
The Ambivalent Role of Skin Microbiota and Adrenaline in Wound Healing and the Interplay between Them


Abstract
:1. The Ambivalent Role of the Skin Microbiota in Wound Healing
2. Certain Members of the Skin Microbiota Can Have a Negative Impact on Wound Healing
3. Positive Effect of Some Commensals and Probiotics on Wound Healing
4. Adrenergic Receptors (ARs) Are Expressed in Many Cells of the Skin and Contribute to Wound Healing
4.1. Keratinocytes
4.2. Immune Cells
4.3. Fibroblasts
4.4. Blood Vessels
5. Interplay between Adrenaline and Skin Bacteria
5.1. TA-Producing Skin Microbiota Might Accelerate Wound Healing via ARs
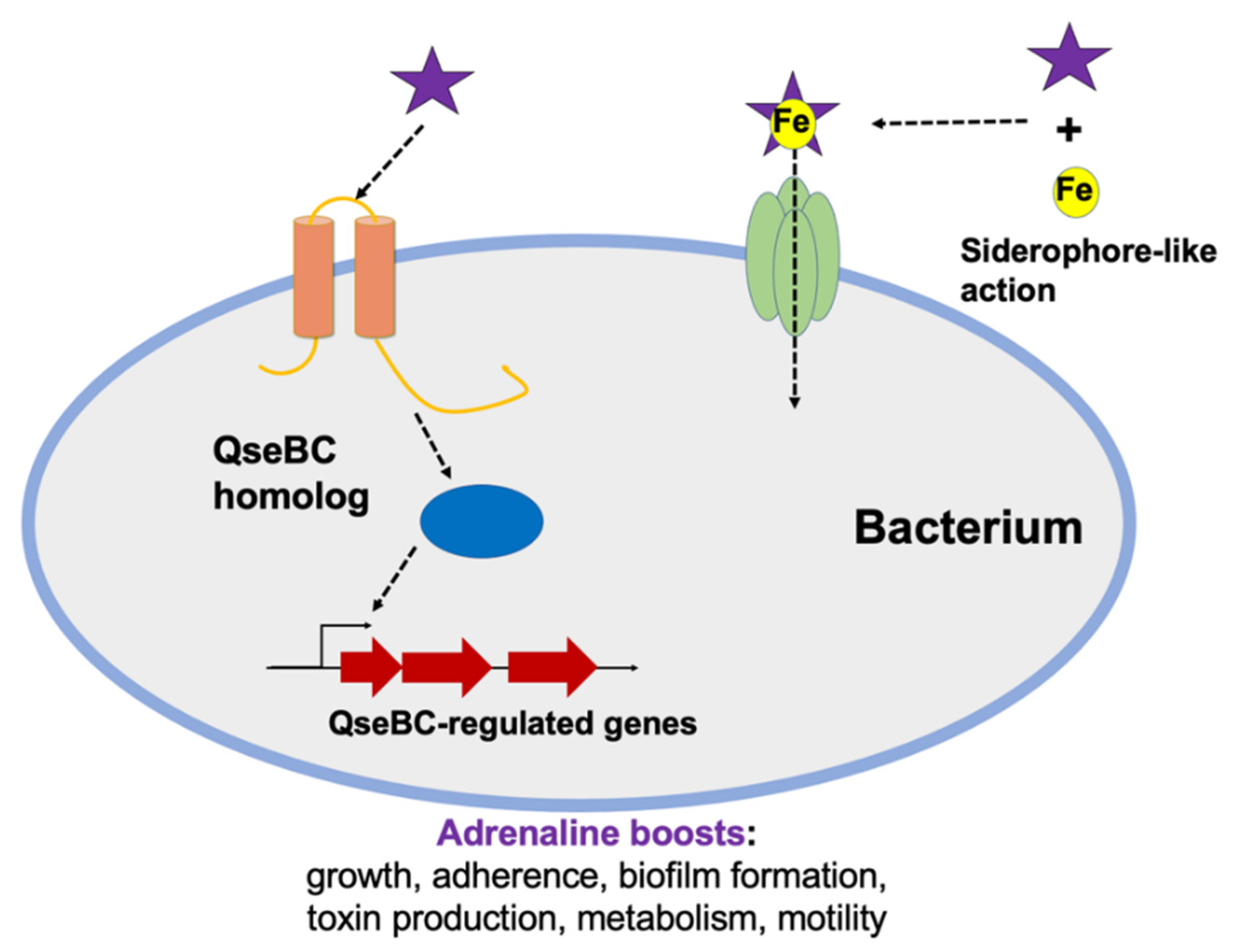
5.2. Adrenaline Controls Not Only Sympathetic Nervous System but Also Quorum Sensing in Bacteria
6. Topical Probiotics Are a Therapeutic Option in the Treatment of Chronic Wounds
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARs | adrenergic receptors |
| ß2-ARs | beta-2-adrenergic-receptors |
| cAMP | cyclic-adenosine monophosphate |
| DOP | dopamine |
| EPI | epinephrine (adrenaline) |
| ERK | extracellular signal-regulated kinase |
| FGF2 | fibroblast growth factor 2 |
| KGF1 | keratinocyte growth factor 1 |
| PKA | protein kinase A |
| PEA | phenethylamine |
| PP2A | phosphatase 2A |
| sadA | gene encoding staphylococcal aromatic amino acid decarboxylase |
| SadA | enzyme of staphylococcal aromatic amino acid decarboxylase |
| SNS | sympathetic nervous system |
| TAs | trace amines (PEA, TRY, TYM) |
| TRY | tryptamine |
| TYM | tyramine |
| VEGF-A | vascular endothelial growth factor A |
References
- Casey, G. The stages of wound healing. Nurs. N. Z. 1997, 3, 31. [Google Scholar]
- Gilmore, M.A. Phases of wound healing. Dimens. Oncol. Nurs. 1991, 5, 32–34. [Google Scholar]
- Heng, M.C.Y. Wound healing in adult skin: Aiming for perfect regeneration. Int. J. Derm. 2011, 50, 1058–1066. [Google Scholar] [CrossRef]
- Martin, P. Wound Healing--Aiming for Perfect Skin Regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Beyer, S.; Koch, M.; Lee, Y.H.; Jung, F.; Blocki, A. An In Vitro Model of Angiogenesis during Wound Healing Provides Insights into the Complex Role of Cells and Factors in the Inflammatory and Proliferation Phase. Int. J. Mol. Sci. 2018, 19, 2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in Wound Repair: Molecular and Cellular Mechanisms. J. Investig. Derm. 2007, 127, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, R.; Harding, K.G. Bacteria and wound healing. Curr. Opin. Infect. Dis. 2004, 17, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.-J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.; et al. Commensal–dendritic-cell interaction specifies a unique protective skin immune signature. Nat. Cell Biol. 2015, 520, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Snitkin, E.S.; Yockey, L.J.; Bermudez, D.M.; Liechty, K.W.; Segre, J.A.; Mullikin, J.; Blakesley, R.; Young, A.; Chu, G.; et al. Longitudinal shift in diabetic wound microbiota correlates with prolonged skin defense response. Proc. Natl. Acad. Sci. USA 2010, 107, 14799–14804. [Google Scholar] [CrossRef] [Green Version]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain- and Species-Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host Microbe 2019, 25, 641–655.e5. [Google Scholar] [CrossRef]
- Dowd, S.E.; Sun, Y.; Secor, P.R.; Rhoads, D.D.; Wolcott, B.M.; James, G.A.; Wolcott, R.D. Survey of bacterial diversity in chronic wounds using Pyrosequencing, DGGE, and full ribosome shotgun sequencing. BMC Microbiol. 2008, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, R.A.; Hsieh, J.-C.; Galiano, R.D. The Impact of Biofilm Formation on Wound Healing. In Wound Healing—Current Perspectives; IntechOpen: London, UK, 2019. [Google Scholar]
