
The Effect of Fullerenol C60(OH)36 on the Antioxidant Defense System in Erythrocytes

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Total Concentration of -SH Groups in Erythrocytes Incubated with C60(OH)36
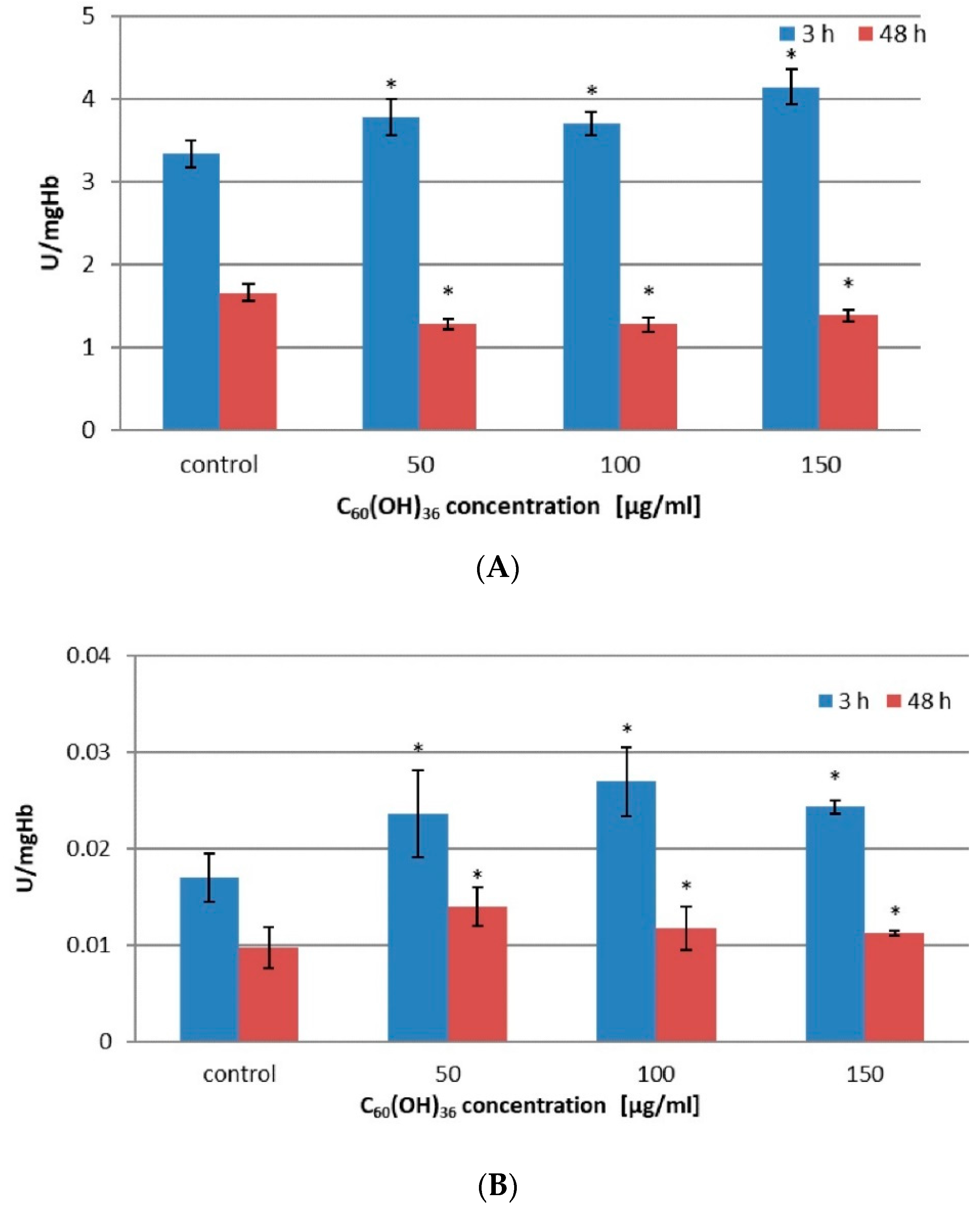
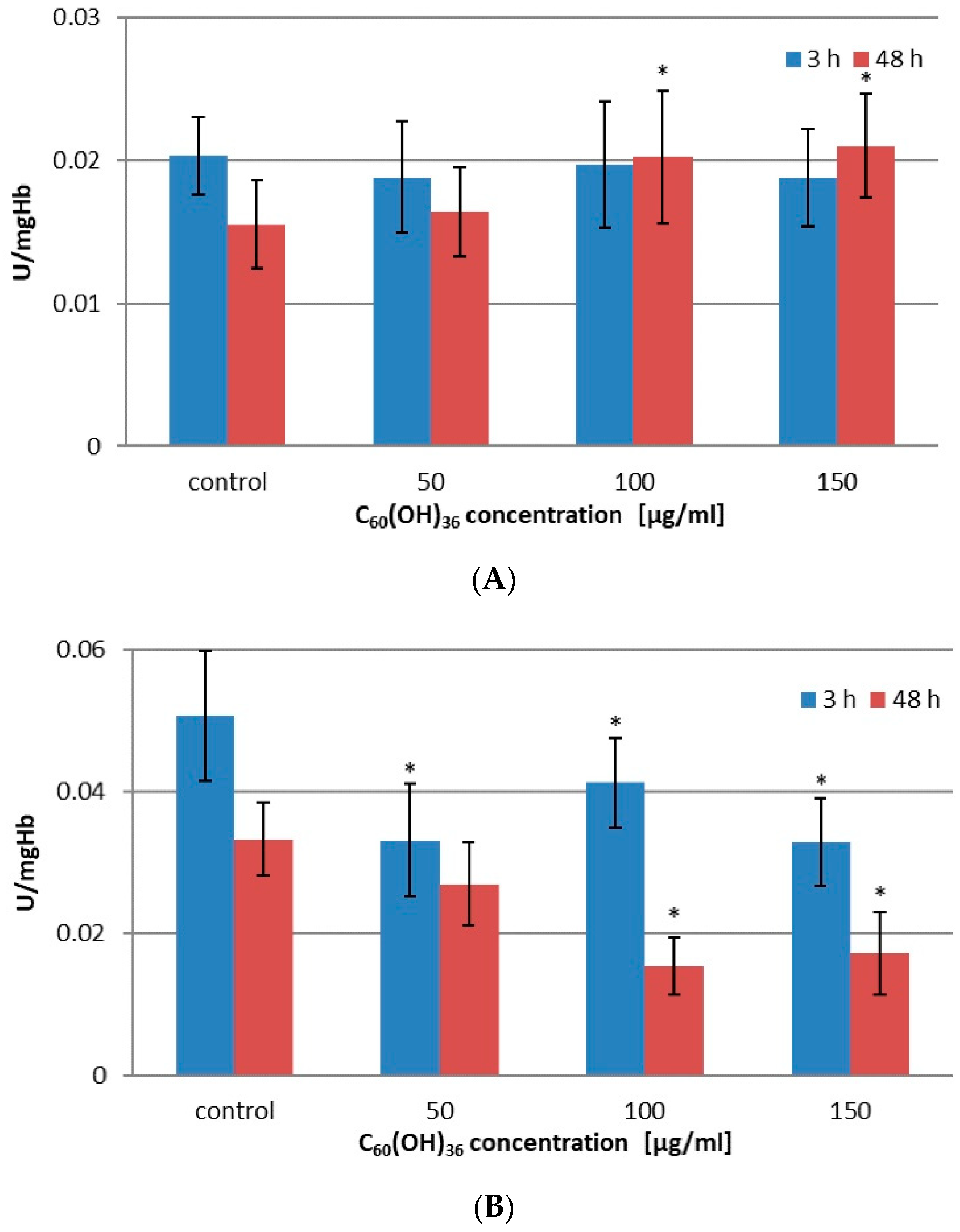
2.2. Activity of Enzymes in Erythrocytes Incubated with C60(OH)36
2.3. The Effect of C60(OH)36 on Erythrocyte Microviscosity
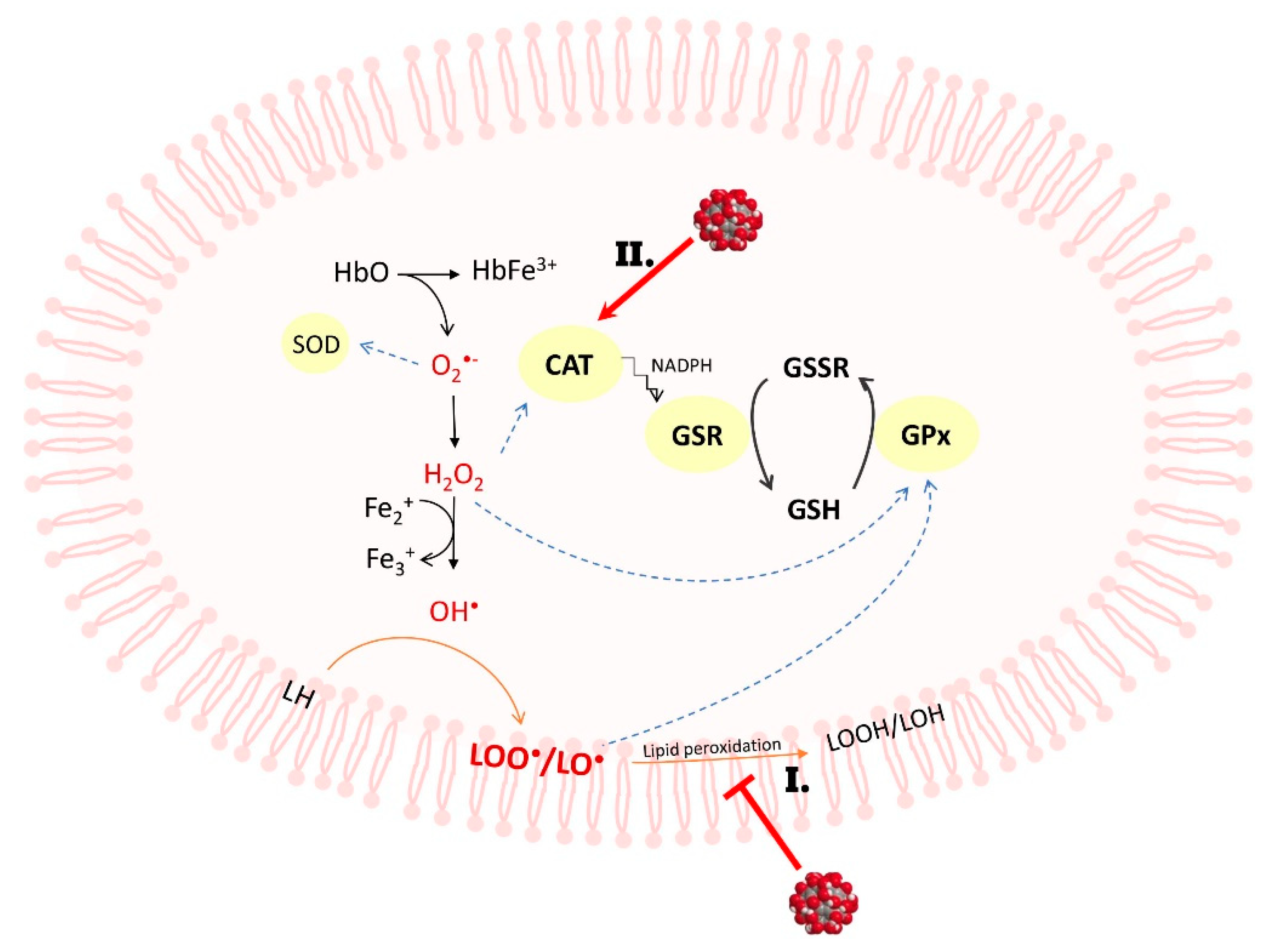
3. Discussion
4. Materials and Methods
4.1. Synthesis of Fullerenol
4.2. Erythrocyte Isolation
4.3. Hemolysate Preparation
4.4. Determination of Hemoglobin Concentration in Hemolysate Using the Drabkin Method
4.5. Fullerenol Treatment of Erythorocytes
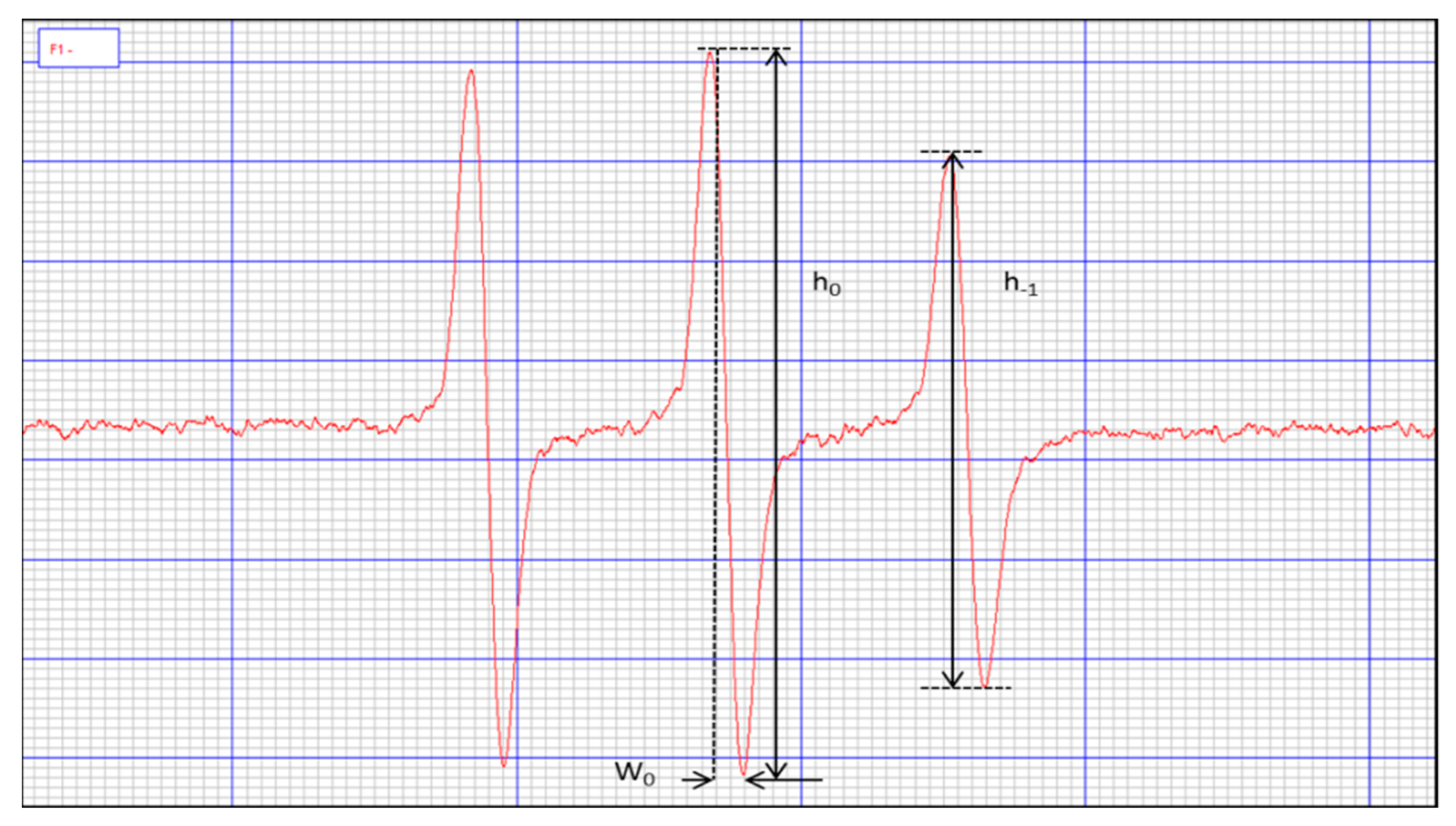
4.6. Identification of Erythrocyte Microviscosity
- k = 6.5 × 10−9 s/mT
- W0 = width of the spectral center line
- h0 = height of the spectral midfield line
- h–1 = height of the highfield line of the spectrum
- τR = rotational correlation time of the tracer inside erythrocytes
- τB = rotational correlation time of the tracer dissolved in a solution of known viscosity
- ηR = erythrocyte inner viscosity
- ηB = viscosity of known solution
4.7. Total Concentration of -SH Groups in Erythrocytes
4.8. Determination of Catalase Activity
4.9. Determination of Glutathione Peroxidase Activity
4.10. Determination of Glutathione Transferase Activity
4.11. Determination of Glutathione Reductase Activity
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meng, Q.; Zaharieva, E.K.; Sasatani, M.; Kobayashi, J. Possible relationship between mitochondrial changes and oxidative stress under low dose-rate irradiation. Redox Rep. 2021, 26, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Remacle, J.; Michiels, C.; Raes, M. The importance of antioxidant enzymes in cellular aging and degeneration. EXS 1992, 62, 99–108. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Su, S.Y.; Fang, M.Y. Zinc supplement modulates oxidative stress and antioxidant values in rats with severe acute pancreatitis. Biol. Trace Elem. Res. 2014, 159, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Pannuru, P.; Vaddi, D.R.; Kindinti, R.R.; Varadacharyulu, N. Increased erythrocyte antioxidant status protects against smoking induced hemolysis in moderate smokers. Hum. Exp. Toxicol. 2011, 30, 1475–1481. [Google Scholar] [CrossRef]
- Milic, V.D.; Stankov, K.; Injac, R.; Djordjevic, A.; Srdjenovic, B.; Govedarica, B.; Radic, N.; Simic, V.D.; Strukelj, B. Activity of antioxidative enzymes in erythrocytes after a single dose administration of doxorubicin in rats pretreated with fullerenol C60(OH)24. Toxicol. Mech. Methods 2009, 19, 24–28. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Grebowski, J.; Kazmierska, P.; Krokosz, A. Fullerenols as a new therapeutic approach in nanomedicine. Biomed. Res. Int. 2013, 2013, 751913. [Google Scholar] [CrossRef] [Green Version]
- Krokosz, A.; Grebowski, J.; Rodacka, A.; Pasternak, B.; Puchala, M. The effect of fullerenol C60(OH)~30 on the alcohol dehydrogenase activity irradiated with X-rays. Radiat. Phys. Chem. 2014, 97, 102–106. [Google Scholar] [CrossRef]
- Golomidov, I.; Bolshakova, O.; Komissarov, A.; Sharoyko, V.; Slepneva, E.; Slobodina, A.; Latypova, E.; Zherebyateva, O.; Tennikova, T.; Sarantseva, S. The neuroprotective effect of fullerenols on a model of Parkinson’s disease in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2020, 523, 446–451. [Google Scholar] [CrossRef]
- Kojić, D.; Purać, J.; Čelić, T.V.; Jović, D.; Vukašinović, E.L.; Pihler, I.; Borišev, I.; Djordjevic, A. Effect of fullerenol nanoparticles on oxidative stress induced by paraquat in honey bees. Environ. Sci. Pollut. Res. Int. 2020, 27, 6603–6612. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, F.; Iqbal, M.; Ali, M.; Ashraf, M.A. Fullerenol regulates oxidative stress and tissue ionic homeostasis in spring wheat to improve net-primary productivity under salt-stress. Ecotoxicol. Environ. Saf. 2021, 211, 111901. [Google Scholar] [CrossRef]
