
Repeated Irradiation with γ-Ray Induces Cancer Stemness through TGF-β-DLX2 Signaling in the A549 Human Lung Cancer Cell Line


Abstract
:1. Introduction
2. Results
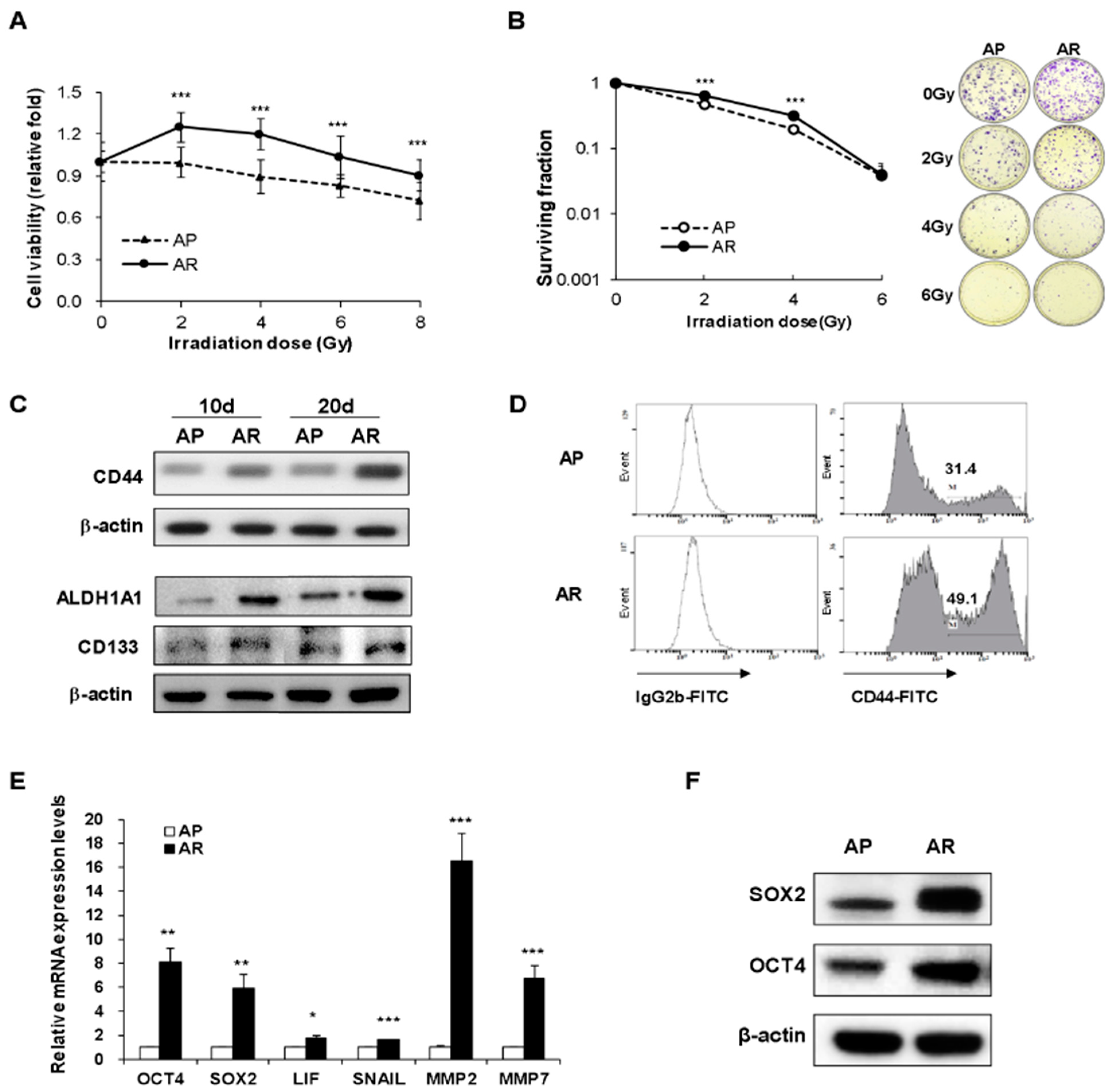
2.1. Surviving A549 Cells after Fractionated Irradiation Acquire Radioresistance and Stemness
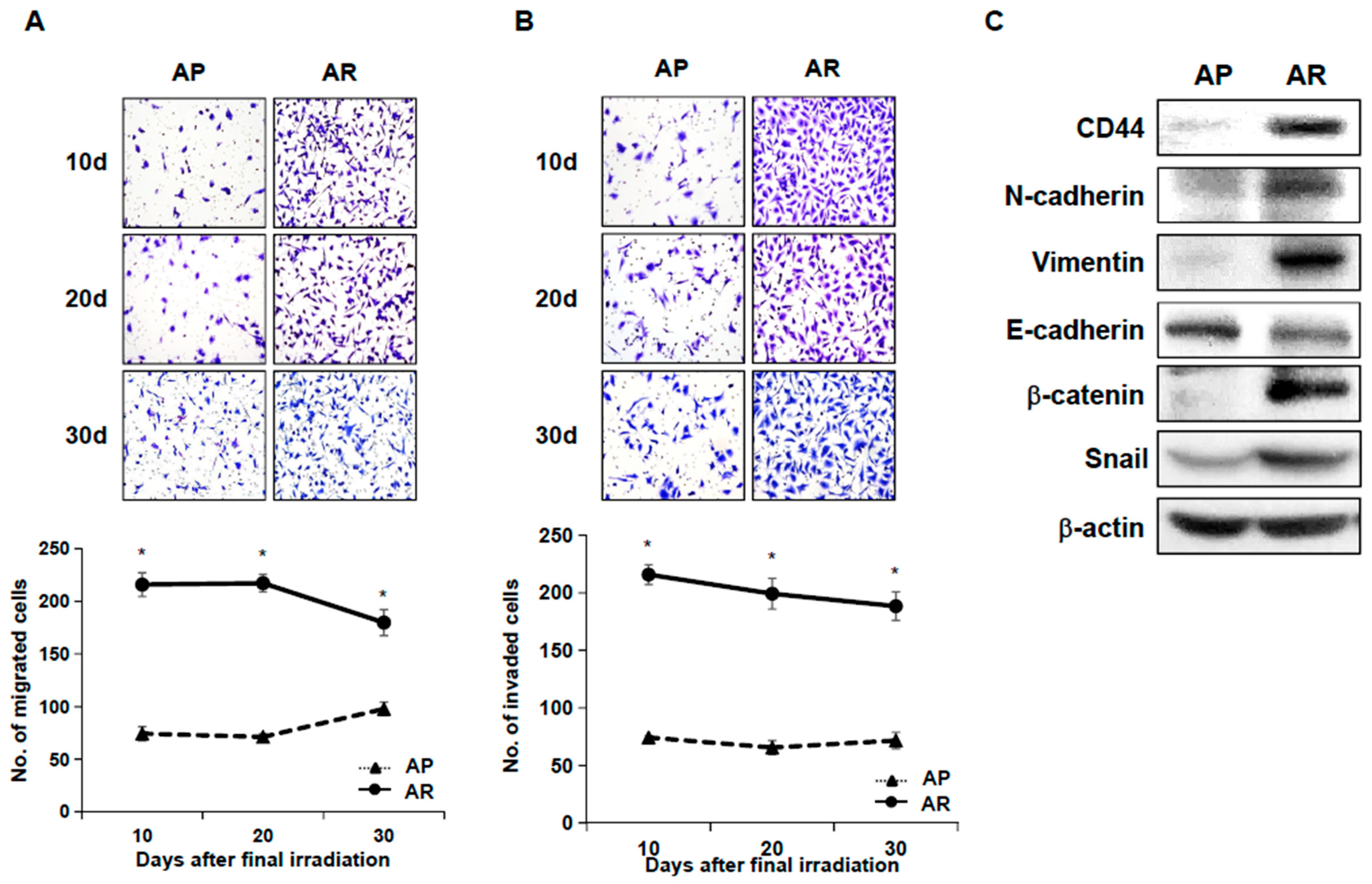
2.2. Fractionated Ionizing Radiation Enhances Migration and Invasion of A549 Cells
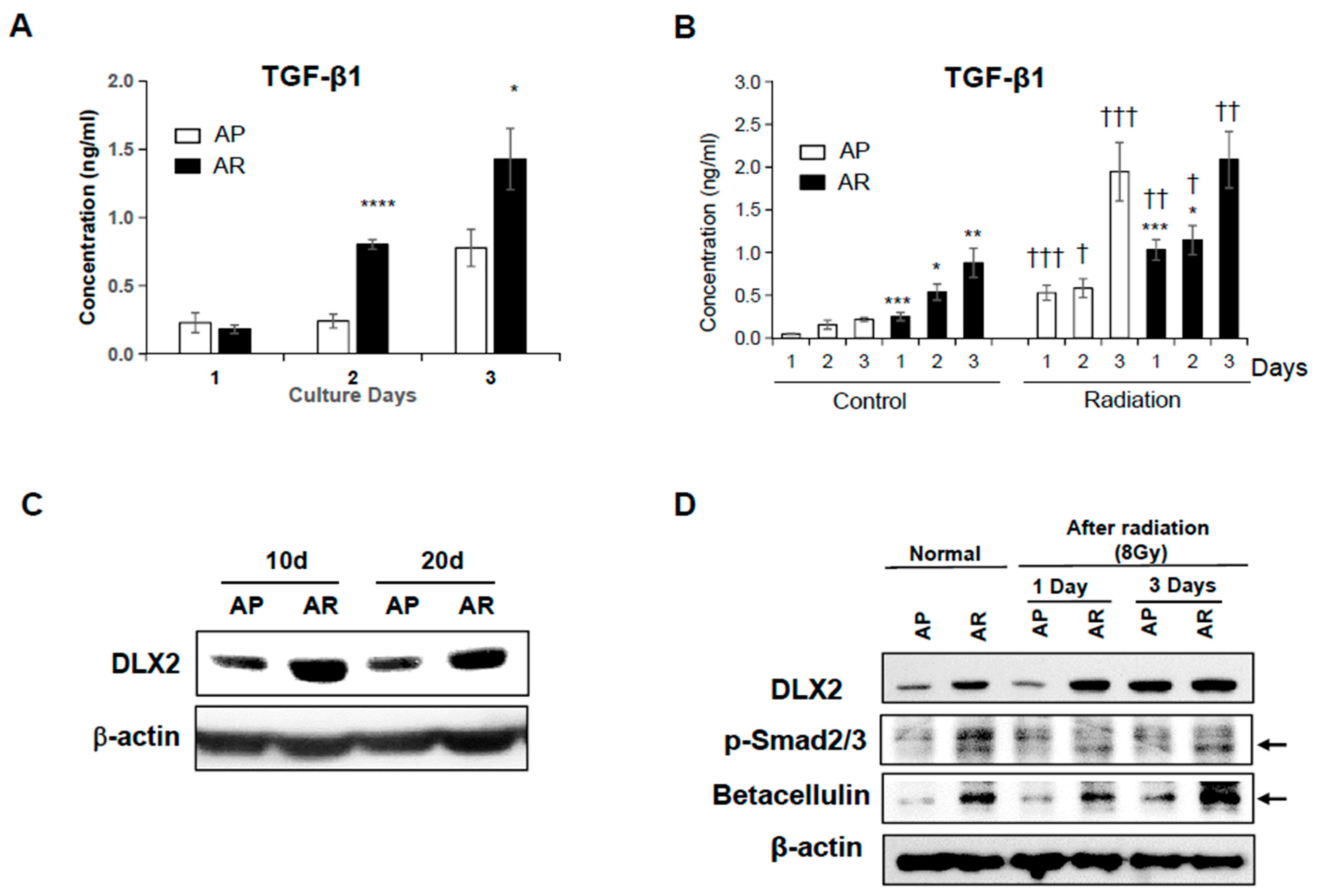
2.3. Fractionated Ionizing Radiation Induces Radioresistance through TGF-β-Smad2/3-DLX2 Signaling
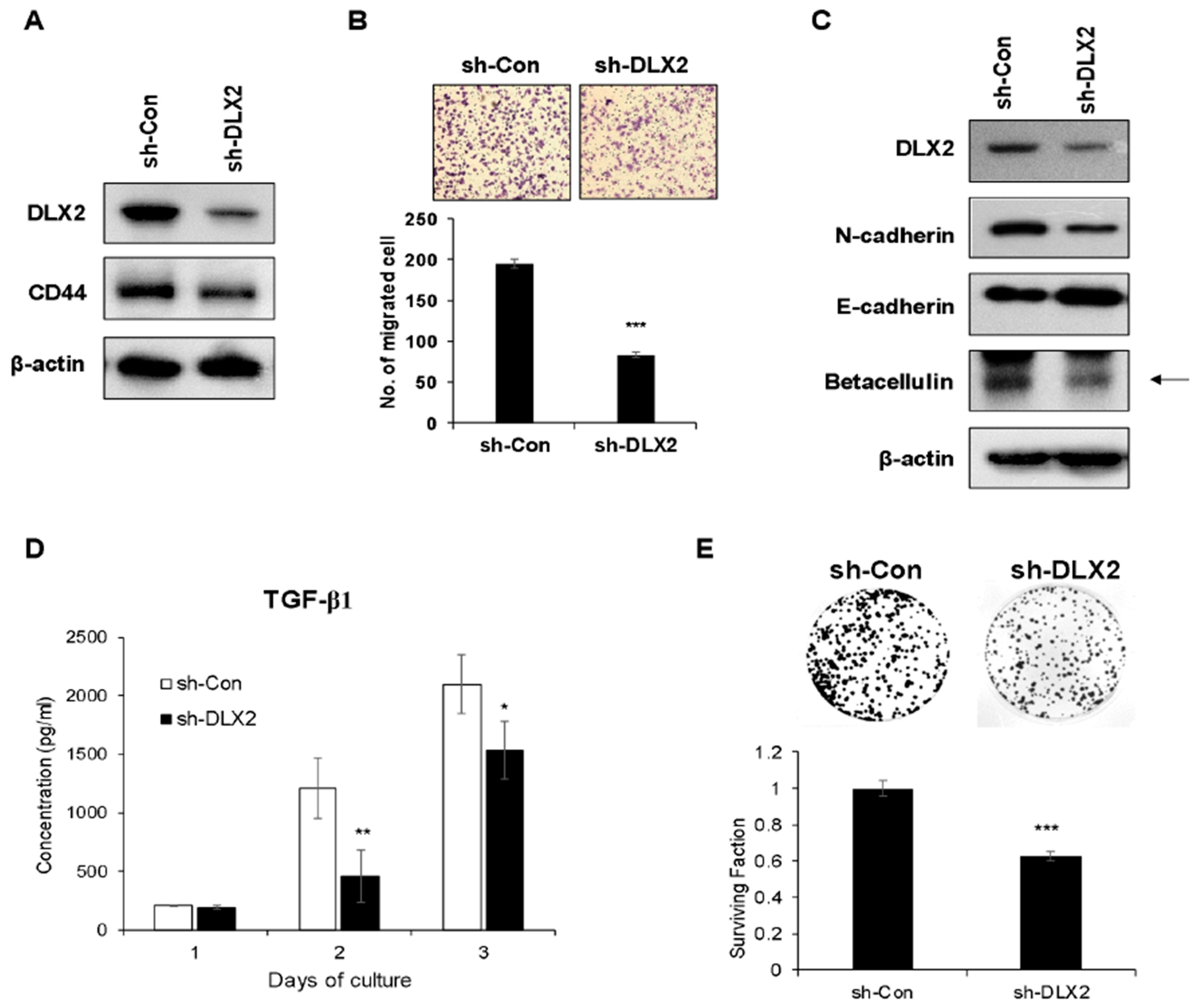
2.4. Knockdown of DLX2 Attenuates the Characteristic of Cancer Stemness and Cancer Cell Proliferation In Vitro
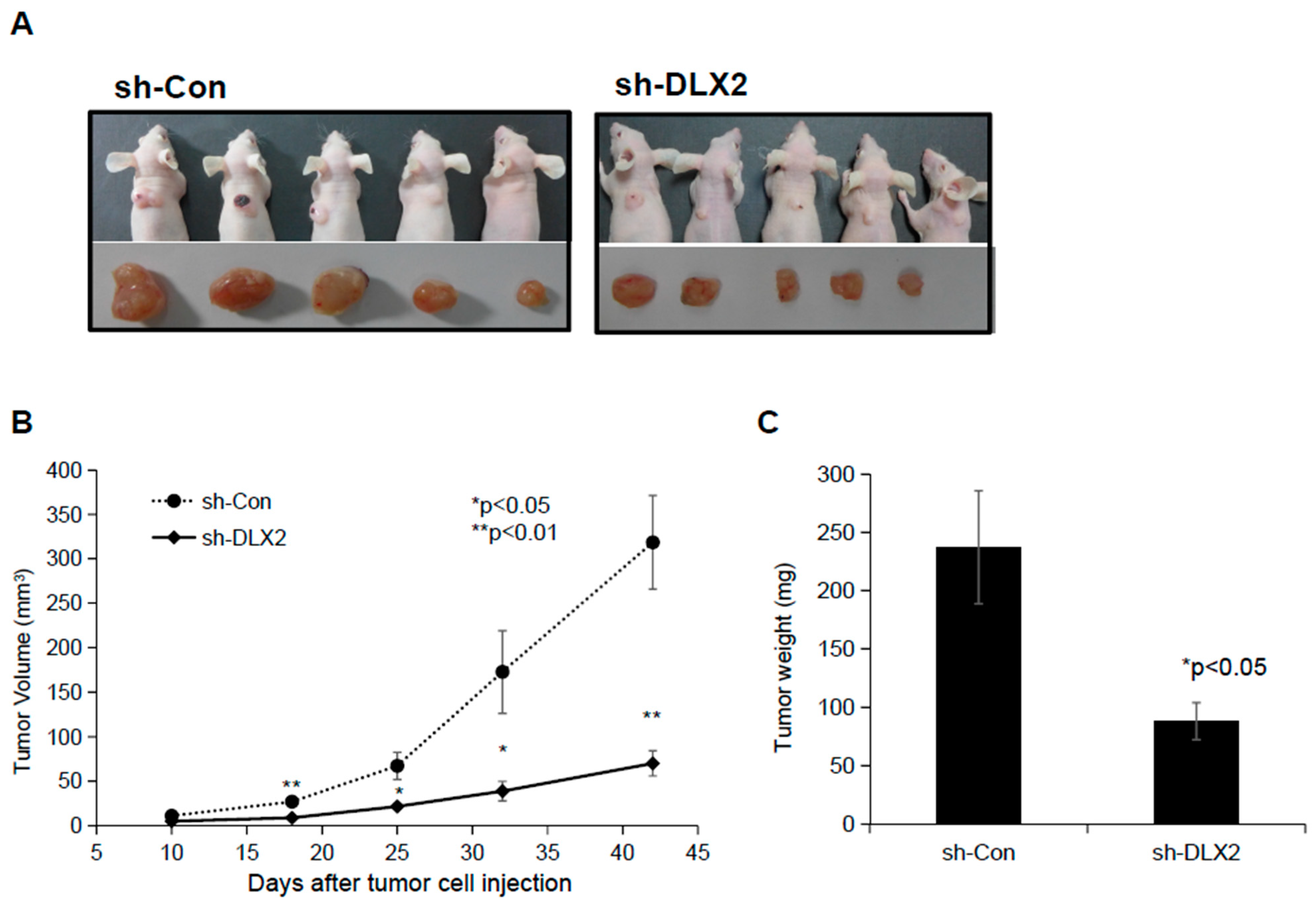
2.5. Knockdown of DLX2 Results in a Reduction of Tumor Growth
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Irradiation
4.2. Cell Counting Kit-8 (CCK-8) Assay for Cell Viability
4.3. Colony Forming Assay
4.4. Cell Migration Assay
4.5. Cell Invasion Assay
4.6. Western Blot Analysis
4.7. Flow Cytometry Analysis
4.8. RNA Extraction and Quantitative Real-Time PCR
4.9. Measurement of TGF-β1 in Cell Culture Supernatant
4.10. Establishment of shRNA-Based DLX2 Knockdown A549 Cells
4.11. Xenograft Tumor Growth Assay
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hubenak, J.R.; Zhang, Q.; Branch, C.D.; Kronowitz, S.J. Mechanisms of Injury to Normal Tissue after Radiotherapy: A Review. Plast. Reconstr. Surg. 2014, 133, 49e–56e. [Google Scholar] [CrossRef] [Green Version]
- De Bacco, F.; Luraghi, P.; Medico, E.; Reato, G.; Girolami, F.; Perera, T.; Gabriele, P.; Comoglio, P.M.; Boccaccio, C. Induction of MET by Ionizing Radiation and Its Role in Radioresistance and Invasive Growth of Cancer. J. Natl. Cancer Inst. 2011, 103, 645–661. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, X.; Zhang, N.; Yang, Q.; Moran, M.S. Low Doses Ionizing Radiation Enhances the Invasiveness of Breast Cancer Cells by Inducing Epithelial-mesenchymal Transition. Biochem. Biophys. Res. Commun. 2011, 412, 188–192. [Google Scholar] [CrossRef]
