
A Review on the Role of miR-149-5p in the Carcinogenesis

 , ,
, ,
Abstract
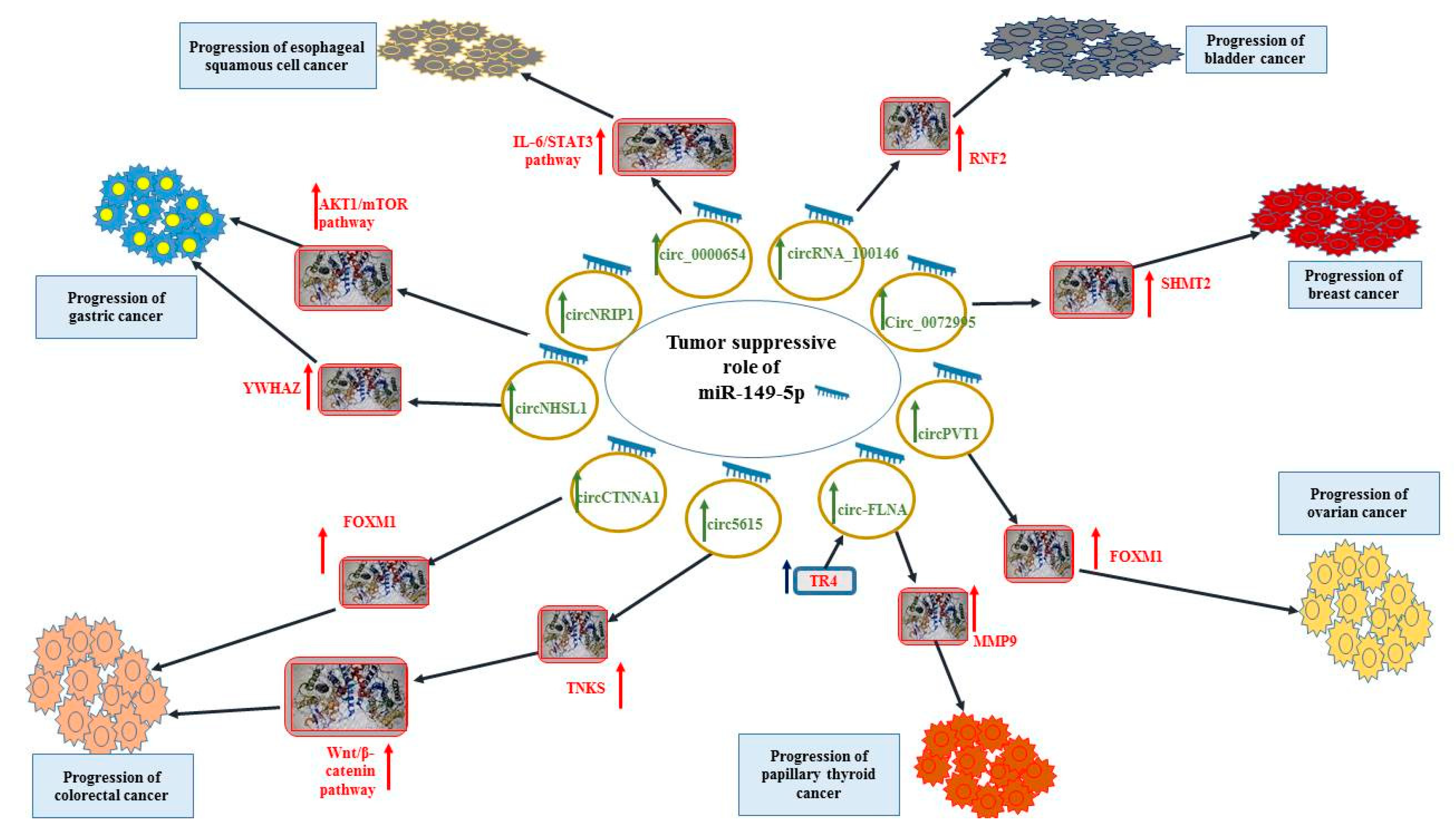
:1. Introduction
2. Cell Line Studies
2.1. Gastrointestinal Tumors
2.2. Thyroid Cancer
2.3. Oral Cancer
2.4. Ovarian Cancer
2.5. Osteosarcoma
2.6. Breast Cancer
2.7. Urogenital Cancers
2.8. Lung Cancer
2.9. Hepatocellular Carcinoma
3. Animal Studies
4. Clinical Studies

{kind=link}
{kind=link}
{kind=link}
| Tumor Type | Samples | Expression of miR-149-5p or Other Genes (Tumor vs. Normal) | Kaplan-Meier Analysis (Impact of miR-149-5p Dysregulation or Other Genes Dysregulation) | Association of Expression of miR-149-5p or Expression of Other Genes with Clinicopathologic Characteristics | Reference |
|---|---|---|---|---|---|
| Gastric cancer (GC) | 3 pairs of GC tissues and ANCTs | Up-regulation of circNRIP1 (which sponges miR-149-5p) | shorter OS and DFS | GC tumor size and lymphatic invasion | [6] |
| 40 GC patients and 40 normal controls | Up-regulation of circNRIP1 (which sponges miR-149-5p) | _ | _ | ||
| 52 pairs of GC tissues and ANCTs | Up-regulation of BLACAT1 (which sponges miR-149-5p) | _ | WHO grade and TNM stage | [7] | |
| 20 GC patients and 20 normal controls | Up-regulation of CircNHSL1 (which sponges miR-149-5p) | _ | tumor size, TNM stages, lymphatic metastasis, and distant metastasis | [8] | |
| GEO databases: (GSE23739, GSE26595, GSE26645, GSE28700, GSE33743, GSE54397, GSE63121, GSE78091, and GSE93415 | Down-regulation of miR-149-5p | _ | distal stomach | [50] | |
| TCGA database | Down-regulation of miR-149-5p | _ | _ | ||
| Stomach Adenocarcinoma (STAD) | serum samples from 130 STAD patients and 116 healthy controls | Down-regulation of miR-149-5p | _ | _ | [51] |
| serum samples from 30 STAD patients and 24 healthy controls | Down-regulation of miR-149-5p | _ | _ | ||
| Colorectal cancer (CRC) | 21 pairs of CRC tissues and ANCTs | Up-regulation of LINC00460 (which sponges miR-149-5p) | _ | clinical stage and node status | [9] |
| TCGA database | Up-regulation of LINC00460 (which sponges miR-149-5p) | _ | _ | ||
| 40 pairs of CRC tissues and ANCTs | Up-regulation of LINC00460 (which sponges miR-149-5p) | Poorer DFS | _ | [10] | |
| 60 pairs of colon cancer tissues and ANCTs | Up-regulation of circCTNNA1 (which sponges miR-149-5p), Up-regulation of FOXM1 | _ | advanced TNM stage | [11] | |
| 180 pairs of CRC tissues and ANCTs | Up-regulation of circCTNNA1 (which sponges miR-149-5p) | lower OS | _ | ||
| 55 pairs of CRC tissues and ANCTs | Down-regulation of miR-149-5p | _ | _ | [12] | |
| 42 pairs of CRC tumor tissues and ANCTs | Up-regulation of DLGAP1-AS1 (which sponges miR-149-5p) | lower OS | advanced clinical stage (phase III–IV) | [13] | |
| GSE142837 analysis | Up-regulation of circ5615 (which sponges miR-149-5p) | _ | _ | [14] | |
| 99 pairs of CRC tissues and ANCTs | Up-regulation of circ5615 (which sponges miR-149-5p) | shorter OS | higher T stage | ||
| TCGA and GEO dataset: GSE33113 and GSE41328 | Up-regulation of LINC00460 (which sponges miR-149-5p) | shorter OS | larger tumor sizes, advanced TNM stages, and lymph node metastasis | [40] | |
| Medullary thyroid carcinoma (MTC) | 36 42 pairs of MTC tumor tissues and ANCTs | Down-regulation of miR-149-5p | shorter OS | distant metastases and TNM stage | [15] |
| Papillary thyroid cancer (PTC) | 20 pairs of primary PTC tumors and the paired lymph node metastatic PTC tumors | Up-regulation of TR4 | shorter OS | nodal metastasis | [16] |
| Oral squamous cell carcinoma (OSCC) | 34 pairs of OSCC tissues and ANCTs | Down-regulation of miR-149-5p | _ | _ | [17] |
| 10 pairs of OSCC tissues and ANCTs | Up-regulation of DLEU1 (which sponges miR-149-5p) | _ | advanced stage | [18] | |
| Ovarian cancer | TCGA datasets | Up-regulation of miR-149-5p in chemoresistant tissues | _ | _ | [19] |
| 20 ovarian cancer tissues and 10 benign ovarian lesion tissues | Up-regulation of miR-149-5p in ovarian cancer tissues than benign ovarian tissues | _ | _ | ||
| GTEx database | Up-regulation of circPVT1 | shorter PFS | stage 3/4 and grade 3 OV | [20] | |
| Osteosarcoma | 191 sarcoma patients and 66 adjacent normal samples | Down-regulation of miR-149-5p | shorter OS | age and tumor size | [21] |
| Breast cancer | 70 pairs of breast cancer tissues and ANCTs | Up-regulation of circ_0072995 | _ | _ | [22] |