- Djordjevic, A.; Srdjenovic, B.; Seke, M.; Petrovic, D.; Injac, R.; Mrdjanovic, J. Review of Synthesis and Antioxidant Potential of Fullerenol Nanoparticles. J. Nanomater. 2015, 2015, 567073. [Google Scholar] [CrossRef] [Green Version]
- Markovic, Z.; Trajkovic, V. Biomedical potential of the reactive oxygen species generation and quenching by fullerenes (C60). Biomaterials 2008, 29, 3561–3573. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.J.; Lao, F.; Fu, P.P.; Wamer, W.G.; Zhao, Y.; Wang, P.C.; Qiu, Y.; Sun, B.; Xing, G.; Dong, J.; et al. The scavenging of reactive oxygen species and the potential for cell protection by functionalized fullerene materials. Biomaterials 2009, 30, 611–621. [Google Scholar] [CrossRef]
- Kazmierska-Grebowska, P.; Kobrzycka, A.; Bocian, R.; Kowalczyk, T.; Krokosz, A.; Grebowski, J. Fullerenol C60(OH)36 at relatively high concentrations impairs hippocampal theta oscillations (in vivo and in vitro) and triggers epilepsy (in vitro)–A dose response study. Exp. Mol. Pathol. 2018, 105, 98–109. [Google Scholar] [CrossRef]
- Grebowski, J.; Konopko, A.; Krokosz, A.; DiLabio, G.A.; Litwinienko, G. Antioxidant activity of highly hydroxylated fullerene C. Free Radic. Biol. Med. 2020, 160, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, S.; Da Ros, T.; Conde, J.; Sefat, F.; Mozafari, M. Fullerene: Biomedical engineers get to revisit an old friend. Mater. Today 2017, 20, 460–480. [Google Scholar] [CrossRef] [Green Version]
- Dawid, A.; Górny, K.; Gburski, Z. The structural studies of fullerenol C60(OH)24 and nitric oxide mixture in water solvent–MD simulation. Nitric Oxide 2011, 25, 373–380. [Google Scholar] [CrossRef]
- Lao, F.; Li, W.; Han, D.; Qu, Y.; Liu, Y.; Zhao, Y.; Chen, C. Fullerene derivatives protect endothelial cells against NO-induced damage. Nanotechnology 2009, 20, 225103. [Google Scholar] [CrossRef]
