
Genome-Wide Association Study Identifies Candidate Genes Related to the Linoleic Acid Content in Soybean Seeds


Abstract
:1. Introduction
2. Results
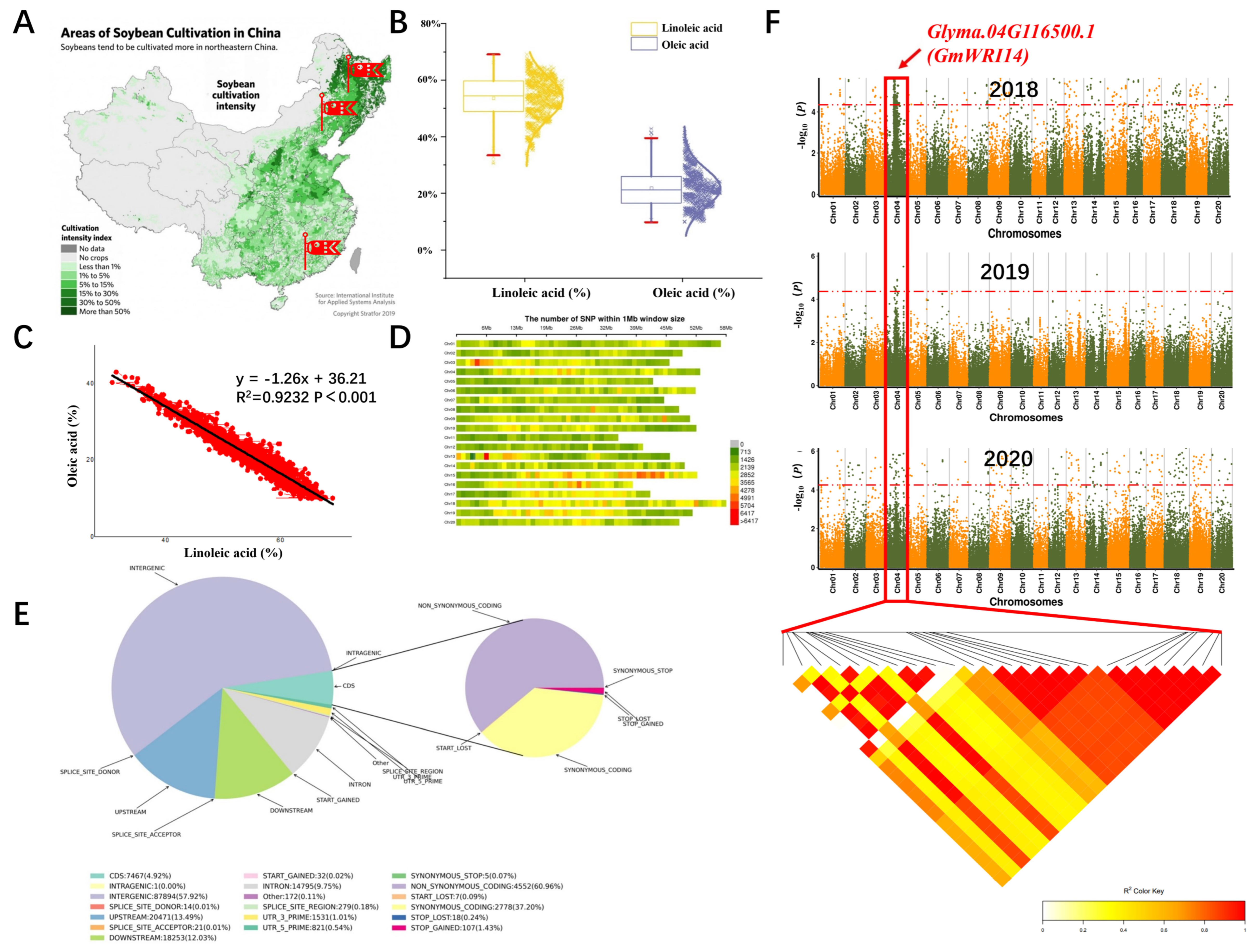
2.1. Phenotypic Variation of LA Content in Soybean Seeds
2.2. A New Candidate Gene Related to LA Was Discovered by GWAS in 2018–2020
2.3. The Expression of Candidate Gene GmWRI14 in Different Tissues and Developmental Stages of Soybean
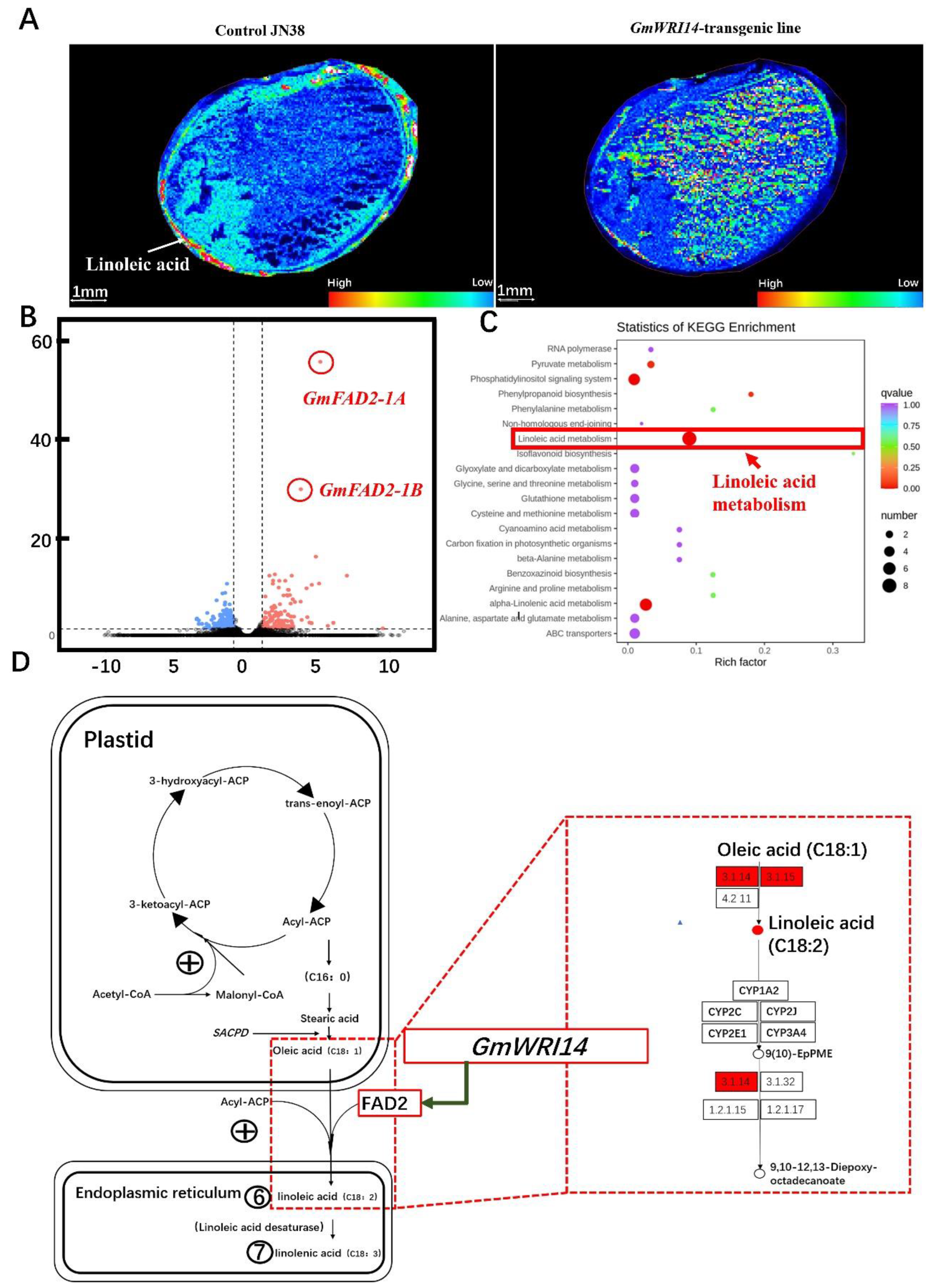
2.4. The Generation and Molecular Characterization of Transgenic Soybean Plants Over-Expressing GmWRI14
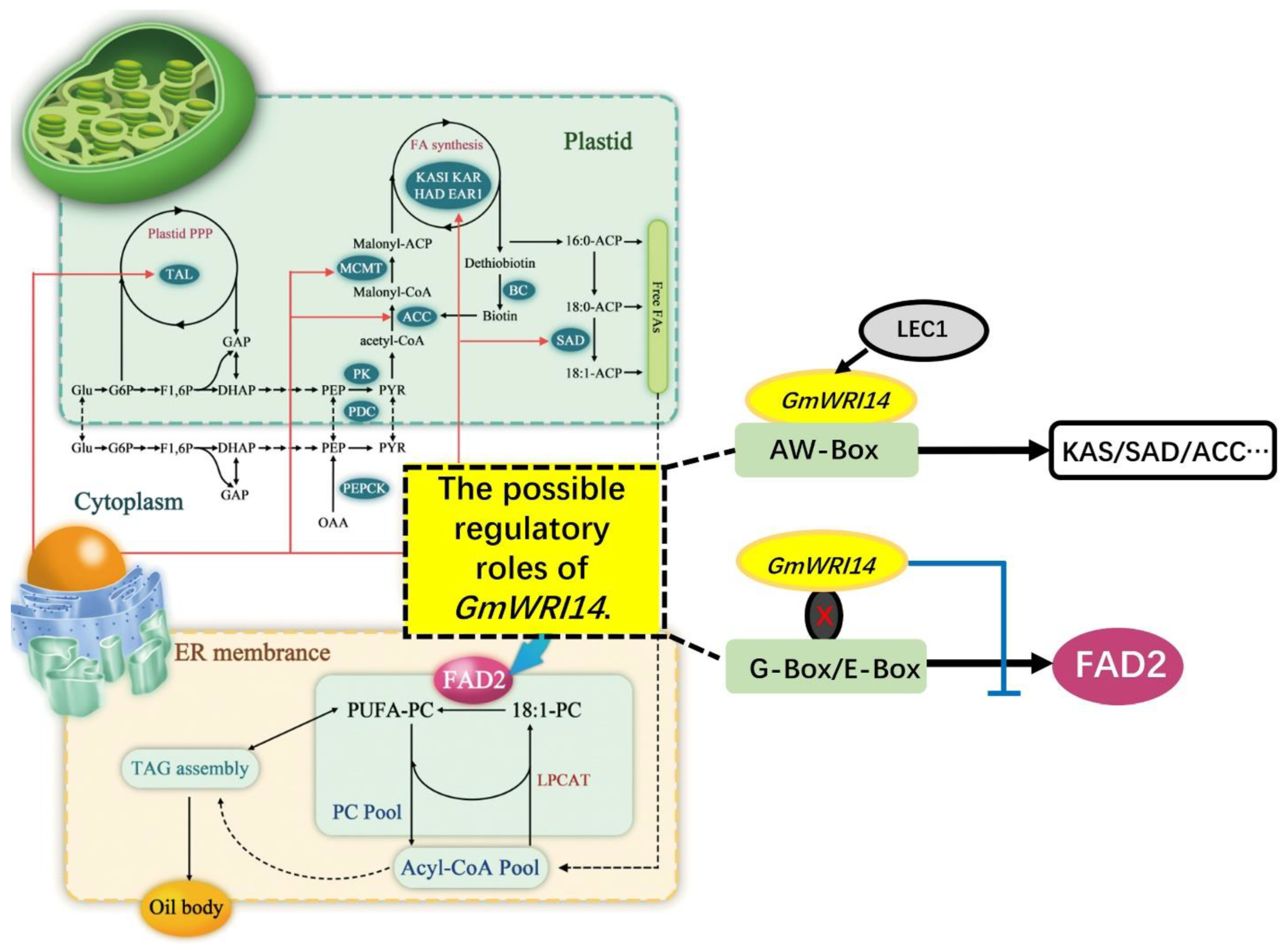
2.5. The Reduction in LA Triggered by GmWRI14 Expression Is Due to the Down-Regulation of Soybean FAD2
3. Discussion
4. Materials and Methods
4.1. The Plant Materials
4.2. Determination of Fatty Acids in Soybean Seeds
4.3. Genotyping of Soybean Germplasms
4.4. The Genome-Wide Association Analysis (GWAS)
4.5. Quantitative Reverse Transcription-PCR
4.6. Vector Construction and Plant Transformation
4.7. The RNA-Seq Library Preparation and Sequencing
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Qin, D.; Piersanti, A.; Zhang, Q.; Miceli, C.; Wang, P. Genome-wide association study identifies candidate genes related to oleic acid content in soybean seeds. BMC Plant. Biol. 2020, 20, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, H.; Feng, L.; Qu, Y.; Teng, W.; Qiu, L.; Zheng, H.; Han, Y.; Li, W. Genome-wide association and transcriptional studies reveal novel genes for unsaturated fatty acid synthesis in a panel of soybean accessions. BMC Genom. 2019, 20, 68–84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, J.; Wang, H.; Meng, S.; Xing, G.; Li, Y.; Yang, S.; Zhao, J.; Zhao, T.; Gai, J. Detecting the QTL-allele system of seed oil traits using multi-locus genome-wide association analysis for population characterization and optimal cross prediction in soybean. Front. Plant Sci. 2018, 9, 1793. [Google Scholar] [CrossRef] [Green Version]