- Mihai, M.M.; Holban, A.M.; Giurcăneanu, C.; Popa, L.G.; Buzea, M.; Filipov, M.; Lazăr, V.; Chifiriuc, M.C.; Popa, M.I. Identification and phenotypic characterization of the most frequent bacterial etiologies in chronic skin ulcers. Romanian J. Morphol. Embryol. 2014, 55, 1401–1408. [Google Scholar]
- Park, J.-U.; Oh, B.; Lee, J.P.; Choi, M.-H.; Lee, M.-J.; Kim, B.-S. Influence of Microbiota on Diabetic Foot Wound in Comparison with Adjacent Normal Skin Based on the Clinical Features. Biomed. Res. Int. 2019, 2019, 7459236. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.; Wall, I.; Wilson, M.; Hill, K.; Davies, C.; Hill, C.; Harding, K.; Thomas, D. Anaerobic cocci populating the deep tissues of chronic wounds impair cellular wound healing responses in vitro. Br. J. Derm. 2003, 148, 456–466. [Google Scholar] [CrossRef]
- Kim, J.H.; Yang, B.; Tedesco, A.; Lebig, E.G.D.; Ruegger, P.M.; Xu, K.; Borneman, J.; Martins-Green, M. High Levels of Oxidative Stress and Skin Microbiome are Critical for Initiation and Development of Chronic Wounds in Diabetic Mice. Sci. Rep. 2019, 9, 19318. [Google Scholar] [CrossRef] [Green Version]
- Luqman, A.; Zabel, S.; Rahmdel, S.; Merz, B.; Gruenheit, N.; Harter, J.; Nieselt, K.; Götz, F. The Neuromodulator-Encoding sadA Gene Is Widely Distributed in the Human Skin Microbiome. Front. Microbiol. 2020, 11, 573679. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Joo, H.-S.; Chatterjee, S.S.; Otto, M. Phenol-soluble modulins—Critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 2014, 38, 698–719. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Genet. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin Wound Healing Is Accelerated and Scarless in the Absence of Commensal Microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.; Sanford, N.; Gabrilska, R.; Oates, J.; Wilkinson, J.; Rumbaugh, K. Microbiota is a primary cause of pathogenesis of chronic wounds. J. Wound Care 2016, 25, S33–S43. [Google Scholar] [CrossRef] [PubMed]
- Flowers, L.; Grice, E.A. The Skin Microbiota: Balancing Risk and Reward. Cell Host Microbe 2020, 28, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.G.; Edwards, R.; Thomas, D.W. Inflammation and Wound Healing: The Role of Bacteria in the Immuno-Regulation of Wound Healing. Int. J. Low. Extrem. Wounds 2004, 3, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Raymond, S.L.; Holden, D.C.; Mira, J.C.; Stortz, J.A.; Loftus, T.J.; Mohr, A.M.; Moldawer, L.L.; Moore, F.A.; Larson, S.D.; Efron, P.A. Microbial recognition and danger signals in sepsis and trauma. Biochim. Biophys Acta Mol. Basis Dis. 2017, 1863, 2564–2573. [Google Scholar] [CrossRef] [PubMed]
- Strbo, N.; Yin, N.; Stojadinovic, O. Innate and Adaptive Immune Responses in Wound Epithelialization. Adv. Wound Care 2014, 3, 492–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Götz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin Commensals Amplify the Innate Immune Response to Pathogens by Activation of Distinct Signaling Pathways. J. Investig. Derm. 2011, 131, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Di Domizio, J.; Belkhodja, C.; Chenuet, P.; Fries, A.; Murray, T.; Mondéjar, P.M.; DeMaria, O.; Conrad, C.; Homey, B.; Werner, S.; et al. The commensal skin microbiota triggers type I IFN–dependent innate repair responses in injured skin. Nat. Immunol. 2020, 21, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.D.; Ledo, C.; Cela, E.; Stella, I.; Xu, C.; Ojeda, D.S.; Frenette, P.S.; Gómez, M.I. The good side of inflammation: Staphylococcus aureus proteins SpA and Sbi contribute to proper abscess formation and wound healing during skin and soft tissue infections. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 2657–2670. [Google Scholar] [CrossRef]
