
TGF-β1 Potentiates the Cytotoxicity of Cadmium by Induction of a Metal Transporter, ZIP8, Mediated by the ALK5-Smad2/3 and ALK5-Smad3-p38 MAPK Signal Pathways in Cultured Vascular Endothelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Influence of TGF-β1 on Cadmium Cytotoxicity towards Vascular Endothelial Cells
2.2. TGF-β1 Specifically Induces ZIP8 Expression in Vascular Endothelial Cells
2.3. TGF-β1 Increases RSS Levels in Vascular Endothelial Cells
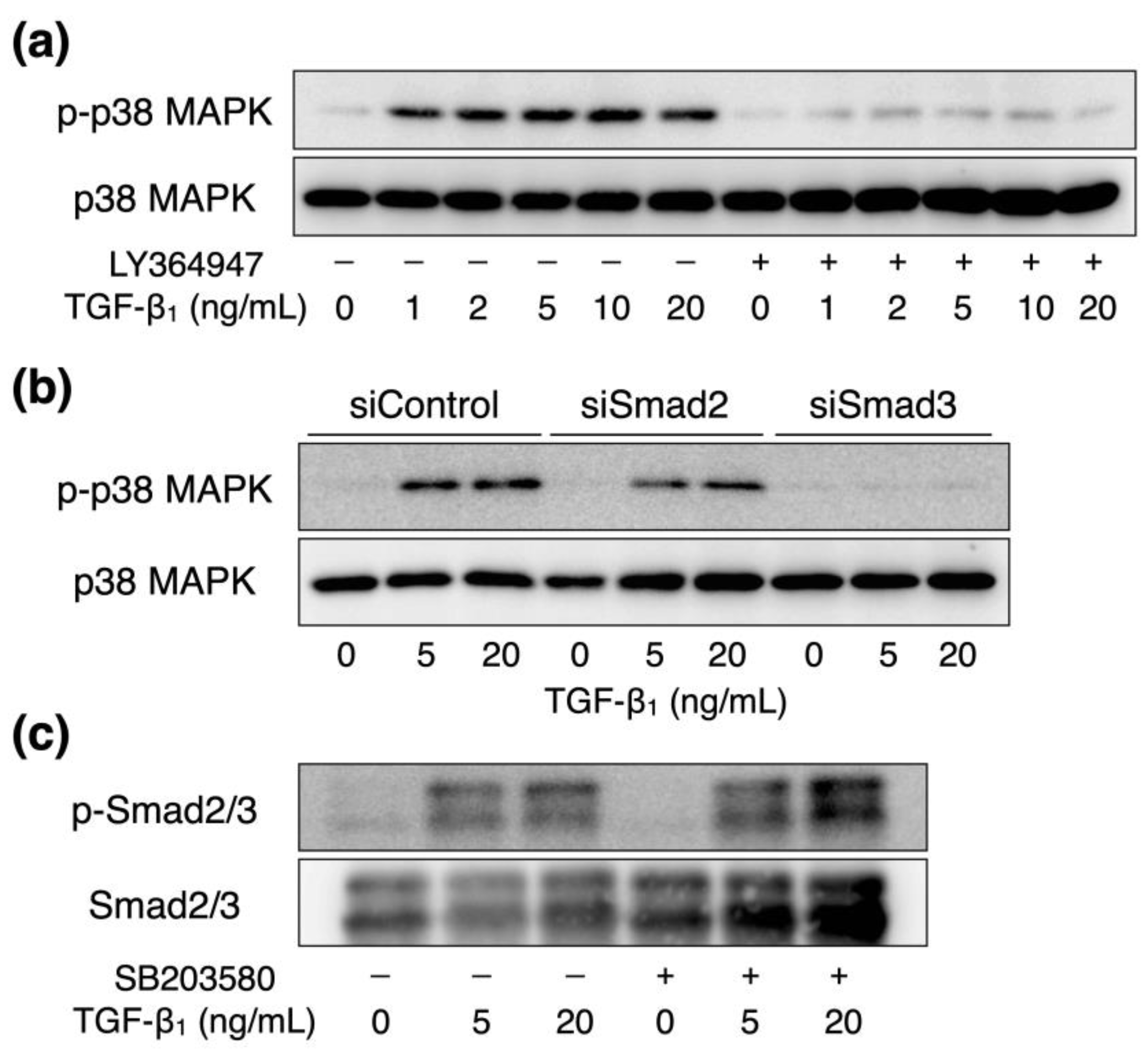
2.4. Interaction between TGF-β1-Activated Smad2/3 and p38 MAPK Signaling in Vascular Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatment
4.3. Small Interfering RNA Transfection
4.4. Morphological Appearance and Lactate Dehydrogenase Assay
4.5. Intracellular Accumulation of Cadmium
4.6. Extraction of the Membrane Fraction
4.7. Western Blot Analysis
4.8. Real-Time Reverse Transcription Polymerase Chain Reaction
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Palmer, R.M.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Weksler, B.B.; Marcus, A.J.; Jaffe, E.A. Synthesis of prostaglandin I2 (prostacyclin) by cultured human and bovine endothelial cells. Proc. Natl. Acad. Sci. USA 1977, 74, 3922–3926. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Levin, E.G.; Louskutoff, D.J. Cultured bovine endothelial cells produce both urokinase and tissue-type plasminogen activators. J. Cell Biol. 1982, 94, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelehrter, T.D.; Sznycer-Laszuk, R. Thrombin induction of plasminogen activator-inhibitor in cultured human endothelial cells. J. Clin. Investig. 1986, 77, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Ross, R. Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Toma, I.; McCaffrey, T.A. Transforming growth factor-β and atherosclerosis: Interwoven atherogenic and atheroprotective aspects. Cell Tissue Res. 2012, 347, 155–175. [Google Scholar] [CrossRef] [Green Version]
- Fràter-Schröder, M.; Müller, G.; Birchmeier, W.; Böhlen, P. Transforming growth factor-beta inhibits endothelial cell proliferation. Biochem. Biophys. Res. Commun. 1986, 137, 295–302. [Google Scholar] [CrossRef]
- Saksela, O.; Moscatelli, D.; Rifkin, D.B. The opposing effects of basic fibroblast growth factor and transforming growth factor beta on the regulation of plasminogen activator activity in capillary endothelial cells. J. Cell Biol. 1987, 105, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Kaji, T.; Yamada, A.; Miyajima, S.; Yamamoto, C.; Fujiwara, Y.; Wight, T.N.; Kinsella, M.G. Cell density-dependent regulation of proteoglycan synthesis by transforming growth factor-β1 in cultured bovine aortic endothelial cells. J. Biol. Chem. 2000, 275, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Yoshida, E.; Fujiwara, Y.; Yamamoto, C.; Kaji, T. Transforming growth factor-β1 modulates the expression of syndecan-4 in cultured vascular endothelial cells in a biphasic manner. J. Cell. Biochem. 