- Park, J.K.; Jang, S.J.; Kang, S.W.; Park, S.; Hwang, S.-G.; Kim, W.-J.; Kang, J.H.; Um, H.-D. Establishment of Animal Model for the Analysis of Cancer Cell Metastasis during Radiotherapy. Radiat. Oncol. 2012, 7, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glarke, M.F.; Fuller, M. Stem Cells and Cancer: Two Faces of Eve. Cell 2006, 124, 1111–1115. [Google Scholar]
- Baumann, M.; Krause, M.; Hill, R. Exploring the Role of Cancer Stem Cells in Radioresistance. Nat. Rev. Cancer 2008, 8, 545–554. [Google Scholar] [CrossRef]
- Nguyen, L.V.; Vanner, R.; Dirks, P.; Eaves, C.J. Cancer Stem Cells: An Evolving Concept. Nat. Rev. Cancer 2012, 12, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, I.; Trumpp, A. The Evolving Concept of Cancer and Metastasis Stem Cells. J. Cell Biol. 2012, 198, 281–293. [Google Scholar] [CrossRef]
- Tirino, V.; Desiderio, V.; Paino, F.; De Rosa, A.; Papaccio, F.; La Noce, M.; Laino, L.; De Francesco, F.; Papaccio, G. Cancer Stem Cells in Solid Tumors: An Overview and New Approaches for Their Isolation and Characterization. FASEB J. 2013, 27, 13–24. [Google Scholar] [CrossRef]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A Cell Initiating Human Acute Myeloid Leukemia after Transplantation into SCID Mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective Identification of Tumorigenic Breast Cancer Cell. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of Human Brain Tumour Initiating Cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Li, C.; Heidt, D.G.; Dalerba, P.; Burant, C.F.; Zhang, L.; Adsay, V.; Wicha, M.; Clarke, M.F.; Simeone, D.M. Identification of Pancreatic Cancer Stem Cells. Cancer Res. 2007, 67, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Ricci-Vitiani, L.; Lombardi, D.G.; Pilozzi, E.; Biffoni, M.; Todaro, M.; Peschle, C.; De Maria, R. Identification and Expansion of Human Colon-cancer-initiating Cells. Nature 2007, 445, 111–115. [Google Scholar] [CrossRef]
- Eramo, A.; Lotti, F.; Sette, G.; Pilozzi, E.; Biffoni, M.; Di Virgilio, A.; Conticello, C.; Ruco, L.; Peschle, C.; De Maria, R. Identification and Expansion of the Tumorigenic Lung Cancer Stem Cell Population. Cell Death Differ. 2008, 15, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.; Krause, M. CD44: A Cancer Stem Cell-related Biomarker with Predictive Potential for Radiotherapy. Clin. Cancer Res. 2010, 16, 5091–5093. [Google Scholar] [CrossRef] [Green Version]
- Vlashi, E.; Pajonk, F. Cancer Stem Cells, Cancer Cell Plasticity and Radiation Therapy. Semin. Cancer Biol. 2015, 31, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosse-Gehling, P.; Fargeas, C.A.; Dittfeld, C.; Garbe, Y.; Alison, M.R.; Corbeil, D.; Kunz-Schughart, L.A. CD133 as a Biomarker for Putative Cancer Stem Cells in Solid Tumours: Limitations, Problems and Challenges. J. Pathol. 2013, 229, 355–378. [Google Scholar] [CrossRef]
- Jaggupilli, A.; Elkord, E. Significance of C44 and CD24 as Cancer Stem Cell Markers: An Enduring Ambiguity. Clin. Dev. Immunol. 2012, 2012, 708036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokrowiecka, A.; Veits, L.; Fakleis, C.; Musial, J.; Kordek, R.; Lochowski, M.; Kozak, J.; Wierzchniewska-Lawska, A.; Vieth, M.; Malecka-Panas, E. Expression Profiles of Cancer Stem Cell Markers: CD133, CD44, Musashi-1 and EpCAM in the Cardiac Mucosa-Barrett’s Esophagus-early Esophageal Adenocarcinoma-advanced Esophageal Adenocarcinoma Sequence. Pathol. Res. Pract. 