| 1104 breast cancer tissues and 113 adjacent normal tissues | Up-regulation of SHMT2 | _ | _ | ||
| Lung cancer | TCGA dataset | Up-regulation of B3GNT3 (a target of miR-149-5p) | lower OS and DFS | advanced TNM stages | [27] |
| 120 pairs of lung cancer tissues and ANCTs | Up-regulation of B3GNT3 (a target of miR-149-5p) | lower OS and DFS | late TNM stages, bigger tumor size, distant metastasis and recurrence | ||
| GEO dataset: GSE19188 (91 NSCLC tissues and 65 ANCTs | Up-regulation of HOTAIR (which sponges miR-149-5p) | unfavorable DFS | _ | [28] | |
| 60 pairs of NSCLC tissues and ANCTs | Up-regulation of HNF1A-AS1 (which sponges miR-149-5p) | shorter OS | advanced TMN stage, big tumor size, and lymph node metastasis | [29] | |
| 80 pairs of NSCLC and ANCTs | Up-regulation of MIAT (which sponges miR-149-5p) | shorter OS | advanced pathological stage | [30] | |
| 35 DDP-resistant NSCLC tumors and 35 DDP-sensitive NSCLC tumors | Up-regulation of HOTAIR in DDP-resistant NSCLC tumors than DDP-sensitive NSCLC tumors | _ | TNM stage, lymph node metastasis and DDP response | [31] | |
| GEO dataset: GSE111803 analysis (5 lung adenocarcinoma patients and 5 healthy controls) | Up-regulation of miR-149-5p | _ | advanced clinical stage | [34] | |
| TCGA dataset | Up-regulation of LINC00460 in lung adenocarcinoma tissues with EGFR-activating mutations than in normal tissues | _ | _ | [32] | |
| 62 patients with EGFR-mutant lung adenocarcinoma treated with EGFR-TKI | Up-regulation of LINC00460 | shorter OS and PFS | _ | ||
| Hepatocellular carcinoma (HCC) | 79 pairs of HCC tissues and ANCTs | Up-regulation of NEAT1 (which sponges miR-149-5p) Down-regulation of miR-149-5p | shorter OS and DFS | tumor stage, lymphatic metastasis, and sorafenib resistance | [36] |
| 48 pairs of HCC tissues and ANCTs | Up-regulation of PART1 (which sponges miR-149-5p) | _ | _ | [37] | |
| 23 pairs of HCC tissues and ANCTs | Up-regulation of SNHG8 (which sponges miR-149-5p) | higher recurrence rates | _ | [41] | |
| Prostate carcinoma (PCa) | GEO DataSets: GSE17317 and GSE34932 | Down-regulation of miR-149-5p | _ | _ | [25] |
| 30 pairs of PCa tissues and ANCTs | Down-regulation of miR-149-5p | _ | _ | ||
| Esophageal squamous cell cancer (ESCC) | 55 pairs of ESCC tissues and ANCTs | Up-regulation of circ_0000654 (which sponges miR-149-5p) | _ | higher T stage and local lymph node metastasis | [39] |
| Bladder cancer | 68 pairs of Bladder cancer tissues and ANCTs | Up-regulation of circRNA_100146 (which sponges miR-149-5p) Down-regulation of miR-149-5p | _ | tumor stage, LN status, histological grade, and tumor size | [26] |
| Bladder cancer | 10 Bladder cancer patients and 10 healthy controls | Up-regulation of miR-149-5p | _ | _ | [49] |
| TCGA database | Up-regulation of miR-149-5p | poor OS | _ | ||
| urine of 70 Bladder cancer patients and 90 healthy controls | Up-regulation of miR-149-5p | _ | _ | ||
| Renal cell carcinoma (RCC) | 32 pairs of RCC tissues and ANCTs | Down-regulation of miR-149-5p | _ | _ | [42] |
| TCGA database (237 pairs of ccRCC tissues and ANCTs) | Down-regulation of miR-149-5p | shorter OS | _ | [52] | |
| 16 pairs of ccRCC tissues and ANCTs | Down-regulation of miR-149-5p | _ | _ | [43] | |
| Cervical cancer (CC) | 37 pairs of CC tissues and ANCTs | Up-regulation of circ_0075341 (which sponges miR-149-5p) | _ | larger tumor size, advanced FIGO stage, and lymph-node metastasis | [44] |
| GEO database: GSE102686 | Up-regulation of circ_0011385 (which sponges miR-149-5p) | _ | _ | [45] | |
| 50 pairs of CC tissues and ANCTs | Up-regulation of circ_0011385 (which sponges miR-149-5p) | _ | higher FIGO stage | ||
| Melanoma | _ melanoma tissue and ANCTs | Up-regulation of LRIG2 (a target of miR-149-5p) | _ | _ | [46] |
| Acute myeloid leukemia (AML) | 45 AML, T cell ALL, and CML patients and 20 healthy controls | Up-regulation of miR-149-5p | _ | _ | [47] |
| Acute lymphocytic leukemia (ALL) | GSE166579 | Dow-regulation of circADD2 (which sponges miR-149-5p) | _ | _ | [48] |
| 30 ALL patients and 30 controls | Dow-regulation of circADD2 (which sponges miR-149-5p) | _ | _ | ||
| Pancreatic ductal adenocarcinoma (PDAC) | 27 PDAC patients and 3 healthy controls | Up-regulation of miR-149-5p | _ | _ | [53] |