- Mirkov, S.M.; Djordjevic, A.N.; Andric, N.L.; Andric, S.A.; Kostic, T.S.; Bogdanovic, G.M.; Vojinovic-Miloradov, M.B.; Kovacevic, R.Z. Nitric oxide-scavenging activity of polyhydroxylated fullerenol, C60(OH)24. Nitric Oxide 2004, 11, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.J.; Ahamed, M.; Alhadlaq, H.A.; Alshamsan, A. Mechanism of ROS scavenging and antioxidant signalling by redox metallic and fullerene nanomaterials: Potential implications in ROS associated degenerative disorders. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Srdjenovic, B.; Milic-Torres, V.; Grujic, N.; Stankov, K.; Djordjevic, A.; Vasovic, V. Antioxidant properties of fullerenol C60(OH)24 in rat kidneys, testes, and lungs treated with doxorubicin. Toxicol. Mech. Methods 2010, 20, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Grebowski, J.; Krokosz, A.; Puchala, M. Fullerenol C60(OH)36 could associate to band 3 protein of human erythrocyte membranes. Biochim. Biophys. Acta 2013, 1828, 2007–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharoyko, V.V.; Iamalova, N.R.; Ageev, S.V.; Meshcheriakov, A.A.; Iurev, G.O.; Petrov, A.V.; Nerukh, D.A.; Farafonov, V.S.; Vasina, L.V.; Penkova, A.V.; et al. In Vitro and In Silico Investigation of Water-Soluble Fullerenol C60(OH)24: Bioactivity and Biocompatibility. J. Phys. Chem. B 2021, 125, 9197–9212. [Google Scholar] [CrossRef] [PubMed]
- Grebowski, J.; Kaźmierska, P.; Krokosz, A. Fullerenol–Properties and applications in biomedical sciences. Postȩpy Hig. I Med. Doświadczalnej 2013, 67, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Grebowski, J.; Krokosz, A. The Effect of Highly Hydroxylated Fullerenol C60(OH)36 on Human Erythrocyte Membrane Organization. J. Spectrosc. 2015, 2015, 825914. [Google Scholar] [CrossRef] [Green Version]
- Lichota, A.; Piwoński, I.; Michlewska, S.; Krokosz, A. A Multiparametric Study of Internalization of Fullerenol C60(OH)36 Nanoparticles into Peripheral Blood Mononuclear Cells: Cytotoxicity in Oxidative Stress Induced by Ionizing Radiation. Int. J. Mol. Sci. 2020, 21, 2281. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Chi, M.C.; Peng, K.T.; Chiang, Y.C.; Hsu, L.F.; Yan, Y.L.; Li, H.Y.; Chen, M.C.; Lee, I.T.; Lai, C.H. Water-Soluble Fullerenol C60(OH)36 toward Effective Anti-Air Pollution Induced by Urban Particulate Matter in HaCaT Cell. Int. J. Mol. Sci. 2019, 20, 4259. [Google Scholar] [CrossRef] [Green Version]