- Paharik, A.E.; Parlet, C.P.; Chung, N.; Todd, D.A.; Rodriguez, E.I.; Van Dyke, M.J.; Cech, N.B.; Horswill, A.R. Coagulase-Negative Staphylococcal Strain Prevents Staphylococcus aureus Colonization and Skin Infection by Blocking Quorum Sensing. Cell Host Microbe 2017, 22, 746–756.e5. [Google Scholar] [CrossRef] [Green Version]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Marschal, M.; et al. Human commensals producing a novel antibiotic impair pathogen colonization. Nat. Cell Biol. 2016, 535, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Goncheva, M.I.; Flannagan, R.S.; Deecker, S.R.; Guariglia-Oropeza, V.; Ensminger, A.W.; Heinrichs, D.E. Coagulase-negative staphylococci release a purine analog that inhibits Staphylococcus aureus virulence. Nat. Commun. 2021, 12, 1887. [Google Scholar] [CrossRef] [PubMed]
- Götz, F.; Perconti, S.; Popella, P.; Werner, R.; Schlag, M. Epidermin and gallidermin: Staphylococcal lantibiotics. Int. J. Med. Microbiol. 2014, 304, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Schnell, N.; Entian, K.-D.; Schneider, U.; Götz, F.; Zähner, H.; Kellner, R. Prepeptide sequence of epidermin, a ribosomally synthesized antibiotic with four sulphide-rings. Nat. Cell Biol. 1988, 333, 276–278. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Fijan, S.; Frauwallner, A.; Langerholc, T.; Krebs, B.; ter Haar née Younes, J.A.; Heschl, A.; Turk, D.M.; Rogelj, I. Efficacy of Using Probiotics with Antagonistic Activity against Pathogens of Wound Infections: An Integrative Review of Literature. BioMed Res. Int. 2019, 2019, 7585486. [Google Scholar] [CrossRef] [Green Version]
- Vågesjö, E.; Öhnstedt, E.; Mortier, A.; Lofton, H.; Huss, F.; Proost, P.; Roos, S.; Phillipson, M. Accelerated wound healing in mice by on-site production and delivery of CXCL12 by transformed lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Ruegger, P.R.; Lebig, E.G.; VanSchalkwyk, S.; Jeske, D.R.; Hsiao, A.; Borneman, J.; Martins-Green, M. High Levels of Oxidative Stress Create a Microenvironment That Significantly Decreases the Diversity of the Microbiota in Diabetic Chronic Wounds and Promotes Biofilm Formation. Front. Cell. Infect. Microbiol. 2020. [Google Scholar] [CrossRef]
- Kennedy, P.; Brammah, S.; Wills, E. Burns, biofilm and a new appraisal of burn wound sepsis. Burns 2010, 36, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Boldock, E.; Surewaard, B.G.J.; Shamarina, D.; Na, M.; Fei, Y.; Ali, A.; Williams, A.; Pollitt, E.J.G.; Szkuta, P.; Morris, P.; et al. Human skin commensals augment Staphylococcus aureus pathogenesis. Nat. Microbiol. 2018, 3, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.; Campbell, L.; Crompton, R.A.; Singh, G.; McHugh, B.J.; Davidson, D.J.; McBain, A.J.; Cruickshank, S.M.; Hardman, M.J. Microbial Host Interactions and Impaired Wound Healing in Mice and Humans: Defining a Role for BD14 and NOD2. J. Invest. Dermatol. 2018, 138, 2264–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the Chronic-Wound Microbiome: Fungal Communities Are Prevalent, Dynamic, and Associated with Delayed Healing. mBio 2016, 7, e01058-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luqman, A.; Muttaqin, M.Z.; Yulaipi, S.; Ebner, P.; Matsuo, M.; Zabel, S.; Tribelli, P.M.; Nieselt, K.; Hidayati, D.; Götz, F. Trace amines produced by skin bacteria accelerate wound healing in mice. Commun. Biol. 2020, 3, 277. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.N.; Rea, M.C.; O’Connor, P.M.; Hill, C.; Ross, R.P. Human skin microbiota is a rich source of bacteriocin-producing staphylococci that kill human pathogens. FEMS Microbiol. Ecol. 2019, 95, fiy241. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.R.; Costa, S.K.; Zaramela, L.S.; Khalil, S.; Todd, D.A.; Winter, H.L.; Sanford, J.A.; O’Neill, A.M.; Liggins, M.C.; Nakatsuji, T.; et al. Quorum sensing between bacterial species on the skin protects against epidermal injury in atopic dermatitis. Sci. Transl. Med. 2019, 11, eaat8329. [Google Scholar] [CrossRef] [PubMed]