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Macfarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussen, B.M.; Hidayat, H.J.; Salihi, A.; Sabir, D.K.; Taheri, M.; Ghafouri-Fard, S. MicroRNA: A signature for cancer progression. Biomed. Pharmacother. 2021, 138, 111528. [Google Scholar] [CrossRef]
- He, Y.; Yu, D.; Zhu, L.; Zhong, S.; Zhao, J.; Tang, J. miR-149 in Human Cancer: A Systemic Review. J. Cancer 2018, 9, 375–388. [Google Scholar] [CrossRef]
- Khan, R.; Raza, S.H.A.; Junjvlieke, Z.; Wang, X.; Wang, H.; Cheng, G.; Mei, C.; Elnour, I.E.; Zan, L. Bta-miR-149-5p inhibits proliferation and differentiation of bovine adipocytes through targeting CRTCs at both transcriptional and posttranscriptional levels. J. Cell. Physiol. 2020, 235, 5796–5810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Dong, Y.; Liu, M.; Yang, L.; Zhao, Z. miR-149-5p Inhibits Vascular Smooth Muscle Cells Proliferation, Invasion, and Migration by Targeting Histone Deacetylase 4 (HDAC4). Med. Sci. Monit. 2019, 25, 7581–7590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, S.; Wang, H.; Cao, J.; Huang, X.; Chen, Z.; Xu, P.; Sun, G.; Xu, J.; Lv, J.; et al. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol. Cancer 2019, 18, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, X.; Liu, X.; Niu, D. Long Non-Coding RNA BLACAT1 Promotes the Tumorigenesis of Gastric Cancer by Sponging microRNA-149-5p and Targeting KIF2A. Cancer Manag. Res. 2020, 12, 6629–6640. [Google Scholar] [CrossRef]
- Hui, C.; Tian, L.; He, X. Circular RNA circNHSL1 Contributes to Gastric Cancer Progression Through the miR-149-5p/YWHAZ Axis. Cancer Manag. Res. 2020, 12, 7117–7130. [Google Scholar] [CrossRef]
- Meng, X.; Sun, W.; Yu, J.; Zhou, Y.; Gu, Y.; Han, J.; Zhou, L.; Jiang, X.; Wang, C. LINC00460-miR-149-5p/miR-150-5p-Mutant p53 Feedback Loop Promotes Oxaliplatin Resistance in Colorectal Cancer. Mol. Ther. Nucleic Acids 2020, 22, 1004–1015. [Google Scholar] [CrossRef]
- Ruan, T.; Lu, S.; Xu, J.; Zhou, J.-Y. lncRNA LINC00460 Functions as a Competing Endogenous RNA and Regulates Expression of BGN by Sponging miR-149-5p in Colorectal Cancer. Technol. Cancer Res. Treat. 2021, 20, 1533033820964238. [Google Scholar] [CrossRef]
- Chen, P.; Yao, Y.; Yang, N.; Gong, L.; Kong, Y.; Wu, A. Circular RNA circCTNNA1 promotes colorectal cancer progression by sponging miR-149-5p and regulating FOXM1 expression. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Wang, A.-H.; Fan, W.-J.; Fu, L.; Wang, X.-T. LncRNA PCAT-1 regulated cell proliferation, invasion, migration and apoptosis in colorectal cancer through targeting miR-149-5p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8310–8320. [Google Scholar] [PubMed]
- Qu, L.; Chen, Y.; Zhang, F.; He, L. The lncRNA DLGAP1-AS1/miR-149-5p/TGFB2 axis contributes to colorectal cancer progression and 5-FU resistance by regulating smad2 pathway. Mol. Ther. Oncol. 2021, 20, 607–624. [Google Scholar] [CrossRef]
- Ma, Z.; Han, C.; Xia, W.; Wang, S.; Li, X.; Fang, P.; Yin, R.; Xu, L.; Yang, L. circ5615 functions as a ceRNA to promote colorectal cancer progression by upregulating TNKS. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Chen, X. miR-149-5p inhibits cell proliferation and invasion through targeting GIT1 in medullary thyroid carcinoma. Oncol. Lett. 2018, 17, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, X.; Feng, L.; Yao, L.; Xiao, Y.; Hu, X.; Zhang, G.; Liu, G.; Wang, Z. Testicular orphan receptor 4 (TR4) promotes papillary thyroid cancer invasion via activating circ-FNLA/miR-149-5p/MMP9 signaling. Mol. Ther. Nucleic Acids 2021, 24, 755–767. [Google Scholar] [CrossRef]
- Luo, K.; He, J.; Yu, D.; Açil, Y. MiR-149-5p regulates cisplatin chemosensitivity, cell growth, and metastasis of oral squamous cell carcinoma cells by targeting TGFβ2. Int. J. Clin. Exp. Pathol. 2019, 12, 3728–3739. [Google Scholar]
- Lv, T.; Liu, H.; Wu, Y.; Huang, W. Knockdown of lncRNA DLEU1 inhibits the tumorigenesis of oral squamous cell carcinoma via regulation of miR-149-5p/CDK6 axis. Mol. Med. Rep. 2021, 23, 1–11. [Google Scholar] [CrossRef]
- Xu, M.; Xiao, J.; Chen, M.; Yuan, L.; Li, J.; Shen, H.; Yao, S. miR-149-5p promotes chemotherapeutic resistance in ovarian cancer via the inactivation of the Hippo signaling pathway. Int. J. Oncol. 2018, 52, 815–827. [Google Scholar] [CrossRef]