2017, 213, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Ma, I.; Allan, A.I. The Role of Human Aldehyde Dehydrogenase in Normal and Cancer Stem Cells. Stem Cell Rev. 2011, 7, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Prud’homme, G.J. Cancer Stem Cells and Novel Targets for Antitumor Strategies. Curr. Pharm. Des. 2012, 18, 2838–2849. [Google Scholar] [CrossRef]
- Ghisolfi, L.; Keates, A.C.; Hu, X.; Lee, D.K.; Li, C.J. Ionizing Radiation Induces Stemness in Cancer Cells. PLoS ONE 2012, 7, e43628. [Google Scholar] [CrossRef] [PubMed]
- Peitzsch, C.; Kurth, I.; Kunz-Schughart, L.; Baumann, M.; Dubrovska, A. Discovery of the Cancer Stem Cell Related Determinants of Radioresistance. Radiothera. Oncol. 2013, 108, 378–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Casal, R.; Bhattacharya, C.; Ganesh, N.; Bailey, L.; Basse, P.; Gibson, M.; Epperly, M.; Levina, V. Non-small Cell Lung Cancer Cells Survived Ionizing Radiation Treatment Display Cancer Stem Cell and Epithelial-mesenchymal Transition Phenotypes. Mol. Cancer 2013, 12, 94. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Jeong, E.K.; Ju, M.K.; Jeon, H.M.; Kim, M.Y.; Kim, C.H.; Park, G.H.; Han, S.I.; Kang, H.S. Induction of Metastasis, Cancer Stem Cell Phenotype, and Oncogenic Metabolism in Cancer Cells by Ionizing Radiation. Mol. Cancer 2017, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Martins-Neves, S.R.; Gleton-Jansen, A.M.; Gomes, C.M.F. Therapy-induced Enrichment of Cancer Stem-like Cells in Solid Human Tumors: Where Do We Stand? Pharmacol. Res. 2018, 137, 193–204. [Google Scholar] [CrossRef]