- Lopes, E.G.; Moreira, D.A.; Gullón, P.; Gullón, B.; Cardelle-Cobas, A.; Tavaria, F.K. Topical application of probiotics in skin: Adhesion, antimicrobial and antibiofilm in vitro assays. J. Appl. Microbiol. 2017, 122, 450–461. [Google Scholar] [CrossRef]
- Valdéz, J.C.; Peral, M.C.; Rachid, M.; Santana, M.; Perdigón, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: The potential use of probiotics in wound treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Mohammedsaeed, W.; Cruickshank, S.; McBain, A.J.; O’Neill, C.A. Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration. Sci. Rep. 2015, 5, 16147. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris. Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [Google Scholar] [CrossRef] [Green Version]
- Cogen, A.L.; Yamasaki, K.; Muto, J.; Sanchez, K.M.; Alexander, L.C.; Tanios, J.; Lai, Y.; Kim, J.E.; Nizet, V.; Gallo, R.L. Staphylococcus epidermidis antimicrobial delta-toxin (phenol-soluble modulin-gamma) cooperates with host antimicrobial peptides to kill group A Streptococcus. PLoS ONE. 2010, 5, e8557. [Google Scholar] [CrossRef]
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.-R.; Hooper, L.V.; Schmidt, R.R.; von Aulock, S.; et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef]
- Imegwu, O.; Chang, T.H.; Steinberg, J.J.; Levenson, S.M. Staphylococcus aureus peptidoglycan ameliorates cyclophosphamide-induced impairment of wound healing. Wound Repair Regen. 1997, 5, 364–372. [Google Scholar] [CrossRef]
- Linehan, J.L.; Harrison, O.J.; Han, S.J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.; Tamoutounour, S.; et al. Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair. Cell 2018, 172, 784–796.e18. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, T.L.; Levine, S.; Schonfeld-Warden, N.; O’mara, M.S.; Greenhalgh, D.G. Hypothalamic–Pituitary–Adrenal Axis Response to Sustained Stress after Major Burn Injury in Children. J. Burn. Care Res. 2006, 27, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Sedowofia, K.; Barclay, C.; Quaba, A.; Smith, A.; Stephen, R.; Thomson, M.; Watson, A.; McIntosh, N. The systemic stress response to thermal injury in children. Clin. Endocrinol. 1998, 49, 335–341. [Google Scholar] [CrossRef]
- Sivamani, R.K.; Pullar, C.E.; Manabat-Hidalgo, C.G.; Rocke, D.M.; Carlsen, R.C.; Greenhalgh, D.G.; Isseroff, R.R. Stress-Mediated Increases in Systemic and Local Epinephrine Impair Skin Wound Healing: Potential New Indication for Beta Blockers. PLoS Med. 2009, 6, e1000012. [Google Scholar] [CrossRef] [Green Version]
- Stojadinovic, O.; Gordon, K.A.; Lebrun, E.; Tomic-Canic, M. Stress-Induced Hormones Cortisol and Epinephrine Impair Wound Epithelization. Adv. Wound Care 2012, 1, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Romana-Souza, B.; Otranto, M.; Vieira, A.M.; Filgueiras, C.C.; Fierro, I.M.; Monte-Alto-Costa, A. Rotational stress-induced increase in epinephrine levels delays cutaneous wound healing in mice. BrainBehav. Immun. 2010, 24, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Wikberg, J.E. The pharmacological classification of adrenergic alpha 1 and alpha 2 receptors and their mechanisms of action. Acta Physiol. Scand. Suppl. 1979, 468, 1–99. [Google Scholar] [PubMed]
- Gazith, J.; Cavey, M.T.; Cavey, D.; Shroot, B.; Reichert, U. Characterization of the beta-adrenergic receptors of cultured human epidermal keratinocytes. Biochem. Pharm. 1983, 32, 3397–3403. [Google Scholar] [CrossRef]
- Steinkraus, V.; Mak, J.C.; Pichlmeier, U.; Mensing, H.; Ring, J.; Barnes, P.J. Autoradiographic mapping of beta-adrenoceptors in human skin. Arch. Dermatol. Res. 1996, 288, 549–553. [Google Scholar] [CrossRef]
- Wijaya, L.K.; Stumbles, P.A.; Drummond, P.D. Tumor necrosis factor α induces α1B-adrenergic receptor expression in keratinocytes. Cytokine 2020, 125, 154851. [Google Scholar] [CrossRef]