- Clemente, T.E.; Cahoon, E.B. Soybean oil: Genetic approaches for modification of functionality and total content. Plant. Physiol. 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- De Groot, R.H.; Hornstra, G.; van Houwelingen, A.C.; Roumen, F. Effect of α-linolenic acid supplementation during pregnancy on maternal and neonatal polyunsaturated fatty acid status and pregnancy outcome. Am. J. Clin. Nutr. 2004, 79, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.K.; Bruce, V.M.; McDonald, B.E. Dietary α-linolenic acid is as effective as oleic acid and linoleic acid in lowering blood cholesterol in normolipidemic men. Am. J. Clin. Nutr. 1991, 53, 1230–1234. [Google Scholar] [CrossRef]
- Maskan, M.; Karataş, Ş. Fatty acid oxidation of pistachio nuts stored under various atmospheric conditions and different temperatures. J. Sci. Food Agric. 1998, 77, 334–340. [Google Scholar] [CrossRef]
- Yang, L.; Cao, Y.; Chen, J.-N.; Chen, Z.-Y. Oxidative stability of conjugated linolenic acids. J. Agric. Food Chem. 2009, 57, 4212–4217. [Google Scholar] [CrossRef]
- Tompkins, C.; Perkins, E.G. Frying performance of low-linolenic acid soybean oil. J. Am. Oil Chem. Soc. 2000, 77, 223–229. [Google Scholar] [CrossRef]
- Marangoni, F.; Agostoni, C.; Borgh, C.; Catapano, A.L.; Cena, H.; Ghiselli, A.; La Vecchia, C.; Lercker, G.; Manzato, E.; Pirillo, A. Dietary linoleic acid and human health: Focus on cardiovascular and cardiometabolic effects. Atherosclerosis 2020, 292, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, C.; Rasmussen, O.; Lousen, T.; Holst, J.J.; Fenselau, S.; Schrezenmeir, J.; Hermansen, K. Differential effects of saturated and monounsaturated fatty acids on postprandial lipemia and incretin responses in healthy subjects. Am. J. Clin. Nutr. 1999, 69, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Do, P.T.; Nguyen, C.X.; Bui, H.T.; Tran, L.T.; Stacey, G.; Gillman, J.D.; Zhang, Z.J.; Stacey, M.G. Demonstration of highly efficient dual gRNA CRISPR/Cas9 editing of the homeologous GmFAD2–1A and GmFAD2–1B genes to yield a high oleic, low linoleic and α-linolenic acid phenotype in soybean. BMC Plant Biol. 2019, 19, 311–325. [Google Scholar] [CrossRef]
- Singh, S.; Thomaeus, S.; Lee, M.; Stymne, S.; Green, A. Transgenic expression of a Δ12-epoxygenase gene in Arabidopsis seeds inhibits accumulation of linoleic acid. Planta 2001, 212, 872–879. [Google Scholar] [CrossRef]