- Christianson, J.J.; Rajasekaran, A.K. Reassessing Epithelial to Mesenchymal Transition as a Prerequisite for Carcinoma Invasion and Metastasis. Cancer Res. 2006, 66, 8319–8326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-mesenchymal Transition in Cancer: Parallels between Normal Development and Tumor Progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Scheel, C.; Weinberg, R.A. Cancer Stem Cells and Epithelial-mesenchymal Transition: Concepts and Molecular Links. Semin. Cancer Biol. 2012, 22, 396–403. [Google Scholar] [CrossRef]
- De Craene, B.; Berx, G. Regulatory Networks Defining EMT during Cancer Initiation and Progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef]
- Puisieux, A.; Brabletz, T.; Caramel, J. Oncogenic Roles of EMT-inducing Transcription Factors. Nat. Cell Biol. 2014, 16, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Kang, Y. Multilayer Control of the EMT Master Regulators. Oncogene 2014, 33, 1755–1763. [Google Scholar] [CrossRef]
- Siegel, P.M.; Massague, J. Cytostatic and Apoptotic Actions of TGF-beta in Homeostasis and Cancer. Nat. Rev. Cancer 2003, 3, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Massague, J. TGFbeta in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakefield, L.M.; Roberts, A.B. TGF-beta Signaling: Positive and Negative Effects on Tumorigenesis. Curr. Opin. Genet. Dev. 2002, 12, 22–29. [Google Scholar] [CrossRef]
- Pardali, K.; Moustakas, A. Actions of TGF-beta as Tumor Suppressor and Pro-metastatic Factor in Human Cancer. Biochem. Biophys. Acta 2007, 1775, 21–62. [Google Scholar]
- Choi, Y.J.; Baek, G.Y.; Park, H.R.; Jo, S.K.; Jung, U. Smad2/3-regulated Expression of DLX2 is Associated with Radiation-induced Epithelial-mesenchymal Transition and Radioresistance of A549 and MDA-MB-231 Human Cancer Cell Lines. PLoS ONE 2016, 11, e0147343. [Google Scholar] [CrossRef] [Green Version]
- Merlo, G.R.; Zerega, B.; Paleari, L.; Trombino, S.; Mantero, S.; Levi, G. Multiple Functions of DLX Genes. Int. J. Dev. Biol. 2000, 44, 619–626. [Google Scholar]