- Howell, R.E.; Albelda, S.M.; Daise, M.L.; Levine, E.M. Characterization of beta-adrenergic receptors in cultured human and bovine endothelial cells. J. Appl. Physiol. 1988, 65, 1251–1257. [Google Scholar] [CrossRef]
- Steinberg, S.F.; Jaffe, E.A.; Bilezikian, J.P. Endothelial cells contain beta adrenoceptors. Naunyn-Schmiedeberg’s Arch. Pharm. 1984, 325, 310–313. [Google Scholar] [CrossRef] [PubMed]
- De Coupade, C.; Gear, R.W.; Dazin, P.F.; Sroussi, H.Y.; Green, P.G.; Levine, J.D. Beta 2-adrenergic receptor regulation of human neutrophil function is sexually dimorphic. Br. J. Pharmacol. 2004, 143, 1033–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeckx, K.C.; Doornbos, R.P.; Witkamp, R.F.; van der Greef, J.; Rodenburg, R.J. Beta-adrenergic receptor agonists induce the release of granulocyte chemotactic protein-2, oncostatin M, and vascular endothelial growth factor from macrophages. Int Immunopharmacol. 2006, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Berrettini, W.H.; Bardakjian, J.; Cappellari, C.B.; Barnett, A.L., Jr.; Albright, A.; Nurnberger, J.I., Jr.; Gershon, E.S. Skin fibroblast beta-adrenergic receptor function in manic-depressive illness. Biol. Psychiatry 1987, 22, 1439–1443. [Google Scholar] [CrossRef]
- Furlan, C.; Sterin-Borda, L.; Borda, E. Activation of beta3 adrenergic receptor decreases DNA synthesis in human skin fibroblasts via cyclic GMP/nitric oxide pathway. Cell Physiol. Biochem. 2005, 16, 175–182. [Google Scholar] [CrossRef]
- Huang, H.; Gibson, G. Altered beta-adrenergic receptor-stimulated cAMP formation in cultured skin fibroblasts from Alzheimer donors. J. Biol. Chem. 1993, 268, 14616–14621. [Google Scholar] [CrossRef]
- Liao, M.-H.; Liu, S.-S.; Peng, I.-C.; Tsai, F.-J.; Huang, H.H. The stimulatory effects of alpha1-adrenergic receptors on TGF-beta1, IGF-1 and hyaluronan production in human skin fibroblasts. Cell Tissue Res. 2014, 357, 681–693. [Google Scholar] [CrossRef]
- Pullar, C.E.; Isseroff, R.R. The beta 2-adrenergic receptor activates pro-migratory and pro-proliferative pathways in dermal fibroblasts via divergent mechanisms. J. Cell Sci. 2006, 119, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Sterin-Borda, L.; Furlan, C.; Orman, B.; Borda, E. Differential regulation on human skin fibroblast by α1 adrenergic receptor subtypes. Biochem. Pharm. 2007, 74, 1401–1412. [Google Scholar] [CrossRef]
- Pullar, C.E.; Rizzo, A.; Isseroff, R.R. beta-Adrenergic receptor antagonists accelerate skin wound healing: Evidence for a catecholamine synthesis network in the epidermis. J. Biol. Chem. 2006, 281, 21225–21235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koizumi, H.; Yasui, C.; Fukaya, T.; Ohkawara, A.; Ueda, T. Beta-Adrenergic Stimulation Induces Intracellular Ca++ Increase in Human Epidermal Keratinocytes. J. Investig. Derm. 1991, 96, 234–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munaron, L.; Antoniotti, S.; Lovisolo, D. Intracellular calcium signals and control of cell proliferation: How many mechanisms? J. Cell. Mol. Med. 2004, 8, 161–168. [Google Scholar] [CrossRef]
- Pinto, M.C.X.; Kihara, A.H.; Goulart, V.A.; Tonelli, F.M.; Gomes, K.N.; Ulrich, H.; Resende, R.R. Calcium signaling and cell proliferation. Cell. Signal. 2015, 27, 2139–2149. [Google Scholar] [CrossRef] [PubMed]
- Romana-Souza, B.; Porto, L.C.; Monte-Alto-Costa, A. Cutaneous wound healing of chronically stressed mice is improved through catecholamines blockade. Exp. Derm. 2010, 19, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Pullar, C.E.; Chen, J.; Isseroff, R.R. PP2A activation by beta2-adrenergic receptor agonists: Novel regulatory mechanism of keratinocyte migration. J Biol. Chem. 2003, 278, 22555–22562. [Google Scholar] [CrossRef] [Green Version]