- Krokosz, A.; Grebowski, J.; Szweda-Lewandowska, Z.; Rodacka, A.; Puchala, M. Can melatonin delay oxidative damage of human erythrocytes during prolonged incubation? Adv. Med. Sci. 2013, 58, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Qasim, N.; Mahmood, R. Diminution of Oxidative Damage to Human Erythrocytes and Lymphocytes by Creatine: Possible Role of Creatine in Blood. PLoS ONE 2015, 10, e0141975. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Yang, G.; Wang, H.; Liu, J.; Zheng, J.; Lv, B.; Yang, M.; Yang, Y.; Gao, C.; Guo, Y. Aging erythrocyte membranes as biomimetic nanometer carriers of liver-targeting chromium poisoning treatment. Drug Deliv. 2021, 28, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Remigante, A.; Morabito, R.; Marino, A. Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants 2019, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remigante, A.; Morabito, R.; Marino, A. Band 3 protein function and oxidative stress in erythrocytes. J. Cell. Physiol. 2021, 236, 6225–6234. [Google Scholar] [CrossRef]
- Grebowski, J.; Kazmierska, P.; Litwinienko, G.; Lankoff, A.; Wolszczak, M.; Krokosz, A. Fullerenol C60(OH)36 protects human erythrocyte membrane against high-energy electrons. Biochim. Biophys. Acta (BBA)–Biomembr. 2018, 1860, 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Morse, P.D., II. Use of the spin label tempamine for measuring the internal viscosity of red blood cells. Biochem. Biophys. Res. Commun. 1977, 77, 1486–1491. [Google Scholar] [CrossRef]
- Bartosz, G.; Gaczyńska, M.; Grzelińska, E.; Judkiewicz, L. A spin-label study of membrane proteins and internal microviscosity of erythrocytes in hereditary spherocytosis. Life Sci. 1987, 41, 2285–2288. [Google Scholar] [CrossRef]
