Irinotecan. Pharm. J. 2011, 11, 429–436. [Google Scholar] [CrossRef]
- Alacam, H.; Akgun, S.; Akca, H.; Ozturk, O.; Kabukcu, B.B.; Herken, H. MiR-181b-5p, MiR-195-5p and MiR-301a-3p Are Related with Treatment Resistance in Schizophrenia. Psychiatry Res. 2016, 245, 200–206. [Google Scholar] [CrossRef]
- Duroux-Richard, I.; Pers, Y.-M.; Fabre, S.; Ammari, M.; Baeten, D.; Cartron, G.; Touitou, I.; Jorgensen, C.; Apparailly, F. Circulating MiRNA-125b Is a Potential Biomarker Predicting Response to Rituximab in Rheumatoid Arthritis. Mediators Inflamm. 2014, 2014, 342524. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, L.; Li, G.W.; Yang, Z.Y.; Lin, X.Z. [Dexmedetomidine up-regulates peripheral blood microRNA 183 to improve remifentanil-related hyperalgesia]. Zhonghua Yi Xue Za Zhi 2021, 101, 1246–1249. [Google Scholar] [CrossRef]
- Duan, K.M.; Fang, C.; Yang, S.Q.; Yang, S.T.; Xiao, J.D.; Chang, H.; Lin, G.X.; Zhang, L.B.; Peng, M.C.; Liu, Z.Q.; et al. Genetic Polymorphism of Rs13306146 Affects A2AAR Expression and Associated With Postpartum Depressive Symptoms in Chinese Women Who Received Cesarean Section. Front. Genet. 2021, 12, 959. [Google Scholar] [CrossRef]
- Yu, H.-Y.; Wang, S.-Y.; Quan, C.-X.; Fang, C.; Luo, S.-C.; Li, D.-Y.; Zhen, S.-S.; Ma, J.-H.; Duan, K.-M. Dexmedetomidine Alleviates Postpartum Depressive Symptoms Following Cesarean Section in Chinese Women: A Randomized Placebo-Controlled Study. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2019, 39, 994–1004. [Google Scholar] [CrossRef]
- Yang, X.; Chen, H.; Chen, Y.; Birnbaum, Y.; Liang, R.; Ye, Y.; Qian, J. Circulating MiRNA Expression Profiling and Target Prediction in Patients Receiving Dexmedetomidine. Cell. Physiol. Biochem. 2018, 50, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Pignataro, G. Emerging Role of MicroRNAs in Stroke Protection Elicited by Remote Postconditioning. Front. Neurol. 2021, 12, 1833. [Google Scholar] [CrossRef] [PubMed]
- Joilin, G.; Leigh, P.N.; Newbury, S.F.; Hafezparast, M. An Overview of MicroRNAs as Biomarkers of ALS. Front. Neurol. 2019, 10, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Chokkalla, A.K.; Shi, J.J.; Lee, J.; Venna, V.R.; Vemuganti, R.; McCullough, L.D. Microarray Profiling Reveals Distinct Circulating MiRNAs in Aged Male and Female Mice Subjected to Post-Stroke Social Isolation. Neuromol. Med. 2021, 23, 305–314. [Google Scholar] [CrossRef]
- Baumann, V.; Winkler, J. MiRNA-Based Therapies: Strategies and Delivery Platforms for Oligonucleotide and Non-Oligonucleotide Agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Morris-Blanco, K.C.; Lopez, M.S.; Yang, T.; Zhao, H.; Vemuganti, R.; Luo, Y. Impact of MicroRNAs on Ischemic Stroke: From Pre- to Post-Disease. Prog. Neurobiol. 2018, 163–164, 59–78. [Google Scholar] [CrossRef]
- Cuomo, O.; Cepparulo, P.; Anzilotti, S.; Serani, A.; Sirabella, R.; Brancaccio, P.; Guida, N.; Valsecchi, V.; Vinciguerra, A.; Molinaro, P.; et al. Anti-MiR-223-5p Ameliorates Ischemic Damage and Improves Neurological Function by Preventing NCKX2 Downregulation after Ischemia in Rats. Mol. Ther. Nucleic Acids 2019, 18, 1063–1071. [Google Scholar] [CrossRef] [Green Version]
- Dirnagl, U. Bench to Bedside: The Quest for Quality in Experimental Stroke Research. J. Cereb. Blood Flow Metab. 2006, 26, 1465–1478. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Effect | Mechanism/Consequences | References |
|---|---|---|
| Sedative | activates central pre- and postsynaptic α2ARs in the locus coeruleus decreases norepinephrine release mimics physiological sleep | [39] |
| Analgesic | activates α2ARs in the spinal cord–dorsal horn (locus coeruleus) decreases the release of substance P | [54,55] |
| Anti-inflammatory | ↓ TNF-α, IL-6, IL-8, IL-1β ↑ IL-10 ↑ NF-κB and CD4 cells ↓ CD8 cells ↓ plasma level of epinephrine, norepinephrine, and cortisol increases the degradation of NLRP3 inflammasome (a molecule with a role in activating caspase-1 and secreting proinflammatory cytokines such as IL-1β or IL-18) | [49,56,57,58,59] |