- Li, M.; Chi, C.; Zhou, L.; Chen, Y.; Tang, X. Circular PVT1 regulates cell proliferation and invasion via miR-149-5p/FOXM1 axis in ovarian cancer. J. Cancer 2021, 12, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.D.; Feng, F.; Yu, X.-S.; Liu, Z.-D.; Lao, L.-F. miR-149-5p inhibits cell growth by regulating TWEAK/Fn14/PI3K/AKT pathway and predicts favorable survival in human osteosarcoma. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418786656. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Qin, X.; Zhou, Z.; Wang, Y.; Yang, Q.; Liao, T. Circ_0072995 promotes cell carcinogenesis via up-regulating miR-149-5p-mediated SHMT2 in breast cancer. Cancer Manag. Res. 2020, 12, 11169–11181. [Google Scholar] [CrossRef]
- Tian, D.; Tian, M.; Ma, Z.-m.; Zhang, L.-l.; Cui, Y.-f.; Li, J.-l. Anesthetic propofol epigenetically regulates breast cancer trastuzumab resistance through IL-6/miR-149-5p axis. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xiang, F.; Fan, Y.; Ni, Z.; Liu, Q.; Zhu, Z.; Chen, Z.; Hao, W.; Yue, H.; Wu, R.; Kang, X. Ursolic Acid Reverses the Chemoresistance of Breast Cancer Cells to Paclitaxel by Targeting MiRNA-149-5p/MyD88. Front. Oncol. 2019, 9, 501. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wei, H.; Li, X.; Qu, X. Hsa-miR-149-5p Suppresses Prostate Carcinoma Malignancy by Suppressing RGS17. Cancer Manag. Res. 2021, 13, 2773–2783. [Google Scholar] [CrossRef]
- Wang, H.; Niu, X.; Mao, F.; Liu, X.; Zhong, B.; Jiang, H.; Fu, G. Hsa_circRNA_100146 Acts as a Sponge of miR-149-5p in Promoting Bladder Cancer Progression via Regulating RNF2. OncoTargets Ther. 2020, 13, 11007–11017. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, T.; Xian, L.; Liu, W.; Liu, J.; Zhou, H. B3GNT3, a Direct Target of miR-149-5p, Promotes Lung Cancer Development and Indicates Poor Prognosis of Lung Cancer. Cancer Manag. Res. 2020, 12, 2381–2391. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cui, Z.; Lv, X.; Li, J.; Gao, M.; Yang, Z.; Bi, Y.; Zhang, Z.; Wang, S.; Li, S.; et al. Long Non-coding RNA HOTAIR Function as a Competing Endogenous RNA for miR-149-5p to Promote the Cell Growth, Migration, and Invasion in Non-small Cell Lung Cancer. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Y.; Li, Q.; Duan, P. lncRNA HNF1A-AS1 modulates non–small cell lung cancer progression by targeting miR-149-5p/Cdk6. J. Cell. Biochem. 2019, 120, 18736–18750. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, S.; Xiong, Y. Long noncoding RNA MIAT promotes non-small cell lung cancer progression by sponging miR-149-5p and regulating FOXM1 expression. Cancer Cell Int. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Zhan, Y.; Abuduwaili, K.; Wang, X.; Shen, Y.; Nuerlan, S.; Liu, C. Knockdown of Long Non-Coding RNA HOTAIR Suppresses Cisplatin Resistance, Cell Proliferation, Migration and Invasion of DDP-Resistant NSCLC Cells by Targeting miR-149-5p/Doublecortin-Like Kinase 1 Axis. Cancer Manag. Res. 2020, 12, 7725–7737. [Google Scholar] [CrossRef]
- Nakano, Y.; Isobe, K.; Kobayashi, H.; Kaburaki, K.; Isshiki, T.; Sakamoto, S.; Takai, Y.; Tochigi, N.; Mikami, T.; Iyoda, A.; et al. Clinical importance of long non-coding RNA LINC00460 expression in EGFR-mutant lung adenocarcinoma. Int. J. Oncol. 2019, 56, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Qin, X.; Yan, D.; Cao, H.; Zhou, L.; Fan, F.; Zang, J.; Ni, J.; Xu, X.; Sha, H.; et al. Genome-wide profiling of micro-RNA expression in gefitinib-resistant human lung adenocarcinoma using microarray for the identification of miR-149-5p modulation. Tumor Biol. 2017, 39, 1010428317691659. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Yang, H.; Zhou, B. Integrative analysis of exosomal microRNA-149-5p in lung adenocarcinoma. Aging 2021, 13, 7382–7396. [Google Scholar] [CrossRef]