- Injac, R.; Prijatelj, M.; Strukelj, B. Fullerenol nanoparticles: Toxicity and antioxidant activity. Methods Mol. Biol. 2013, 1028, 75–100. [Google Scholar] [CrossRef]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Markers of oxidative stress in erythrocytes and plasma during aging in humans. Oxid. Med. Cell. Longev. 2010, 3, 2–12. [Google Scholar] [CrossRef]
- Yamawaki, H.; Iwai, N. Cytotoxicity of water-soluble fullerene in vascular endothelial cells. Am. J. Physiol. Cell. Physiol. 2006, 290, C1495–C1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkman, H.N.; Galiano, S.; Gaetani, G.F. The function of catalase-bound NADPH. J. Biol. Chem. 1987, 262, 660–666. [Google Scholar] [CrossRef]
- Bocedi, A.; Fabrini, R.; Lai, O.; Alfieri, L.; Roncoroni, C.; Noce, A.; Pedersen, J.Z.; Ricci, G. Erythrocyte glutathione transferase: A general probe for chemical contaminations in mammals. Cell. Death. Discov. 2016, 2, 16029. [Google Scholar] [CrossRef] [Green Version]
- Bocedi, A.; Noce, A.; Marrone, G.; Noce, G.; Cattani, G.; Gambardella, G.; Di Lauro, M.; Di Daniele, N.; Ricci, G. Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution. Nutrients 2019, 11, 1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchewka, Z.; Piwowar, A.; Ruzik, S.; Długosz, A. Glutathione S–transferases class Pi and Mi and their significance in oncology. Postepy. Hig. Med. Dosw. 2017, 71, 541–550. [Google Scholar] [CrossRef]
- Rebrova, T.Y.; Afanas’ev, S.A.; Putrova, O.D.; Popov, S.V. Age-related characteristics of erythrocyte membrane microviscosity in experimental cardiosclerosis. Adv. Gerontol. 2013, 3, 211–214. [Google Scholar] [CrossRef]
- Blat, A.; Stepanenko, T.; Bulat, K.; Wajda, A.; Dybas, J.; Mohaissen, T.; Alcicek, F.C.; Szczesny-Malysiak, E.; Malek, K.; Fedorowicz, A.; et al. Spectroscopic Signature of Red Blood Cells in a D-Galactose-Induced Accelerated Aging Model. Int. J. Mol. Sci. 2021, 22, 2660. [Google Scholar] [CrossRef]