- Pullar, C.E.; Isseroff, R.R. Cyclic AMP mediates keratinocyte directional migration in an electric field. J. Cell Sci. 2005, 118, 2023–2034. [Google Scholar] [CrossRef] [Green Version]
- Kavelaars, A. Regulated expression of α-1 adrenergic receptors in the immune system. Brain Behav. Immun. 2002, 16, 799–807. [Google Scholar] [CrossRef]
- Kohm, A.P.; Sanders, V.M. Norepinephrine and beta 2-adrenergic receptor stimulation regulate CD4+ T and B lymphocyte function in vitro and in vivo. Pharmacol. Rev. 2001, 53, 487–525. [Google Scholar]
- Kim, M.H.; Gorouhi, F.; Ramirez, S.; Granick, J.L.; Byrne, B.A.; Soulika, A.M.; Simon, S.I.; Isseroff, R.R. Catecholamine stress alters neutrophil trafficking and impairs wound healing by beta2-adrenergic receptor-mediated upregulation of IL-6. J. Investig. Dermatol. 2014, 134, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Lorton, D.; Bellinger, D.L. Molecular mechanisms underlying beta-adrenergic receptor-mediated cross-talk between sympathetic neurons and immune cells. Int. J. Mol. Sci. 2015, 16, 5635–5665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romana-Souza, B.; Otranto, M.; Almeida, T.F.; Porto, L.C.; Monte-Alto-Costa, A. Stress-induced epinephrine levels compromise murine dermal fibroblast activity through beta-adrenoceptors. Exp. Dermatol. 2011, 20, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Chotani, M.A.; Flavahan, S.; Mitra, S.; Daunt, D.; Flavahan, N.A. Silent alpha(2C)-adrenergic receptors enable cold-induced vasoconstriction in cutaneous arteries. Am. J. Physiol. Circ. Physiol. 2000, 278, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Ekenvall, L.; E Lindblad, L.; Norbeck, O.; Etzell, B.M. alpha-Adrenoceptors and cold-induced vasoconstriction in human finger skin. Am. J. Physiol. Content 1988, 255, 1000–1003. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, A.P.; Fox, J.M.; Pullar, C.E. Beta-Adrenoceptor Activation Reduces Both Dermal Microvascular Endothelial Cell Migration via a cAMP-Dependent Mechanism and Wound Angiogenesis. J. Cell Physiol. 2015, 230, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Götz, F.; Bannerman, T.; Schleifer, K.H. The Genera Staphylococcus and Macrococcus. In Procaryotes; Dworkin, M., Ed.; Springer: New York, NY, USA, 2006; pp. 5–75. [Google Scholar]
- Luqman, A.; Nega, M.; Nguyen, M.T.; Ebner, P.; Götz, F. SadA-Expressing Staphylococci in the Human Gut Show Increased Cell Adherence and Internalization. Cell Rep. 2018, 22, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Luqman, A.; Ebner, P.; Reichert, S.; Sass, P.; Kabagema-Bilan, C.; Heilmann, C.; Ruth, P.; Götz, F. A new host cell internalisation pathway for SadA-expressing staphylococci triggered by excreted neurochemicals. Cell Microbiol. 2019, 21, e13044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Bavadekar, S.A.; Schaneberg, B.T.; Khan, I.A.; Feller, D.R. Effects of synephrine and beta-phenethylamine on human alpha-adrenoceptor subtypes. Planta Med. 2010, 76, 981–986. [Google Scholar] [CrossRef]
- Kleinau, G.; Pratzka, J.; Nürnberg, D.; Grüters, A.; Führer-Sakel, D.; Krude, H.; Köhrle, J.; Schöneberg, T.; Biebermann, H. Differential Modulation of Beta-Adrenergic Receptor Signaling by Trace Amine-Associated Receptor 1 Agonists. PLoS ONE 2011, 6, e27073. [Google Scholar] [CrossRef]
- Luqman, A.; Kharisma, V.D.; Ruiz, R.A.; Götz, F. In Silico and In Vitro Study of Trace Amines (TA) and Dopamine (DOP) Interaction with Human Alpha 1-Adrenergic Receptor and the Bacterial Adrenergic Receptor QseC. Cell. Physiol. Biochem. 2020, 54, 888–898. [Google Scholar] [CrossRef]