2017, 118, 2009–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.P.; Seki, T.; Goss, K.A.; Imamura, T.; Yi, Y.; Donahoe, P.K.; Li, L.; Miyazono, K.; ten Dijke, P.; Kim, S.; et al. Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2626–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goumans, M.-J.; Valdimarsdottir, G.; Itoh, S.; Rosendahl, A.; Sideras, P.; ten Dijke, P. Balancing the activation state of the endothelium via two distinct TGF-beta type I receptors. EMBO J. 2002, 21, 1743–1753. [Google Scholar] [CrossRef] [PubMed]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Goumans, M.-J.; Valdimarsdottir, G.; Itoh, S.; Lebrin, F.; Larsson, J.; Mummery, C.; Karlsson, S.; ten Dijke, P. Activin receptor-like kinase (ALK)1 is an antagonistic mediator of lateral TGFbeta/ALK5 signaling. Mol. Cell 2003, 12, 817–828. [Google Scholar] [CrossRef]
- Nogawa, K.; Yamada, Y.; Honda, R.; Ishizaki, M.; Tsuritani, I.; Kawano, S.; Kato, T. The relationship between itai-itai disease among inhabitants of the Jinzu River basin and cadmium in rice. Toxicol. Lett. 1983, 17, 263–266. [Google Scholar] [CrossRef]
- Navas-Acien, A.; Selvin, E.; Sharrett, A.R.; Calderon-Aranda, E.; Silbergeld, E.; Guallar, E. Lead, cadmium, smoking, and increased risk of peripheral arterial disease. Circulation 2004, 109, 3196–3201. [Google Scholar] [CrossRef] [Green Version]
- Kaji, T.; Mishima, A.; Koyanagi, E.; Yamamoto, C.; Sakamoto, M.; Kozuka, H. Possible mechanism for zinc protection against cadmium cytotoxicity in cultured vascular endothelial cells. Toxicology 1992, 76, 257–270. [Google Scholar] [CrossRef]
- Yamamoto, C.; Kaji, T.; Sakamoto, M.; Kozuka, H. Cadmium stimulation of plasminogen activator inhibitor-1 release from human vascular endothelial cells in culture. Toxicology 1993, 83, 215–223. [Google Scholar] [CrossRef]
- Hara, T.; Sakuma, M.; Fujie, T.; Kaji, T.; Yamamoto, C. Cadmium induces plasminogen activator inhibitor-1 via Smad2/3 signaling pathway in human endothelial EA.hy926 cells. J. Toxicol. Sci. 2021, 46, 249–253. [Google Scholar] [CrossRef]
- Ohkawara, S.; Yamamoto, C.; Fujiwara, Y.; Sakamoto, M.; Kaji, T. Cadmium induces the production of high molecular weight heparan sulfate proteoglycan molecules in cultured vascular endothelial cells. Environ. Toxicol. Pharmacol. 1997, 3, 187–194. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Yamamoto, C.; Yoshida, E.; Kumagai, Y.; Kaji, T. Heparan sulfate chains potentiate cadmium cytotoxicity in cultured vascular endothelial cells. Arch. Toxicol. 2016, 90, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Himeno, S.; Yanagiya, T.; Fujishiro, H. The role of zinc transporters in cadmium and manganese transport in mammalian cells. Biochimie 2009, 91, 1218–1222. [Google Scholar] [CrossRef]