| Antioxidant | decreased ROS and increased SOD, GSH, and CAT after LPS or acetaminophen-induced oxidative stress | [60,61] |
| Increases blood pressure after rapid bolus injection | changes the expression α2 AR from the subtype α2A to a1 and α2B. α2A ARs are involved in vasodilation by NO production activates phospholipase A and increases the vascular response to angiotensin and catecholamines. | [54,62] |
| Decreases blood pressure after intravenous continuous infusion | activates presynaptic α2ARs decreases norepinephrine release decreases sympathetic outflow | [63,64] |
| Study | miRNA | Role of miRNA | Effect of DEX on miRNA | Biological Effect of miRNA | References |
|---|---|---|---|---|---|
| In vitro Human neuroblastoma SH-SY5Y cells—bupivacaine-induced Neuron injury | miR-7-5p | Improved mitochondrial membrane potential Increased antioxidative enzymes and reduced apoptosis Impaired bupivacaine-induced parthanatos | Upregulated | Inhibiting PARP1 ↓ SOD1, CAT ↑ GPx ↓ Bax, cleaved caspase 3 | [69] |
| In vivo Sprague-Dawley rats—postoperative cognitive dysfunction induced by sevoflurane | miR-129 | involved in cognitive dysfunction-related diseases | Increases expression in hippocampus and cortical neurons | Inhibiting TLR4 and NF-κB p65 phosphorylation | [70] |
| In vivo: APPswe/PS1dE9-induced AD mice | miR-214-5p | involved in reducing apoptosis neuroinflammation and oxidative stress of hippocampal neurons Attenuating cognitive impairments of AD mice | Upregulated expression in hippocampal neurons | Downregulated SUZ12 | [71] |
| In vivo Newborn Sprague-Dawley rats—induced HIBD | miR-29a-3p | Elevated MiR-29a-3p reduces inflammation and apoptosis rate of hippocampal neurons increase spatial learning and memory abilities Improved brain atrophy and alleviated pathological changes in ischemic rat brain | Upregulated | Negatively regulating HDAC4 ↓ IL-6, IL-8, and TNFα ↑ IL-10 | [72] |
| In vivo Wistar rats—induced CI/R injury In vitro Rat hippocampal neurons cells | miR-205-5p | May inhibit the inflammatory response and oxidative stress reduced brain infraction volume, improved neurological function score in acute cerebral ischemia induced in rats | Upregulated expression in hippocampal neurons | Negatively regulating HMGB1 ↑ SOD, ↓ ROS, MDA ↑ Nrf2, GR, GPX, HO-1, CAT ↓ IL-1β, IL-6, and TNF-α | [30] |
| In vivo Sprague-Dawley rats —CI/R injury In vitro Primary hippocampal neuron cell culture exposed to OGD/R treatment | miR-381 | inhibited inflammation response and neuron cell apoptosis improved neurological function | Upregulated | Inhibiting IRF4-IL-9 | [73] |
| In vivo C57BL/6 mice—sevoflurane-induced POCD In vitro primary hippocampal neurons from newborn mice | involved in apoptosis, neuroinflammation, and DNA damage repair Alleviates Cognitive Dysfunction | Upregulated | Inhibiting EGR1/p53 Disrupts interaction of EGR1 with p53 Decreasing EGR1-mediated p53 transcription | [74] | |
| In vitro cell culture (Pheochromocytoma cell line—PC12)—ropivacaine-induced neuronal injury | may reduce the proliferation and apoptosis of ropivacaine-induced PC12 cells | Upregulated | Negatively regulating LRRC4 expression ↓ Cleaved-Caspase-3 ↑ Bcl-2 | [31] | |
| In vivo Wistar rats—lipopolysaccharide (LPS)-induced inflammation | miR-21-5p | involved in inflammation, autophagy, and apoptosis | Decreased the expression in the spleen and hippocampus | Attenuated PARP cleavage in the spleen | [75] |
| miR-204-5p | |||||
| miR-30a-5p | |||||
| In vivo Wistar rats—LPS-induced neuroinflammation | miR-124 | involved in neuronal differentiation | Decreased the expression in the hippocampus and cortex | Targeted SHIP1, SOCS1 | [76] |
| miR-132 | involved in the inflammatory response in alveolar macrophages potentiated cholinergic anti-inflammatory pathway | ||||
| miR-155 | involved in pro- and anti-inflammatory mechanisms | ||||
| In vivo 40 patients undergoing off-pump coronary artery bypass grafting | miR-320a | associated with cell proliferation | Decreased miRNA-320 expression level in the blood of patients after treatment with DEX | Stimulated NGB expression | [77] |