- Panganiban, G.; Rubenstein, J.L. Developmental Functions of the Distal-less/Dlx Homeobox Genes. Development 2002, 129, 4371–4386. [Google Scholar]
- Morini, M.; Astigiano, S.; Gitton, Y.; Emionite, L.; Mirisola, V.; Levi, G.; Barbieri, O. Mutually Exclusive Expression of DLX2 and DLX5/6 is Associated with the Metastatic Potential of the Human Breast Cancer Cell Line MDA-MB-231. BMC Cancer 2010, 10, 649. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, M.; Maaβ, D.; Tiwari, N.; Waldmeier, L.; Schmidt, P.; Lehembre, F.; Christofori, G. Transcription Factor Dlx2 Protects from TGFβ-induced cell-cycle Arrest and Apoptosis. EMBO J. 2011, 30, 4489–4499. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.; Huang, H.; Chang, J.; Zhao, G.F.; Lu, M.L.; Wang, Y. Increased Expression of DLX2 Correlates with Advanced Stage of Gastric Adenocarcinoma. World J. Gastroenterol. 2013, 19, 2697–2703. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Jeon, H.M.; Ju, M.K.; Jeong, E.K.; Kim, C.H.; Yoo, M.; Park, H.G.; Han, S.I.; Kang, H.S. Dlx2 is Implicated in TGF-β- and Wnt-induced Epithelial-mesenchymal, Glycolytic Switch, and Mitochondrial Repression by Snail Activation. Int. J. Oncol. 2015, 46, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Deryneck, R. Molecular Mechanisms of Epithelial-mesenchymal Transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Weinberg, R.A. Epithelial-mesenchymal Transition: At the Cross Roads of Development and Tumor Metastasis. Dev. Cell 2008, 14, 818–829. [Google Scholar] [CrossRef] [Green Version]
- Massague, J. G1 Cell-cycle Control and Cancer. Nature 2004, 432, 298–306. [Google Scholar] [CrossRef]
- Siegel, R.; Miller, K.D.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Sishc, B.J.; Nelson, C.B.; Hahnfeldt, P.; Bailey, S.M.; Hlatky, L. Radiation-induced Reprogramming of Pre-senescent Mammary Epithelial Cells Enriches Putative CD44+/CD24-/low Stem Cell Phenotype. Front. Oncol. 2016, 6, 138. [Google Scholar] [CrossRef] [Green Version]