- Dalton, T.P.; He, L.; Wang, B.; Miller, M.L.; Jin, L.; Stringer, K.F.; Chang, X.; Baxter, C.S.; Nebert, D.W. Identification of mouse SLC39A8 as the transporter responsible for cadmium-induced toxicity in the testis. Proc. Natl. Acad. Sci. USA 2005, 102, 3401–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujie, T.; Ito, K.; Ozaki, Y.; Takahashi, S.; Yamamoto, C.; Kaji, T. Induction of ZIP8, a ZIP transporter, via NF-κB signaling by the activation of IκBα and JNK signaling in cultured vascular endothelial cells exposed to cadmium. Toxicol. Appl. Pharmacol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiologic regulation. Annu. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L. The function of metallothionein. Neurochem. Int. 1995, 27, 23–33. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Cousins, R.J. Mammalian zinc transporters. Annu. Rev. Nutr. 2004, 24, 151–172. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Fan, Y.; Yang, L.; Wang, L.; Ma, J. Zinc supplementation attenuates high glucose-induced epithelial-to-mesenchymal transition of peritoneal mesothelial cells. Biol. Trace Elem. Res. 2012, 150, 229–235. [Google Scholar] [CrossRef]
- Liu, M.J.; Bao, S.; Gálvez-Peralta, M.; Pyle, C.J.; Rudawsky, A.C.; Pavlovicz, R.E.; Killilea, D.W.; Li, C.; Nebert, D.W.; Wewers, M.D.; et al. ZIP8 regulates host defense through zinc-mediated inhibition of NF-κB. Cell Rep. 2013, 3, 386–400. [Google Scholar] [CrossRef] [Green Version]
- Johansson, A.; Eriksson, N.; Lindholm, D.; Varenhorst, C.; James, S.; Syvänen, A.C.; Axelsson, T.; Siegbahn, A.; Barratt, B.J.; Becker, R.C.; et al. Genome-wide association and Mendelian randomization of NT-proBNP levels in patients with acute coronary syndrome. Hum. Mol. Genet. 2016, 25, 1447–1456. [Google Scholar] [CrossRef]
- Hill, C.S. Transcriptional control by the SMADs. Cold Spring Harb. Perspect. Biol. 2016, 8, a022079. [Google Scholar] [CrossRef] [Green Version]
- Aragón, E.; Wang, Q.; Zou, Y.; Morgani, S.M.; Ruiz, L.; Kaczmarska, Z.; Su, J.; Torner, C.; Tian, L.; Hu, J.; et al. Structural basis for distinct roles of SMAD2 and SMAD3 in FOXH1 pioneer-directed TGF-β signaling. Genes Dev. 2019, 33, 1506–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besecker, B.; Bao, S.; Bohacova, B.; Papp, A.; Sadee, W.; Knoell, D.L. The human zinc transporter SLC39A8 (Zip8) is critical in zinc-mediated cytoprotection in lung epithelia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L1127–L1136. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.; Paterson, P.J.; Mills, E.A.; McKirdy, A.; Fell, G.S.; Ottoway, J.M.; Husain, F.E.R.; Fitzgerald-Finch, O.P.; Yates, A.J.; Lamont, A.; et al. Clinical and biochemical abnormalities in coppersmiths exposed to cadmium. Lancet 1976, 308, 396–398. [Google Scholar] [CrossRef]
- Watanabe, T.; Zhang, Z.-W.; Moon, C.-S.; Shimbo, S.; Nakatsuka, H.; Matsuda-Inoguchi, N.; Higashikawa, K.; Ikeda, M. Cadmium exposure of women in general populations in Japan during 1991–1997 compared with 1977–1981. Int. Arch. Occup. Environ. Health 2000, 73, 26–34. [Google Scholar] [CrossRef]