- Himbert, S.; Alsop, R.J.; Rose, M.; Hertz, L.; Dhaliwal, A.; Moran-Mirabal, J.M.; Verschoor, C.P.; Bowdish, D.M.; Kaestner, L.; Wagner, C.; et al. The Molecular Structure of Human Red Blood Cell Membranes from Highly Oriented, Solid Supported Multi-Lamellar Membranes. Sci. Rep. 2017, 7, 39661. [Google Scholar] [CrossRef] [Green Version]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for Life–Properties of Red Blood Cell Deformability. Front. Physi. 2018, 9, 656. [Google Scholar] [CrossRef]
- Grebowski, J.; Krokosz, A.; Puchala, M. Membrane fluidity and activity of membrane ATPases in human erythrocytes under the influence of polyhydroxylated fullerene. Biochim. Biophys. Acta 2013, 1828, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Sousa, L.; Garcia, I.J.; Costa, T.G.; Silva, L.N.; Renó, C.O.; Oliveira, E.S.; Tilelli, C.Q.; Santos, L.L.; Cortes, V.F.; Santos, H.L.; et al. Effects of Iron Overload on the Activity of Na,K-ATPase and Lipid Profile of the Human Erythrocyte Membrane. PLoS ONE 2015, 10, e0132852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radosinska, J.; Vrbjar, N. Erythrocyte Deformability and Na,K-ATPase Activity in Various Pathophysiological Situations and Their Protection by Selected Nutritional Antioxidants in Humans. Int. J. Mol. Sci. 2021, 22, 11924. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; He, P.; Zhang, J.M.; Jiang, H.; Zhu, S.Z. Novel and Efficient Synthesis of Water-Soluble [60]Fullerenol by Solvent-Free Reaction. Synth. Commun. 2005, 35, 1803–1808. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C60(OH)36 Concentration [µg/mL] | Incubation Time | |||||
|---|---|---|---|---|---|---|
| 3 h | 48 h | |||||
| Parameter P | SD | Thiol Group Concentration [µM] | Parameter P | SD | Thiol Group Concentration [µM] | |
| 0 | 0.93 | 0.04 | 22.93 | 0.37 | 0.05 | 9.22 |
| 50 | 0.89 | 0.06 | 22.12 | 0.39 | 0.04 | 9.56 |
| 100 | 0.92 | 0.13 | 22.68 | 0.49 * | 0.05 | 10.54 |
| 150 | 0.92 | 0.07 | 22.71 | 0.52 * | 0.07 | 10.72 |