- Steenhuis, P.; Huntley, R.E.; Gurenko, Z.; Yin, L.; Dale, B.A.; Fazel, N.; Isseroff, R.R. Adrenergic Signaling in Human Oral Keratinocytes and Wound Repair. J. Dent. Res. 2010, 90, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Borrel, V.; Thomas, P.; Catovic, C.; Racine, P.-J.; Konto-Ghiorghi, Y.; Lefeuvre, L.; Duclairoir-Poc, C.; Zouboulis, C.C.; Feuilloley, M.G.J. Acne and Stress: Impact of Catecholamines on Cutibacterium acnes. Front. Med. 2019, 6, 155. [Google Scholar] [CrossRef] [PubMed]
- Danilova, N.D.; Solovyeva, T.V.; Mart’Yanov, S.V.; Zhurina, M.V.; Gannesen, A.V. Stimulatory Effect of Epinephrine on Biofilms of Micrococcus luteus C01. Microbiology 2020, 89, 493–497. [Google Scholar] [CrossRef]
- Bearson, B.L.; Bearson, S.M.; Lee, I.S.; Brunelle, B.W. The Salmonella enterica serovar Typhimurium QseB response regulator negatively regulates bacterial motility and swine colonization in the absence of the QseC sensor kinase. Microb. Pathog. 2010, 48, 214–219. [Google Scholar] [CrossRef]
- Clarke, M.B.; Hughes, D.T.; Zhu, C.; Boedeker, E.C.; Sperandio, V. The QseC sensor kinase: A bacterial adrenergic receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 10420–10425. [Google Scholar] [CrossRef] [Green Version]
- Lesouhaitier, O.; Véron, W.; Chapalain, A.; Madi, A.; Blier, A.-S.; Dagorn, A.; Connil, N.; Chevalier, S.; Orange, N.; Feuilloley, M. Gram-Negative Bacterial Sensors for Eukaryotic Signal Molecules. Sensors 2009, 9, 6967–6990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njoroge, J.; Sperandio, V. Enterohemorrhagic Escherichia coli Virulence Regulation by Two Bacterial Adrenergic Kinases, QseC and QseE. Infect. Immun. 2011, 80, 688–703. [Google Scholar] [CrossRef] [Green Version]
- Weigel, W.A.; DeMuth, D.R.; Qse, B.C. A two-component bacterial adrenergic receptor and global regulator of virulence in Enterobacteriaceae and Pasteurellaceae. Mol. Oral Microbiol. 2016, 31, 379–397. [Google Scholar] [CrossRef] [Green Version]
- Cambronel, M.; Nilly, F.; Mesguida, O.; Boukerb, A.M.; Racine, P.-J.; Baccouri, O.; Borrel, V.; Martel, J.; Fécamp, F.; Knowlton, R.; et al. Influence of Catecholamines (Epinephrine/Norepinephrine) on Biofilm Formation and Adhesion in Pathogenic and Probiotic Strains of Enterococcus faecalis. Front. Microbiol. 2020, 11, 1501. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, S.M.; Shergill, R.; Woodward, J.; Muralikuttan, R.; Haigh, R.D.; Lyte, M.; Freestone, P.P. Elucidation of the Mechanism by Which Catecholamine Stress Hormones Liberate Iron from the Innate Immune Defense Proteins Transferrin and Lactoferrin. J. Bacteriol. 2009, 192, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratford, A.F.; Zoutman, D.E.; Davidson, J.S. Effect of lidocaine and epinephrine on Staphylococcus aureus in a guinea pig model of surgical wound infection. Plast. Reconstr. Surg. 2002, 110, 1275–1279. [Google Scholar] [CrossRef]
- Biaggini, K.; Barbey, C.; Borrel, V.; Feuilloley, M.; Déchelotte, P.; Connil, N. The pathogenic potential of Pseudomonas fluorescens MFN1032 on enterocytes can be modulated by serotonin, substance P and epinephrine. Arch. Microbiol. 2015, 197, 983–990. [Google Scholar] [CrossRef]
- Wu, M.; Li, Y.; Guo, D.; Kui, G.; Li, B.; Deng, Y.; Li, F. Microbial Diversity of Chronic Wound and Successful Management of Traditional Chinese Medicine. Evid. Based Complementary Altern. Med. 2018, 2018, 9463295. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Effector | Mode of Action | References |
|---|---|---|---|
| (A) Adverse effect on wound healing | |||
| Peptostreptococcus spp. (P. magnus, P. vaginalis, and P. asaccharolyticus) | Bacterial supernatant | Inhibit fibroblast proliferation, keratinocyte proliferation and repopulation, and endothelial tubule formation | [17] |
| Staphylococcus aureus, Pseudomonas aeruginosa, Peptoniphilus sp., Stenotrophomonas sp., Finegoldia sp., Serratia sp., Bacillus sp., Enterococcus sp., Enterobacter aerogenes, Acinetobacter baumanii, Klebsiella pneumoniae, Proteus mirabilis, Aspergillus fumigatus, Enterobacter cloacae, Corynebacterium frankenforstense, Corynebacterium striatum, Alcaligenes faecalis and Acinetobacter sp. | Biofilm | Related to wound chronicity | [12,13,39,40] |