- Sivaraman, I.; Arumugam, N.; Sodhi, Y.S.; Gupta, V.; Mukhopadhyay, A.; Pradhan, A.K.; Burma, P.K.; Pental, D. Development of high oleic and low linoleic acid transgenics in a zero erucic acid Brassica juncea L. (Indian mustard) line by antisense suppression of the FAD2 gene. Mol. Breed. 2004, 13, 365–375. [Google Scholar] [CrossRef]
- Li, H.; Thrash, A.; Tang, J.D.; He, L.; Yan, J.; Warburton, M.L. Leveraging GWAS data to identify metabolic pathways and networks involved in maize lipid biosynthesis. Plant. J. 2019, 98, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Lakhssassi, N.; Zhou, Z.; Liu, S.; Colantonio, V.; AbuGhazaleh, A.; Meksem, K. Characterization of the FAD2 gene family in soybean reveals the limitations of gel-based TILLING in genes with high copy number. Front. Plant Sci. 2017, 8, 324. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Liu, H.; Li, X.; Chen, X.; Hong, Y.; Li, H.; Lu, Q.; Liang, X. TALEN-mediated targeted mutagenesis of fatty acid desaturase 2 (FAD2) in peanut (Arachis hypogaea L.) promotes the accumulation of oleic acid. Plant. Mol. Biol. 2018, 97, 177–185. [Google Scholar] [CrossRef]
- Yang, J.; Xing, G.; Niu, L.; He, H.; Guo, D.; Du, Q.; Qian, X.; Yao, Y.; Li, H.; Zhong, X. Improved oil quality in transgenic soybean seeds by RNAi-mediated knockdown of GmFAD2-1B. Transgenic Res. 2018, 27, 155–166. [Google Scholar] [CrossRef]
- Povkhova, L.; Pushkova, E.; Dmitriev, A.; Kezimana, P.; Novakovskiy, R.; Melnikova, N.; Rozhmina, T.; Krasnov, G. Molecular markers based on SNPs in FAD3 genes for determination of linolenic acid content in flax seed. Bioinform. Genome Regul. Struct./Syst. Biol. 2020, 35, 334–335. [Google Scholar]
- Thapa, R.; Carrero-Colón, M.; Addo-Quaye, C.; Held, J.; Dilkes, B.; Hudson, K.A. New alleles of FAD3A lower the linolenic acid content of soybean seeds. Crop. Sci. 2018, 58, 713–718. [Google Scholar] [CrossRef]
- Held, J.P.; Carrero-Colón, M.; Hudson, K.A. Combination of novel mutation in FAD3C and FAD3A for low linolenic acid soybean. Agrosystems Geosci. Environ. 2019, 2, 6–10. [Google Scholar] [CrossRef]