- Ikeda, M.; Moon, C.-S.; Zhang, Z.-W.; Iguchi, H.; Watanabe, T.; Iwami, O.; Imai, Y.; Shimbo, S. Urinary α1-microglobulin, β2-microglobulin, and retinol-binding protein levels in general populations in Japan with references to cadmium in urine, blood, and 24-hour food duplicates. Environ. Res. 1995, 70, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Kissane, J.M.; Robins, E. The fluorometric measurement of deoxyribonucleic acid in animal tissues with special reference to the central nervous system. J. Biol. Chem. 1958, 233, 184–188. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (3′→5′) |
|---|---|---|
| ZIP1 | TTCTCTACATCACCTTCCTGG | AACCTTCCTTGCCTGTCTTG |
| ZIP2 | GCTCTCGCTCTCCTTTCAC | ACCAGCCGCAGTCCTACA |
| ZIP3 | GGACACACTCACCTCAACGC | CTCAAGGCTCCAAGCAGAAC |
| ZIP4 | GACAGCCACAGTGACGACAG | CAGACATTCCGTACACAGCC |
| ZIP6 | CCTGAAAATGATGATGATGTGG | CAAGATTGCTGGCTGCTGAG |
| ZIP7 | TATTCTATGTAGCAACGGTGTC | CGAGGTGGCAATCAAC |
| ZIP8 | GAATGAGCACTCGACAAGCC | TAGAGGAACATGCCTCCAGC |
| ZIP9 | CAGCCTCTTGTCTCGCCTTG | ATGTCTGTATCCTTCGCAGTGTG |
| ZIP10 | TTCTATCACTGTCATTAGCCTGC | GCGTCTCCACTCATTGTTCC |
| ZIP11 | CCATCACCATCCACAACATC | TACCAGAAGGCTCTCCAGG |
| ZIP12 | GAGGACAGATGGAAGGCAAG | TGATGTAATAGAGGAGGAGAAGAG |
| ZIP13 | CTGGACAGTAAGGAGAGCGAG | GAGCAGGTGGAAGAAACACG |
| ZIP14 | TCTCGGTAGTGCCTCTGTCC | GAATGTCTCAGTGCTGGTTGG |
| ZnT1 | CAACAGCAGCAACTCCAACG | CCAGTCTTATCTTCATCCTCTTCC |
| DMT1 | CACAGGTAGCCATCAGAGCC | ACCAGGTTAGGAGTTCAGGAG |
| ALK1 | CAACCACTACTGCTGCTACA | CCATCTCCTTGAGGCTGC |
| ALK5 | GTCTGCTTTGTCTGTATCTCACTCA | TCCTCTTCATTTGGCACTCG |
| Smad2 | CAGAATACCGAAGGCAGACG | TGAGCAACGCACTGAAGG |
| Smad3 | ACTACAGCCATTCCATCC | ATCTGGTGGTCACTGGTCTC |
| GAPDH | CAATGACCCCTTCATTGACCTTC | GGATCTCGCTCCTGGAAGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, K.; Fujie, T.; Shimomura, M.; Nakano, T.; Yamamoto, C.; Kaji, T. TGF-β1 Potentiates the Cytotoxicity of Cadmium by Induction of a Metal Transporter, ZIP8, Mediated by the ALK5-Smad2/3 and ALK5-Smad3-p38 MAPK Signal Pathways in Cultured Vascular Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 448. https://doi.org/10.3390/ijms23010448
Ito K, Fujie T, Shimomura M, Nakano T, Yamamoto C, Kaji T. TGF-β1 Potentiates the Cytotoxicity of Cadmium by Induction of a Metal Transporter, ZIP8, Mediated by the ALK5-Smad2/3 and ALK5-Smad3-p38 MAPK Signal Pathways in Cultured Vascular Endothelial Cells. International Journal of Molecular Sciences. 2022; 23(1):448. https://doi.org/10.3390/ijms23010448
Chicago/Turabian StyleIto, Keisuke, Tomoya Fujie, Masahiro Shimomura, Tsuyoshi Nakano, Chika Yamamoto, and Toshiyuki Kaji. 2022. "TGF-β1 Potentiates the Cytotoxicity of Cadmium by Induction of a Metal Transporter, ZIP8, Mediated by the ALK5-Smad2/3 and ALK5-Smad3-p38 MAPK Signal Pathways in Cultured Vascular Endothelial Cells" International Journal of Molecular Sciences 23, no. 1: 448. https://doi.org/10.3390/ijms23010448