| Concentration C60(OH)36 [µg/mL] | CAT Activity ± SD [U/mgHb] | GPx Activity ± SD [U/mgHb] | ||
|---|---|---|---|---|
| 3 h | 48 h | 3 h | 48 h | |
| 0 | 3.337 ± 0.166 | 1.662 ± 0.105 | 0.0169 ± 0.0025 | 0.0097 ± 0.0021 |
| 50 | 3.781 * ± 0.220 | 1.280 * ± 0.066 | 0.0236 * ± 0.0045 | 0.0139 * ± 0.0020 |
| 100 | 3.701 * ± 0.138 | 1.277 * ± 0.088 | 0.0269 * ± 0.0036 | 0.0127 * ± 0.0022 |
| 150 | 4.146 * ± 0.206 | 1.384 * ± 0.074 | 0.0243 * ± 0.0007 | 0.0124 * ± 0.0003 |
| Concentration C60(OH)36 [µg/mL] | GSR Activity ± SD [U/mgHb] | GST Activity ± SD [U/mgHb] | ||
|---|---|---|---|---|
| 3 h | 48 h | 3 h | 48 h | |
| 0 | 0.0203 ± 0.0027 | 0.0155 ± 0.0031 | 0.0506 ± 0.0092 | 0.0333 ± 0.0052 |
| 50 | 0.0188 ± 0.0039 | 0.0164 ± 0.0031 | 0.0331 * ± 0.0080 | 0.0269 ± 0.0059 |
| 100 | 0.0197 ± 0.0044 | 0.0202 * ± 0.0046 | 0.0412 * ± 0.0063 | 0.0153 * ± 0.0040 |
| 150 | 0.0188 ± 0.0034 | 0.0210 * ± 0.0036 | 0.0329 * ± 0.0062 | 0.0172 * ± 0.0058 |
| Concentration C60(OH)36 [µg/mL] | η ± SD * [Pa × s] | |
|---|---|---|
| 3 h | 48 h | |
| 0 | 4.64 ± 0.57 | 5.17 ± 0.59 |
| 50 | 5.14 ± 0.38 | 5.72 ± 0.56 |
| 100 | 4.47 ± 0.29 | 5.32 ± 0.73 |
| 150 | 4.72 ± 0.99 | 5.15 ± 0.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grebowski, J.; Kazmierska-Grebowska, P.; Cichon, N.; Piotrowski, P.; Litwinienko, G. The Effect of Fullerenol C60(OH)36 on the Antioxidant Defense System in Erythrocytes. Int. J. Mol. Sci. 2022, 23, 119. https://doi.org/10.3390/ijms23010119
Grebowski J, Kazmierska-Grebowska P, Cichon N, Piotrowski P, Litwinienko G. The Effect of Fullerenol C60(OH)36 on the Antioxidant Defense System in Erythrocytes. International Journal of Molecular Sciences. 2022; 23(1):119. https://doi.org/10.3390/ijms23010119
Chicago/Turabian StyleGrebowski, Jacek, Paulina Kazmierska-Grebowska, Natalia Cichon, Piotr Piotrowski, and Grzegorz Litwinienko. 2022. "The Effect of Fullerenol C60(OH)36 on the Antioxidant Defense System in Erythrocytes" International Journal of Molecular Sciences 23, no. 1: 119. https://doi.org/10.3390/ijms23010119