| Skin commensals | Can be particulate cell wall peptidoglycan | Augment S. aureus pathogenesis | [41] |
| Bacteria | Muramyl dipeptide | Delay wound repair by reducing re-epithelialization; increasing inflammation; and upregulating of murine β-defensins 1, 3, and 14 | [42] |
| Pathogenic staphylococci | Secreted factors | Activate the mitogen-activated protein kinase and phosphatidylinositol 3-kinase/AKT signaling pathways and suppress NF-κB activation | [28] |
| Pathogenic fungi | Wound necrosis | [43] | |
| (B) Promoting effect on wound healing | |||
| Staphylococcus epidermidis and possibly other skin commensals with sadA gene | Trace amines | Accelerate wound healing by partially antagonizing the β-adrenergic receptor | [19,44] |
| Skin commensals predominantly from Staphylococcus | Bacteriocins | Inhibit pathogenic Gram-positive bacteria such as Cutibacterium acnes, Staphylococcus epidermidis, and MRSA | [45] |
| Staphylococcus caprae and other coagulase-negative staphylococci | Autoinducing peptide | Inhibit quorum sensing of S. aureus | [31,46] |
| Lactobacilli | Organic acids | Antimicrobial activity against skin pathogens and prevent biofilm formation | [37,47,48] |
| Lactobacillus rhamnosus, Lactobacillus reuteri | Lysate | Increase keratinocyte proliferation and migration | [49] |
| S. epidermidis | Short chain fatty acids | Suppress the growth of S. aureus and C. acnes | [50] |
| S. epidermidis | Delta-toxin (PSMγ) | Cooperates with the host-derived antimicrobial peptides in the innate immune system to eliminate pathogens | [51] |
| S. epidermidis | Induces IL-17A+ CD8+ T cells, enhances innate barrier immunity, and limits pathogen invasion | [9] | |
| S. epidermidis and S. hominis | Antimicrobial peptides | Selectively kill S. aureus and synergize with the human AMP LL-37 | [36] |
| Commensal staphylococci | Secreted factors | Induce expression of the AMPs HBD-3 and RNase7 in primary human keratinocytes via Toll-like receptor (TLR)-2, EGFR, and NF-κB activation | [28] |
| Commensal staphylococci | Lipoteichoic acid | Inhibit both inflammatory cytokine release from keratinocytes and inflammation triggered by injury through a TLR2-dependent mechanism | [52] |
| S. aureus | Peptidoglycan | Ameliorate cyclophosphamide-impaired wound healing | [53] |
| Staphylococci | Surface proteins SpA and Sbi | Initiate signaling cascades that lead to the early recruitment of neutrophils, modulate their lifespan in the skin milieu, and contribute to proper abscess formation and bacterial eradication | [30] |
| Skin commensals | Trigger activation of neutrophils to express the chemokine CXCL10 to kill exposed microbiota; activate pDC to produce type I IFNs, which accelerate wound closure by triggering skin inflammation and early T-cell-independent wound repair responses | [29] | |
| Skin commensals | Induce T-cell responses that lead to protection from pathogens and accelerated skin wound closure | [54] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luqman, A.; Götz, F. The Ambivalent Role of Skin Microbiota and Adrenaline in Wound Healing and the Interplay between Them. Int. J. Mol. Sci. 2021, 22, 4996. https://doi.org/10.3390/ijms22094996
Luqman A, Götz F. The Ambivalent Role of Skin Microbiota and Adrenaline in Wound Healing and the Interplay between Them. International Journal of Molecular Sciences. 2021; 22(9):4996. https://doi.org/10.3390/ijms22094996
Chicago/Turabian StyleLuqman, Arif, and Friedrich Götz. 2021. "The Ambivalent Role of Skin Microbiota and Adrenaline in Wound Healing and the Interplay between Them" International Journal of Molecular Sciences 22, no. 9: 4996. https://doi.org/10.3390/ijms22094996