- Liu, G.; Yin, L.; Ouyang, X.; Zeng, K.; Xiao, Y.; Li, Y. M2 Macrophages Promote HCC Cells Invasion and Migration via miR-149-5p/MMP9 Signaling. J. Cancer 2020, 11, 1277–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Niu, Y.; Tang, G.; Wu, C. LncRNA NEAT1 modulates sorafenib resistance in hepatocellular carcinoma through regulating the miR-149-5p/AKT1 axis. Saudi J. Gastroenterol. 2020, 26, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Wang, P.; Tu, M.; Huang, Y.; Xiong, F.; Wu, Y. Long Non-Coding RNA PART1 Promotes Proliferation, Migration and Invasion of Hepatocellular Carcinoma Cells via miR-149-5p/MAP2K1 Axis. Cancer Manag. Res. 2020, 12, 3771–3782. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ni, J.; Liu, Y.-X.; Wang, H.; Liang, Z.-Q.; Wang, X. Response of MiRNA-22-3p and MiRNA-149-5p to Folate Deficiency and the Differential Regulation of MTHFR Expression in Normal and Cancerous Human Hepatocytes. PLoS ONE 2017, 12, e0168049. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Tie, X.; Li, N.; Yi, Z.; Shen, F.; Zhang, Y. Circular RNA hsa_circ_0000654 promotes esophageal squamous cell carcinoma progression by regulating the miR-149-5p/IL-6/STAT3 pathway. IUBMB Life 2020, 72, 426–439. [Google Scholar] [CrossRef]
- Lian, Y.; Yan, C.; Xu, H.; Yang, J.; Yu, Y.; Zhou, J.; Shi, Y.; Ren, J.; Ji, G.; Wang, K. A Novel lncRNA, LINC00460, Affects Cell Proliferation and Apoptosis by Regulating KLF2 and CUL4A Expression in Colorectal Cancer. Mol. Ther. Nucleic Acids 2018, 12, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Teng, F.; Guo, W.; Yang, J.; Ding, G.; Fu, Z. lncRNA SNHG8 Promotes the Tumorigenesis and Metastasis by Sponging miR-149-5p and Predicts Tumor Recurrence in Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2018, 51, 2262–2274. [Google Scholar] [CrossRef]
- Jin, L.; Li, Y.; Liu, J.; Yang, S.; Gui, Y.; Mao, X.; Nie, G.; Lai, Y. Tumor suppressor miR-149-5p is associated with cellular migration, proliferation and apoptosis in renal cell carcinoma. Mol. Med. Rep. 2016, 13, 5386–5392. [Google Scholar] [CrossRef] [Green Version]
- Okato, A.; Arai, T.; Yamada, Y.; Sugawara, S.; Koshizuka, K.; Fujimura, L.; Kurozumi, A.; Kato, M.; Kojima, S.; Naya, Y. Dual strands of pre-miR-149 inhibit cancer cell migration and invasion through targeting FOXM1 in renal cell carcinoma. Int. J. Mol. Sci. 2017, 18, 1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; Wang, C.; Wang, S.; Zhang, H.; Zhang, Y. Hsa_circ_0075341 is up-regulated and exerts oncogenic properties by sponging miR-149-5p in cervical cancer. Biomed. Pharmacother. 2020, 121, 109582. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.L.; Wang, W.-S.; Zhao, M.-Y.; Sun, J.-N.; Chen, X.-R.; Hou, J.-Q. Circular RNA circ_0011385 promotes cervical cancer progression through competitively binding to miR-149-5p and up-regulating SOX4 expression. Kaohsiung J. Med. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, J.; Xu, H.; Dai, J.; Zhang, X. The negative regulation of miR-149-5p in melanoma cell survival and apoptosis by targeting LRIG2. Am. J. Transl. Res. 2017, 9, 4331–4340. [Google Scholar]
- Tian, P.; Yan, L. Inhibition of MicroRNA-149-5p Induces Apoptosis of Acute Myeloid Leukemia Cell Line THP-1 by Targeting Fas Ligand (FASLG). Med. Sci. Monit. 2016, 22, 5116–5123. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ma, X.; Zhang, H.; Wu, Y.; Kang, M.; Fang, Y.; Xue, Y. Mechanism of circADD2 as ceRNA in Childhood Acute Lymphoblastic Leukemia. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Lin, J.-T.; Tsai, K.-W. Circulating miRNAs Act as Diagnostic Biomarkers for Bladder Cancer in Urine. Int. J. Mol. Sci. 2021, 22, 4278. [Google Scholar] [CrossRef]
- Liu, X.; Pu, K.; Wang, Y.; Chen, Y.; Zhou, Y. Gastric cancer-associated microRNA expression signatures: Integrated bioinformatics analysis, validation, and clinical significance. Ann. Transl. Med. 2021, 9, 797. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Peng, X.; Zhang, C.; Liu, K.; Huang, G.; Lai, Y. Use of a Four-miRNA Panel as a Biomarker for the Diagnosis of Stomach Adenocarcinoma. Dis. Markers 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Lv, Y.; Liu, Z.; Zhang, J.; Liang, C.; Liao, X.; Liang, R.; Lin, Y.; Li, Y. Identification and validation of a four-miRNA(miRNA-21-5p, miRNA-9-5p, miR-149-5p, and miRNA-30b-5p) prognosis signature in clear cell renal cell carcinoma. Cancer Manag. Res. 2018, 10, 5759–5766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, J.; Cui, W.; Liu, Y.; Zhou, H.; Wang, Y.; Chen, X.; Chen, X.; Wang, Z. Serum Exosomal miRNA-1226 as Potential Biomarker of Pancreatic Ductal Adenocarcinoma. OncoTargets Ther. 2021, 14, 1441–1451. [Google Scholar] [CrossRef] [PubMed]