- Moncharmont, C.; Levy, A.; Guy, J.B.; Falk, A.T.; Guilbert, M.; Trone, J.C.; Alphonse, G.; Gilormini, M.; Ardail, D.; Magné, N.; et al. Radiation-enhanced Cell Migration/Invasion Process: A Review. Crit. Rev. Oncol. Hematol. 2014, 92, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, A.; Yokoe, T.; Tanaka, K.; Saigusa, S.; Toiyama, Y.; Yasuda, H.; Inoue, Y.; Miki, C.; Kusunoki, M. Radiation Induces Epithelial-mesenchymal Transition in Colorectal Cancer Cells. Oncol. Rep. 2012, 27, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dancea, H.C.; Shareef, M.M.; Ahmed, M.M. Role of Radiation-induced TGF-beta Signaling in Cancer Therapy. Mol. Cell Pharmacol. 2009, 1, 44–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carl, C.; Flindt, A.; Hartmann, J.; Dahlke, M.; Rades, D.; Dunst, J.; Lehnert, H.; Gieseler, F.; Ungefroren, H. Ionizing Radiation Induces a Motile Phenotype in Human Carcinoma Cells in vitro through Hyperactivation of the TGF-beta Signaling Pathway. Cell Mol. Life Sci. 2016, 73, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.A.; Kraut, N.; Beug, H. Molecular Requirements for Epithelial-mesenchymal Transition during Tumor Progression. Curr. Opin. Cell Biol. 2005, 17, 548–558. [Google Scholar] [CrossRef] [PubMed]
- De Melo, J.; Du, G.; Fonseca, M.; Gillespie, L.A.; Turk, W.J.; Rubenstein, J.L.; Eisenstat, D.D. Dlx1 and Dlx2 Function is Necessary for Terminal Differentiation and Survival of Late-born Retinal Ganglion Cells in the Developing Mouse Retina. Development 2005, 132, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, T.M.; McBride, W.H.; Pajonk, F. The Response of CD24(-/low)/CD44+ Breast Cancer-initiating Cells to Radiation. J. Natl. Cancer Inst. 2006, 98, 1777–1785. [Google Scholar] [CrossRef] [Green Version]
