
The Involvement of Long Non-Coding RNAs in Bone

 ,
,
Abstract
:1. Introduction
- -
- lcRNAs that overlap with one or more exonic or intronic or both regions and are transcripted from the same strand of a coding gene (Sense lncRNAs);
- -
- lncRNAs that overlap in a similar way of sense lncRNAs but are transcripted on the opposite strand (Antisense lncRNAs);
- -
- lncRNAs that originate from the opposite direction although sharing the same promoter of the protein-coding gene (Bidirectional lncRNAs);
- -
- lncRNAs derive totally from introns within coding genes (Intronic lncRNAs).
- -
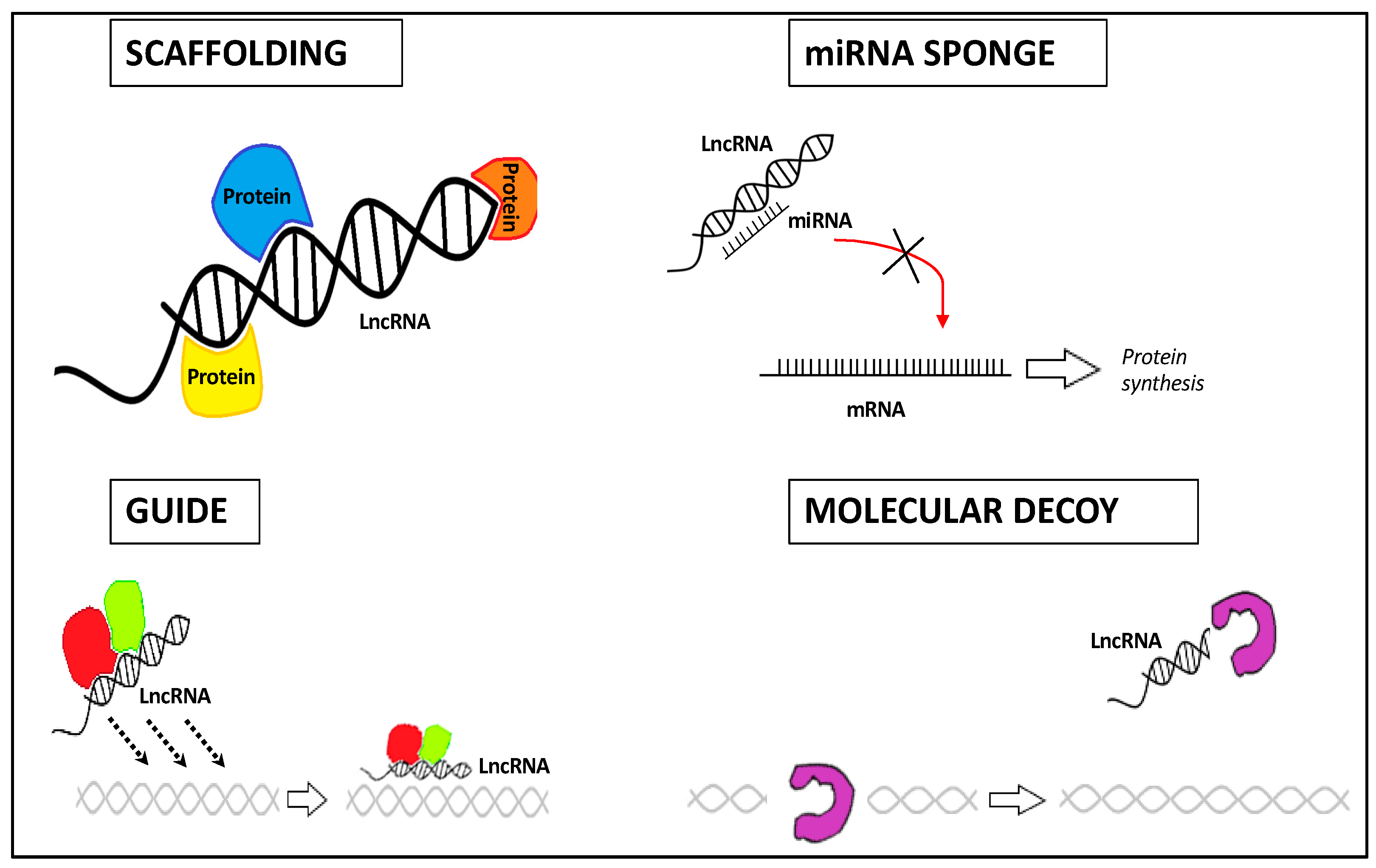
- Their structural plasticity offers interaction sites with a high affinity for proteins, DNAs, or RNAs, thus forming ribonucleoprotein complexes (lncRNA scaffold);
- -
- Depending on the biological context, they can conduct ribonucleoprotein complexes towards a certain genomic location (lncRNA guide);
- -
- lncRNAs can interact with protein complexes, thereby blocking or enhancing protein activities (lncRNA decoy);
- -
- lncRNAs can act as competing endogenous RNAs (ceRNAs) by binding directly and sequestering the miRNA, thus decreasing its regulatory effect on mRNA targets (lncRNA sponge).