| Tumor Type | Targets/Regulators and Signaling Pathways that Interact with miR-149-5p | Function | Reference |
|---|---|---|---|
| Gastric cancer | circNRIP1, AKT1/mTOR pathway | ∆ circNRIP1: 44001proliferation, ↓ invasion, ↓ migration | [6] |
| BLACAT1, KIF2A | ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration | [7] | |
| circNHSL1, YWHAZ | ∆ circNRIP1: ↓ invasion, ↓ migration, ↓ glutaminolysis | [8] | |
| Colorectal cancer | LINC00460, p53, miR-150-5p | ∆ p53 or LINC00460: ↓ oxaliplatin resistance | [9] |
| LINC00460, BGN | ∆ LINC00460: ↓ invasion, ↓ migration | [10] | |
| circCTNNA1, FOXM1 | ↑ circCTNNA1: ↑ proliferation, ↑ invasion, ↑ migration, ↑ viability, ↑ colony-forming, ↑ DNA synthesis ∆ circCTNNA1: ↓ proliferation, ↓ viability, ↓ colony-forming, ↑ G1 arrest | [11] | |
| PCAT-1 | ∆ PCAT-1: ↓ proliferation, ↓ invasion, ↓ migration, ↑ apoptosis | [12] | |
| DLGAP1-AS1, TGFB2 signaling pathway | ∆ DLGAP1-AS1: ↓ proliferation, ↓ invasion, ↓ migration, ↑ apoptosis, ↑ sensitivity to 5-FU ∆ miR-149-5p: ↑ proliferation, ↓ apoptosis | [13] | |
| circ5615, TNKS, Wnt/β-catenin pathway | ∆ DLGAP1-AS1: ↓ proliferation, ↓ invasion ↑ DLGAP1-AS1: ↑ proliferation, ↑ transition from G1/S to G2/M phase | [14] | |
| LINC00460, CUL4A | ∆ DLGAP1-AS1: ↓ proliferation, ↑ apoptosis, ↑ G0/G1 phase arrest | [40] | |
| Medullary thyroid carcinoma | GIT1 | ∆ miR-149-5p: ↑ proliferation, ↑ invasion ↑ miR-149-5p: ↓ proliferation, ↓ invasion | [15] |
| Papillary thyroid cancer | circ-FLNA, TR4, MMP9 | ∆ TR4: ↓ proliferation, ↓ invasion, ↓ migration | [16] |
| Oral squamous cell carcinoma | TGFβ2 | ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration, ↓ CDDP resistance, ↑ apoptosis | [17] |
| DLEU1, CDK6 | ∆ DLEU1: ↓ proliferation, ↑ G1 cell cycle arrest | [18] | |
| Ovarian cancer | Hippo signaling pathway, MST1, SAV1 | ↑ miR-149-5p: ↓ apoptosis, ↑ resistance to CDDP ∆ miR-149-5p: ↑ apoptosis, ↓ resistance to CDDP | [19] |
| circPVT1, FOXM1 | ∆ circPVT1: ↓ viability, ↓ invasion, ↓ migration | [20] | |
| Osteosarcoma | TNFRSF12A, TWEAK/EGFR pathway | ∆ miR-149-5p: ↑ proliferation, ↑ colony formation | [21] |
| Breast cancer | Circ_0072995, SHMT2 | ∆ Circ_0072995: ↓ proliferation, ↓ invasion, ↓ migration, ↓ anaerobic glycolysis↑ apoptosis ∆ SHMT2: ↓ proliferation, ↓ invasion, ↓ migration, ↓ anaerobic glycolysis↑ apoptosis | [22] |
| IL-6 | ∆ miR-149-5p: ↓ propofol effects (↓ sensitization to trastuzumab) | [23] | |
| MyD88 | UA treatment: ↑ miR-149-5p = ↓ PTX resistance | [24] | |
| Lung cancer | B3GNT3 | ∆ B3GNT3: ↓ proliferation, ↓ invasion, ↓ colony formation ↑ miR-149-5p: ↓ proliferation, ↓ tumorigenesis | [27] |
| HOTAIR, HNRNPA1 | ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration, ↑ G0/G1 phase arrest, did not affect apoptosis | [28] | |
| HNF1A-AS1, Cdk6 | ∆ HNF1A-AS1: ↓ proliferation, ↓ invasion, ↓ migration | [29] | |
| MIAT, FOXM1 | ∆ MIAT: ↓ proliferation, ↓ invasion, ↓ migration, ↓ colony formation, ↑ G1 phase arrest | [30] | |
| _ | ∆ miR-149-5p: ↓ gefitinib resistance, ↑ apoptosis ↑ miR-149-5p: ↑ proliferation, ↑ gefitinib resistance | [33] | |
| HOTAIR, DCLK1 | ∆ HOTAIR: ↓ proliferation, ↓ invasion, ↓ migration, ↓ DDP resistance | [31] | |
| AMTOL2 | ↑ miR-149-5p: ↑ proliferation, ↓ apoptosis ∆ miR-149-5p: ↓ proliferation, ↑ apoptosis | [34] | |
| LINC00460, IL-6 | ∆ LINC00460: ↓ EMT process, ↓ migration, ↓ viability following treatment with various EGFR-TKIs, ↑ apoptosis following treatment with various EGFR-TKIs | [32] | |
| Hepatocellular carcinoma | MMP9 | co-culture: PMA + IL-4 and IL-13: ↑ invasion, ↑ migration | [35] |
| NEAT1, AKT1 | ∆ NEAT1: ↑ sensitivity to sorafenib, ↑ apoptosis, ↓ viability | [36] | |
| PART1, MAP2K1 | ∆ PART1: ↓ proliferation, ↓ invasion, ↓ migration ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration | [37] | |
| SNHG8 | ∆ SNHG8: ↓ proliferation, ↓ invasion, ↓ migration, ↓ viability, ↓ colony formation | [41] | |
| MTHFR, TP53INP1, PDCD4 | FA deficiency: ↑ miR-149-5p | [38] | |
| Prostate carcinoma | RGS17 | ↑ miR-149-5p: ↓ proliferation, ↓ migration, ↓ viability | [25] |
| Esophageal squamous cell cancer | circ_0000654, IL-6/STAT3 signaling pathway | ∆ circ_0000654: ↓ proliferation, ↓ invasion, ↓ migration, ↑ apoptosis ↑ circ_0000654: ↑ proliferation, ↑ invasion, ↑ migration, ↓ apoptosis ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration, ↑ apoptosis | [39] |