- Woodward, W.A.; Chen, M.S.; Behbod, F.; Alfaro, M.P.; Buchholz, T.A.; Rosen, J.M. WNT/beta-catenin Mediates Radiation Resistance of Mouse Mammary Progenitor Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 618–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences |
|---|---|
| OCT4 | |
| Forward | 5′-AGCAAAACCCGGAGGAGT-3′ |
| Reverse | 5′-CCACATCGGCCTGTGTATATC-3′ |
| SOX2 | |
| Forward | 5′-GTGAGCGCCCTGCAGTACAA-3′ |
| Reverse | 5′-GCGAGTAGGACATGTGTAGGTG-3′ |
| LIF | |
| Forward | 5′-GTTCCCCAACAACCTGGACA-3′ |
| Reverse | 5′-ACGACTATGCGGTACAGCTC-3′ |
| SNAIL | |
| Forward | 5′-TTTCCTCGTCAGGAAGCCCTC-3′ |
| Reverse | 5′-TGCTGGAAGGTAAAACTCTGGATTAG-3′ |
| MMP2 | |
| Forward | 5′-GGAAAGCCAGGATCCATTTT-3′ |
| Reverse | 5′-ATGCCGCCTTTAACTGGAG-3′ |
| MMP7 | |
| Forward | 5′-GTCACTTCTTCGGTTGTAGGGA-3′ |
| Reverse | 5′-TCAGAGGAATGTCCCATACCCA-3′ |
| β-actin | |
| Forward | 5′-GACCTGTACGCCAACACAGT-3′ |
| Reverse | 5′-CCAGGGCAGTGATCTCCTTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-R.; Choi, Y.-J.; Kim, J.-Y.; Kim, I.-G.; Jung, U. Repeated Irradiation with γ-Ray Induces Cancer Stemness through TGF-β-DLX2 Signaling in the A549 Human Lung Cancer Cell Line. Int. J. Mol. Sci. 2021, 22, 4284. https://doi.org/10.3390/ijms22084284
Park H-R, Choi Y-J, Kim J-Y, Kim I-G, Jung U. Repeated Irradiation with γ-Ray Induces Cancer Stemness through TGF-β-DLX2 Signaling in the A549 Human Lung Cancer Cell Line. International Journal of Molecular Sciences. 2021; 22(8):4284. https://doi.org/10.3390/ijms22084284
Chicago/Turabian StylePark, Hae-Ran, Yeo-Jin Choi, Jee-Young Kim, In-Gyu Kim, and Uhee Jung. 2021. "Repeated Irradiation with γ-Ray Induces Cancer Stemness through TGF-β-DLX2 Signaling in the A549 Human Lung Cancer Cell Line" International Journal of Molecular Sciences 22, no. 8: 4284. https://doi.org/10.3390/ijms22084284