- Kong, Q.; Yuan, L.; Ma, W. Wrinkled1, a “Master Regulator” in transcriptional control of plant oil biosynthesis. Plants 2019, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Yang, Y.; Guo, L.; Yuan, L.; Ma, W. Molecular basis of plant oil biosynthesis: Insights gained from studying the WRINKLED1 transcription factor. Front. Plant Sci. 2020, 291, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Du, C.; Song, H.; Aziz, U.; Wang, L.; Zhao, C.; Zhang, M. Genome-wide analysis reveals the evolution and structural features of Wrinkled1 in plants. Mol. Genet. Genom. 2019, 294, 329–341. [Google Scholar] [CrossRef]
- Kuczynski, C.; McCorkle, S.; Keereetaweep, J.; Shanklin, J.; Schwender, J. An expanded role for WRINKLED1 metabolic control based on combined phylogenetic and biochemical analyses. Bio Rxiv 2020, 56, 231–242. [Google Scholar]
- Guo, W.; Chen, L.; Chen, H.; Yang, H.; You, Q.; Bao, A.; Chen, S.; Hao, Q.; Huang, Y.; Qiu, D. Overexpression of GmWRI1b in soybean stably improves plant architecture and associated yield parameters, and increases total seed oil production under field conditions. Plant. Biotechnol. J. 2020, 18, 1639–1641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hong, M.; Wan, H.; Luo, L.; Yu, Z.; Guo, R. Identification of key genes involved in embryo development and differential oil accumulation in two contrasting maize genotypes. Genes 2019, 10, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, P.A.; Bayon de Noyer, S.; Park, H.; Nguyen, H.; Hou, L.; Changa, T.; Khang, H.L.; Ciftci, O.N.; Wang, T.; Cahoon, E.B. Expression of the Arabidopsis WRINKLED 1 transcription factor leads to higher accumulation of palmitate in soybean seed. Plant. Biotechnol. J. 2019, 17, 1369–1379. [Google Scholar] [CrossRef]
- Kong, Q.; Ma, W. Wrinkled1 transcription factor: How much do we know about its regulatory mechanism? Plant. Sci. 2018, 272, 153–156. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, Y.; Dong, Z.; Meng, F.; Sun, X.; Fan, X.; Zhang, Y.; Wang, M.; Wang, S. Soybean (Glycine max) Wrinkled1 transcription factor, GmWRI1a, positively regulates seed oil accumulation. Mol. Genet. Genom. 2018, 293, 401–415. [Google Scholar] [CrossRef]