| Bladder cancer | circRNA_100146, RNF2 | ∆ circRNA_100146: ↓ proliferation, ↓ invasion, ↓ migration, ↑ apoptosis | [26] |
| Renal cell carcinoma | _ | ↑ miR-149-5p: ↓ proliferation, ↓ migration, ↑ apoptosis | [42] |
| FOXM1 | ↑ miR-149-5p: ↓ proliferation, ↓ invasion, ↓ migration | [43] | |
| Cervical cancer | circ_0075341, AURKA | ∆ circ_0075341: ↓ proliferation, ↓ invasion | [44] |
| circ_0011385, SOX4 | ∆ circ_0011385: ↓ multiplication, ↓ invasion, ↓ migration | [45] | |
| Melanoma | LRIG2 | ↑ miR-149-5p: ↓ colony formation, ↑ apoptosis | [46] |
| Acute myeloid leukemia | FASLG, p-FADD, caspases | ∆ miR-149-5p: ↑ apoptosis | [47] |
| Acute lymphocytic leukemia | circADD2, AKT2 | ↑ circADD2: ↓ proliferation, ↓ miR-149-5p | [48] |
| Tumor Type | Animal Models | Results | Reference |
|---|---|---|---|
| Gastric cancer | BALB/c nude mice | ∆ circNRIP1 (which sponges miR-149-5p): ↓ tumor volume, ↓ tumor weight | [6] |
| female BALB/c nude mice | ∆ BLACAT1 (which sponges miR-149-5p): ↓ tumor volume, ↓ tumor weight | [7] | |
| male BALB/C nude mice | ∆ circNHSL1 (which sponges miR-149-5p): ↓ tumor volume, ↓ tumor weight | [8] | |
| Colorectal cancer | BALB/c nude mice | ∆ LINC00460 (which sponges miR-149-5p): ↓ tumor volume, ↓ tumor weight | [9] |
| female BALB/c nude mice | ↑ circCTNNA1 (which sponges miR-149-5p): ↑tumor volume, ↑tumor weight | [11] | |
| female nude mice | ∆ DLGAP1-AS1 (which sponges miR-149-5p): ↓tumor growth, ↑5-FU sensitivity | [13] | |
| male BALB/C nude mice | ∆ circ5615 (which sponges miR-149-5p): ↓ tumor growth | [14] | |
| athymic male mice | ∆ LINC00460 (which sponges miR-149-5p): ↓tumor volume, ↓tumor weight | [40] | |
| Papillary thyroid cancer | male nude mice | ∆ circ-FLNA (which sponges miR-149-5p): ↓tumor growth, ↓tumor volume, ↓ metastasis | [16] |
| Ovarian cancer | BALB/c-nu mice | ∆ miR-149-5p: ↓ tumor volume, ↓ tumor weight, ↓ resistance to CDDP | [19] |
| Breast cancer | female BALB/c nude mice | ∆ circ_0072995: ↓ tumor volume, ↓ tumor weight | [22] |
| female athymic mice | ∆ miR-149-5p: ↓ ability of propofol to inhibit lung metastasis | [23] | |
| female athymic nude mice | UA and PTX treatment: ↓tumor volume, ↓ tumor weight | [24] | |
| Lung cancer | BALB/c nude mice | ∆ B3GNT3: ↓ tumor volume, ↓ tumor weight | [27] |
| female BALB/c nude mice | ∆ HNF1A- AS1: ↓ tumor volume, ↓ tumor weight | [29] | |
| female BALB/c nude mice | ∆ MIAT: ↓ tumor volume, ↓ tumor weight | [30] | |
| athymic BALB/c mice | ∆ HOTAIR: ↓ tumor volume, ↓ tumor weight, ↓ DDP resistance | [31] | |
| Hepatocellular carcinoma | nude mice | ↑ macrophages (THP-1): ↑ metastasis miR-149-5p: ↓ metastasis | [35] |
| nude mice | ∆ SNHG8: ↓ tumor volume, ↓ tumor weight, ↓ metastasis | [41] | |
| Esophageal squamous cell cancer | male BALB/c athymic nude mice | ∆ circ_0000654: ↓ tumor volume, ↓ tumor weight, ↓ metastasis | [39] |
| Acute lymphocytic leukemia | BALB/c nude mice | ↑ circADD2: ↓ tumor volume, ↓ tumor weight | [48] |
| Rget Gene | Representative Transcript | Gene Name | Number of 3P-seq Tags Supporting UTR + 5 | Conserved Sites | Poorly Conserved Sites | 6mer Sites | Cumulative Weighted Context++ Score | Total Context++ Score | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | 8mer | 7mer-m8 | 7mer-A1 | Total | 8mer | 7mer-m8 | 7mer-A1 | |||||||
| LRIG2 | ENST00000361127.5 | leucine-rich repeats and immunoglobulin-like domains 2 | 348 | 1 | 1 | 0 | 0 | 3 | 1 | 2 | 0 | 1 | −0.96 | −1.08 |
| STRADB | ENST00000392249.2 | STE20-related kinase adaptor beta | 12 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | −0.87 | −0.87 |
| ZBTB37 | ENST00000367701.5 | zinc finger and BTB domain containing 37 | 213 | 0 | 0 | 0 | 0 | 5 | 0 | 1 | 4 | 4 | −0.83 | −0.92 |
| TTLL9 | ENST00000375921.2 | tubulin tyrosine ligase-like family, member 9 | 5 | 0 | 0 | 0 | 0 | 3 | 2 | 0 | 1 | 1 | −0.82 | −0.82 |
| FRMD7 | ENST00000370879.1 | FERM domain containing 7 | 5 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 0 | −0.81 | −0.81 |
| TSPAN31 | ENST00000547992.1 | tetraspanin 31 | 72 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 2 | −0.78 | −1.10 |
| CTRC | ENST00000375943.2 | chymotrypsin C (caldecrin) | 5 | 0 | 0 | 0 | 0 | 3 | 1 | 1 | 1 | 2 | −0.77 | −0.77 |