| In vivo Adult male Sprague-Dawley rats- chronic inflammatory visceral pain | miR-211 | Down-regulated in TNBS-induced chronic inflammatory visceral pain | Upregulated after DEX treatment | Negatively regulating ERK expression ↓ IL-1β, TNF-α, and IL-6 | [78] |
| In vivo Adult male Sprague-Dawley rats In vitro primary myoblasts from young mice In vivo Coopworth ewes sheep | rno-miR-434-3p | miR-434-3p protects myocytes from apoptosis | Upregulated | Targeted EIF5A G2E3, DCAF6, and TMEM68 ↓ caspases-3, −8 and −9 | [79,80] |
| rno-miR-3596d | Targeted G2E3, DCAF6, TMEM68, ATAD2B, NPY1R, SRSF1, ITGA6, MORC3, and RSF1 | ||||
| rno-miR-496-5p | Controlled mTOR pathway Targeted ATAD2B, NPY1R, SRSF1, ITGA6, MORC3, and RSF1 | ||||
| rno-miR-7a-2-3p | Negatively regulating PARP expression | ||||
| rno-miR-702-3p | |||||
| rno-miR-208b-3p | Upregulated in heart failure | Downregulated | Targeted CSNK2A2/NLK Promoted Wnt/β-catenin pathway ↑ Bcl-2 | [79,81] | |
| In vivo Sprague-Dawley rats—MI/R injury | miR-346-3p | miRNA-346 suppresses infarct size inhibits myocardial cell apoptosis may protect against MI/R injury | Upregulated the expression | Negatively regulated CAMK2D expression ↓ NF-κB ↓ NLRP3 inflammasome | [67] |
| In vivo LPS-induced acute lung injury of mice | miR-223-3p | The absence is associated with severe lung inflammation pulmonary up-regulation of this miRNA in mice may provide protection during acute lung injury due to various causes (eg. mechanical ventilation) | Upregulated | Negatively regulated HDAC4 expression ↓ TLR4 ↓ NF-κB | [82] |
| In vivo neonatal mice model—induced HIBD | mmu-miR-128 | Increased the neuroprotective effects of DEX against ischemic brain injury attenuated nerve cell apoptosis enhanced learning and memory abilities reduced left-brain water content | Upregulated | Negatively regulating WNT1 expression | [83] |
| miRNAs | Type of Study | Intervention | Observations | References |
|---|---|---|---|---|
| miR-320 | 40 patients | Off-pump coronary artery bypass grafting | Downregulated levels after treatment | [77] |
| miR-183 | 80 patients | Laparoscopic cholecystectomy and oophorocystectomy surgery | Overexpressed after treatment | [220] |
| miR-646--α2AAR s13306146 polymorphism | 568 cesarean section patients | Chinese women who received cesarean section | miR-646 level in α2AAR s13306146 polymorphism affects α2AAR Postpartum Depressive Symptoms | [221] |
| miR-30a-5p, -101-3p, -140-3p and -141-3p | 133 pediatric patients | Preoperative sedation for different procedures | Increased levels of miR-101-3p and 140-3p in respondents and downregulated levels of miR-101-3p and miR-30a-5p in hypotension and bradycardia patients | [33] |
| hsa-miR-4508, -novel-chr8_87373, -30a-3p, -novel-chr16_26099, -4306, -744-5p, -320a, -novel-chr9_90035, -101-3p, -150-5p, -342-3p, and-140-3p | three patients | Elective surgery | Five miRNAs upregulated (hsa-miR-4508, -novel-chr8_87373, -30a-3p, -novel-chr16_26099, -4306,) and seven miRNAs downregulated (hsa-miR-744-5p, -320a, -novel-chr9_90035, -101-3p, -150-5p, -342-3p, and -140-3p) after DEX treatment | [223] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burlacu, C.-C.; Neag, M.-A.; Mitre, A.-O.; Sirbu, A.-C.; Badulescu, A.-V.; Buzoianu, A.-D. The Role of miRNAs in Dexmedetomidine’s Neuroprotective Effects against Brain Disorders. Int. J. Mol. Sci. 2022, 23, 5452. https://doi.org/10.3390/ijms23105452
Burlacu C-C, Neag M-A, Mitre A-O, Sirbu A-C, Badulescu A-V, Buzoianu A-D. The Role of miRNAs in Dexmedetomidine’s Neuroprotective Effects against Brain Disorders. International Journal of Molecular Sciences. 2022; 23(10):5452. https://doi.org/10.3390/ijms23105452
Chicago/Turabian StyleBurlacu, Codrin-Constantin, Maria-Adriana Neag, Andrei-Otto Mitre, Alexandru-Constantin Sirbu, Andrei-Vlad Badulescu, and Anca-Dana Buzoianu. 2022. "The Role of miRNAs in Dexmedetomidine’s Neuroprotective Effects against Brain Disorders" International Journal of Molecular Sciences 23, no. 10: 5452. https://doi.org/10.3390/ijms23105452