- Svishcheva, G.R.; Axenovich, T.I.; Belonogova, N.M.; Van Duijn, C.M.; Aulchenko, Y.S. Rapid variance components–based method for whole-genome association analysis. Nat. Genet. 2012, 44, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.; Chen, P.; Korth, K.; Hancock, F.; Pereira, A.; Brye, K.; Wu, C.; Shi, A. Genome-wide association study (GWAS) of salt tolerance in worldwide soybean germplasm lines. Mol. Breed. 2017, 37, 30–44. [Google Scholar] [CrossRef]
- Wang, L.; Yang, Y.; Zhang, S.; Che, Z.; Yuan, W.; Yu, D. GWAS reveals two novel loci for photosynthesis-related traits in soybean. Mol. Genet. Genom. 2020, 295, 345–356. [Google Scholar] [CrossRef]
- Yu, Z.; Chang, F.; Lv, W.; Sharmin, R.A.; Wang, Z.; Kong, J.; Bhat, J.A.; Zhao, T. Identification of QTN and candidate gene for seed-flooding tolerance in soybean [Glycine max (L.) Merr.] using genome-wide association study (GWAS). Genes 2019, 10, 957. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wu, T.; Wang, L.; Jiang, B.; Zhen, C.; Yuan, S.; Hou, W.; Wu, C.; Han, T.; Sun, S. A combined linkage and GWAS analysis identifies QTLs linked to soybean seed protein and oil content. Int. J. Mol. Sci. 2019, 20, 5915. [Google Scholar] [CrossRef] [Green Version]
- Whelan, J.; Fritsche, K. Linoleic acid. Adv. Nutr. 2013, 4, 311–312. [Google Scholar] [CrossRef]
- Hammond, E.G.; Johnson, L.A.; Su, C.; Wang, T.; White, P.J. Soybean oil. Bailey’s Ind. Oil Fat Prod. 2005, 32, 114–124. [Google Scholar]
- Cerna, A.; Benning, C. Wrinkled1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 2004, 40, 575–585. [Google Scholar] [CrossRef]
- An, D.; Suh, M.C. Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content in Camelina sativa. Plant Biotechnol. Rep. 2015, 9, 137–148. [Google Scholar] [CrossRef]