| STRA6 | ENST00000395105.4 | stimulated by retinoic acid 6 | 42 | 0 | 0 | 0 | 0 | 4 | 1 | 3 | 0 | 1 | −0.77 | −0.77 |
| GUCD1 | ENST00000447813.2 | guanylyl cyclase domain containing 1 | 2990 | 0 | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 1 | −0.77 | −0.77 |
| KCNJ5 | ENST00000529694.1 | potassium inwardly-rectifying channel, subfamily J, member 5 | 12 | 0 | 0 | 0 | 0 | 4 | 0 | 4 | 0 | 2 | −0.75 | |
| KEGG ID | KEGG Description | Genes | p Value |
|---|---|---|---|
| ko04962 | Vasopressin-regulated water reabsorption | VAMP2, RAB5C, ADCY9, DCTN2, DCTN4, DCTN5 | 2.45 × 10−52 |
| ko04380 | Osteoclast differentiation | ITGB3, SOCS1, SOCS3, FOS, IFNG, JUN, GAB2, CSF1, IFNAR2, SYK, GRB2, IFNGR2, MITF, IL1A, IL1B, TGFB2 | 1.47 × 10−118 |
| ko05218 | Melanoma | PDGFRB, PDGFRA, TP53, RB1, CDK6, MITF, FGFR1 | 4.09× 10−58 |
| ko05210 | Colorectal cancer | TCF7, TP53, AXIN1, FOS, BIRC5, JUN, TGFB2, MSH6, LEF1 | 1.58× 10−70 |
| ko05332 | Graft-versus-host disease | IFNG, IL6, IL1B, PRF1, IL1A | 3.12× 10−28 |
| ko05202 | Transcriptional misregulation in cancers | MEN1, TAF15, NCOR1, FLT1, CEBPA, SPINT1, ZBTB16, SIX4, GOLPH3, AFF1, KLF3, JMJD1C, TRAF1, SP1, PLAU, PBX1, TP53, CCND2, IL6, ELK4, MMP9, BCL6, EWSR1 | 4.43× 10−160 |
| ko05142 | Chagas disease (American trypanosomiasis) | TGFB2, IRAK1, ADCY1, FOS, IFNG, CALR, JUN, NOS2, IL6, IFNGR2, IL1B, MYD88, CD3E, IRAK4, GNAQ | 3.41× 10−103 |
| ko05140 | Leishmaniasis | MARCKSL1, ITGB1, TGFB2, MYD88, FOS, IFNG, JUN, NOS2, IFNGR2, IL1A, IL1B, IRAK1, IRAK4 | 8.32× 10−71 |
| ko05412 | Arrhythmogenic right ventricular cardiomyopathy (ARVC) | ITGB1, ITGA9, ITGB3, SGCD, GJA1, CACNB2, CACNA1C, CACNA1D, TCF7, LEF1, CACNG4 | 5.27× 10−89 |
| ko04540 | Gap junction | GJA1, PDGFRA, GNAQ, PDGFRB, DRD2, ADCY9, SRC, GRB2, ADCY1 | 7.36× 10−78 |
| ko03015 | mRNA surveillance pathway | SMG5, PABPN1, SMG6, RNPS1, SYMPK, CSTF2, SAP18, UPF2, ACIN1, RNMT | 8.17× 10−83 |
| ko04930 | Type II diabetes mellitus | SOCS4, CACNA1C, SOCS1, SOCS3, SLC2A4, CACNA1D | 1.12× 10−50 |
| GO Class | Contributor | Reference |
|---|---|---|
| negative regulation of tumor necrosis factor production | ARUK-UCL | PMID:23595570 |
| negative regulation of cell migration involved in sprouting angiogenesis | BHF-UCL | PMID:24463821 |
| negative regulation of blood vessel endothelial cell proliferation involved in sprouting angiogenesis | BHF-UCL | PMID:24463821 |
| negative regulation of endothelial cell chemotaxis to fibroblast growth factor | BHF-UCL | PMID:24463821 |
| mRNA binding involved in posttranscriptional gene silencing | ARUK-UCL | PMID:23595570 |
| negative regulation of epithelial to mesenchymal transition | ARUK-UCL | PMID:25916550 |
| negative regulation of cell migration | ARUK-UCL | PMID:26498692 |
| negative regulation of interleukin-6 production | ARUK-UCL | PMID:24299952 |
| positive regulation of transforming growth factor beta3 production | ARUK-UCL | GO_REF:0000024 |
| positive regulation of collagen biosynthetic process | ARUK-UCL | GO_REF:0000024 |
| gene silencing by miRNA | ARUK-UCL | PMID:23595570 |
| negative regulation of fibroblast growth factor receptor signaling pathway | BHF-UCL | PMID:24463821 |
| cellular response to fibroblast growth factor stimulus | BHF-UCL | PMID:24463821 |
| negative regulation of inflammatory response | ARUK-UCL | GO_REF:0000024 |
| negative regulation of stress fiber assembly | ARUK-UCL | PMID:26498692 |
| negative regulation of NIK/NF-kappaB signaling | ARUK-UCL | GO_REF:0000024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Kadkhoda, S.; Taheri, M.; Tafrishinejad, A. A Review on the Role of miR-149-5p in the Carcinogenesis. Int. J. Mol. Sci. 2022, 23, 415. https://doi.org/10.3390/ijms23010415
Ghafouri-Fard S, Khoshbakht T, Hussen BM, Kadkhoda S, Taheri M, Tafrishinejad A. A Review on the Role of miR-149-5p in the Carcinogenesis. International Journal of Molecular Sciences. 2022; 23(1):415. https://doi.org/10.3390/ijms23010415
Chicago/Turabian StyleGhafouri-Fard, Soudeh, Tayyebeh Khoshbakht, Bashdar Mahmud Hussen, Sepideh Kadkhoda, Mohammad Taheri, and Arash Tafrishinejad. 2022. "A Review on the Role of miR-149-5p in the Carcinogenesis" International Journal of Molecular Sciences 23, no. 1: 415. https://doi.org/10.3390/ijms23010415