- Baud, S.; Wuillème, S.; To, A.; Rochat, C.; Lepiniec, L. Role of WRINKLED1 in the transcriptional regulation of glycolytic and fatty acid biosynthetic genes in Arabidopsis. Plant J. 2009, 60, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, A.C.; Ario, N.; Tomita, Y.; Tanaka, S.; Murayama, N.; Mizuta, C.; Iwaya-Inoue, M.; Ishibashi, Y. High temperature during soybean seed development differentially alters lipid and protein metabolism. Plant Prod. Sci. 2020, 23, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Aladedunye, F.A.; Przybylski, R. Degradation and nutritional quality changes of oil during frying. J. Am. Oil Chem. Soc. 2009, 86, 149–156. [Google Scholar] [CrossRef]
- Robards, K.; Kerr, A.F.; Patsalides, E. Rancidity and its measurement in edible oils and snack foods, A review. Analyst 1988, 113, 213–224. [Google Scholar] [CrossRef]
- Esfarjani, F.; Khoshtinat, K.; Zargaraan, A.; Mohammadi-Nasrabadi, F.; Salmani, Y.; Saghafi, Z.; Hosseini, H.; Bahmaei, M. Evaluating the rancidity and quality of discarded oils in fast food restaurants. Food Sci. Nutr. 2019, 7, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Holsters, M.; De Waele, D.; Depicker, A.; Messens, E.; Van Montagu, M.; Schell, J. Transfection and transformation of Agrobacterium tumefaciens. Mol. Gen. Genet. MGG 1978, 163, 181–187. [Google Scholar] [CrossRef]
- Zhong, S.; Joung, J.-G.; Zheng, Y.; Chen, Y.; Liu, B.; Shao, Y.; Xiang, J.Z.; Fei, Z.; Giovannoni, J.J. High-throughput illumina strand-specific RNA sequencing library preparation. Cold Spring Harb. Protoc. 2011, 8, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, M.; Dökel, S.; Amstislavskiy, V.; Wuttig, D.; Sültmann, H.; Lehrach, H.; Yaspo, M.-L. A simple strand-specific RNA-Seq library preparation protocol combining the Illumina TruSeq RNA and the dUTP methods. Biochem. Biophys. Res. Commun. 2012, 422, 643–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Ichihashi, Y.; Kimura, S.; Chitwood, D.H.; Headland, L.R.; Peng, J.; Maloof, J.N.; Sinha, N.R. A high-throughput method for Illumina RNA-Seq library preparation. Front. Plant Sci. 2012, 3, 202. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Correlation Coefficient with WRI1 | Soybean Name | Relative Expression in Leaves | Relative Expression in Stem | Relative Expression in Roots | Relative Expression in Seed | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Sig. | Mean | Sig. | Mean | Sig. | Mean | Sig. | |||
| GmFAD2-1A | −0.970–−0.982 | JN38 | 10.15 ± 0.34 | c | 36.11 ± 0.42 | b | 32.21 ± 0.12 | a | 35.11 ± 0.25 | a |
| JN38-GmWRI14-1 | 5.31 ± 0.24 | d | 14.23 ± 0.21 | c | 13.23 ± 1.2 | c | 19.23 ± 0.54 | c | ||
| JN38-GmWRI14-2 | 2.52 ± 0.32 | d | 10.21 ± 0.51 | c | 23.23 ± 0.12 | b | 17.32 ± 0.42 | c | ||
| JN38-GmWRI14-3 | 3.12 ± 0.01 | d | 19.23 ± 0.15 | c | 29.11 ± 0.51 | b | 22.23 ± 0.56 | b | ||
| JN38-GmWRI14-4 | 4.25 ± 0.12 | d | 15.22 ± 0.16 | c | 16.23 ± 0.23 | c | 26.11 ± 0.16 | b | ||
| JN38-GmWRI14-5 | 3.22 ± 0.02 | d | 18.25 ± 0.31 | c | 20.54 ± 0.26 | b | 23.22 ± 0.61 | b | ||
| JN38-GmWRI14-6 | 1.33 ± 0.01 | d | 18.22 ± 0.25 | c | 21.51 ± 0.16 | b | 19.23 ± 0.12 | c | ||
| GmFAD2-2B | −0.880–−0.814 | JN38-GmWRI14-1 | 6.23 ± 0.21 | d | 3.14 ± 0.17 | d | 11.95 ± 0.54 | c | 22.75 ± 0.26 | b |
| JN38-GmWRI14-2 | 5.46 ± 0.11 | d | 4.22 ± 0.22 | d | 12.51 ± 0.34 | c | 21.21 ± 0.13 | b | ||
| JN38-GmWRI14-3 | 6.45 ± 0.15 | d | 4.42 ± 0.2 | d | 11.11 ± 0.54 | c | 26.12 ± 0.26 | b | ||
| JN38-GmWRI14-4 | 7.23 ± 0.17 | d | 5.12 ± 0.15 | d | 13.21 ± 0.32 | c | 25.15 ± 0.65 | b | ||
| JN38-GmWRI14-5 | 7.51 ± 0.22 | d | 4.78 ± 0.25 | d | 9.51 ± 0.26 | d | 27.35 ± 0.43 | b | ||
| JN38-GmWRI14-6 | 9.11 ± 0.12 | d | 3.22 ± 0.19 | d | 12.43 ± 0.62 | c | 26.21 ± 0.22 | b | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, Q.; Piersanti, A.; Zhang, Q.; Miceli, C.; Li, H.; Liu, X. Genome-Wide Association Study Identifies Candidate Genes Related to the Linoleic Acid Content in Soybean Seeds. Int. J. Mol. Sci. 2022, 23, 454. https://doi.org/10.3390/ijms23010454
Di Q, Piersanti A, Zhang Q, Miceli C, Li H, Liu X. Genome-Wide Association Study Identifies Candidate Genes Related to the Linoleic Acid Content in Soybean Seeds. International Journal of Molecular Sciences. 2022; 23(1):454. https://doi.org/10.3390/ijms23010454
Chicago/Turabian StyleDi, Qin, Angela Piersanti, Qi Zhang, Cristina Miceli, Hui Li, and Xiaoyi Liu. 2022. "Genome-Wide Association Study Identifies Candidate Genes Related to the Linoleic Acid Content in Soybean Seeds" International Journal of Molecular Sciences 23, no. 1: 454. https://doi.org/10.3390/ijms23010454