2. LncRNAs Regulating Osteoblastogenesis
2.1. LncRNAs as Positive Regulators
2.2. LncRNAs as Negative Regulators
2.3. Bioinformatic Study
3. LncRNAs That Regulate Osteoclastogenesis
3.1. LncRNAs as Positive Regulators
3.2. LncRNAs as Negative Regulators
3.3. Bioinformatics Studies
4. LncRNAs in Primary Bone Tumors and OP
4.1. LncRNA GAS5
4.2. LncRNA TUG1
4.3. LncRNA MALAT1
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hadjidakis, D.J.; Androulakis, I.I. Bone Remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A Transcriptional Activator of Osteoblast Differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone Morphogenetic Proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Globus, R.K.; Plouet, J.; Gospodarowicz, D. Cultured Bovine Bone Cells Synthesize Basic Fibroblast Growth Factor and Store It in Their Extracellular Matrix. Endocrinology 1989, 124, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Clément-Lacroix, P.; Ormandy, C.; Lepescheux, L.; Ammann, P.; Damotte, D.; Goffin, V.; Bouchard, B.; Amling, M.; Gaillard-Kelly, M.; Binart, N.; et al. Osteoblasts Are a New Target for Prolactin: Analysis of Bone Formation in Prolactin Receptor Knockout Mice. Endocrinology 1999, 140, 96–105. [Google Scholar] [CrossRef]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor Necrosis Factor Receptor Family Member RANK Mediates Osteoclast Differentiation and Activation Induced by Osteoprotegerin Ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef] [Green Version]
- Vaananen, H.K.; Zhao, H.; Mulari, M.; Halleen, J.M. The Cell Biology of Osteoclast Function. J. Cell Sci. 2000, 113, 377–381. [Google Scholar]
- Del Fattore, A.; Cappariello, A.; Teti, A. Genetics, Pathogenesis and Complications of Osteopetrosis. Bone 2008, 42, 19–29. [Google Scholar] [CrossRef]
- Palmini, G.; Zonefrati, R.; Mavilia, C.; Aldinucci, A.; Luzi, E.; Marini, F.; Franchi, A.; Capanna, R.; Tanini, A.; Brandi, M.L. Establishment of Cancer Stem Cell Cultures from Human Conventional Osteosarcoma. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [Green Version]
- Grünewald, T.G.P.; Cidre-Aranaz, F.; Surdez, D.; Tomazou, E.M.; de Álava, E.; Kovar, H.; Sorensen, P.H.; Delattre, O.; Dirksen, U. Ewing Sarcoma. Nat. Rev. Dis. Primers 2018, 4, 5. [Google Scholar] [CrossRef]
- Marini, F.; Cianferotti, L.; Brandi, M.L. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.-W.; Huang, K.; Yang, C.; Kang, C.-S. Non-Coding RNAs as Regulators in Epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, N.P.T.; Anderson, B.A.; Guilak, F.; McAlinden, A. Emerging Roles for Long Noncoding RNAs in Skeletal Biology and Disease. Connect. Tissue Res. 2017, 58, 116–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-Coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. In Long Non Coding RNA Biology; Rao, M.R.S., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; pp. 1–46. ISBN 978-981-10-5203-3. [Google Scholar]
- Lander, E.S. Initial Impact of the Sequencing of the Human Genome. Nature 2011, 470, 187–197. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Finishing the Euchromatic Sequence of the Human Genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial Sequencing and Analysis of the Human Genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Dou, P.; Liu, T.; He, S. Application of Long Noncoding RNAs in Osteosarcoma: Biomarkers and Therapeutic Targets. Cell. Physiol. Biochem. 2017, 42, 1407–1419. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin Signature Reveals over a Thousand Highly Conserved Large Non-Coding RNAs in Mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved Function of LincRNAs in Vertebrate Embryonic Development despite Rapid Sequence Evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GENCODE—Home Page. Available online: https://www.gencodegenes.org/ (accessed on 28 December 2020).
- Bouckenheimer, J.; Assou, S.; Riquier, S.; Hou, C.; Philippe, N.; Sansac, C.; Lavabre-Bertrand, T.; Commes, T.; Lemaître, J.-M.; Boureux, A.; et al. Long Non-Coding RNAs in Human Early Embryonic Development and Their Potential in ART. Hum. Reprod. Update 2016, 23, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.C.; Hughes, J.; Graham, B.; Kowalczyk, M.S.; Higgs, D.R.; Ponting, C.P. Chromatin Signatures at Transcriptional Start Sites Separate Two Equally Populated yet Distinct Classes of Intergenic Long Noncoding RNAs. Genome Biol. 2013, 14, R131. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Guo, Q.; Xiao, Y.; Li, C.; Huang, Y.; Luo, X. Regulation of Bone Marrow Mesenchymal Stem Cell Fate by Long Non-Coding RNA. Bone 2020, 141, 115617. [Google Scholar] [CrossRef]
- Yang, Y.; Yujiao, W.; Fang, W.; Linhui, Y.; Ziqi, G.; Zhichen, W.; Zirui, W.; Shengwang, W. The Roles of MiRNA, LncRNA and CircRNA in the Development of Osteoporosis. Biol. Res. 2020, 53, 40. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 Regulates Assembly and Activation of AMPK Complex under Metabolic Stress. Cell Metab. 2019, 30, 157–173.e7. [Google Scholar] [CrossRef]
- Xiao, Y.-Z.; Yang, M.; Xiao, Y.; Guo, Q.; Huang, Y.; Li, C.-J.; Cai, D.; Luo, X.-H. Reducing Hypothalamic Stem Cell Senescence Protects against Aging-Associated Physiological Decline. Cell Metab. 2020, 31, 534–548.e5. [Google Scholar] [CrossRef]
- Singer, R.A.; Arnes, L.; Cui, Y.; Wang, J.; Gao, Y.; Guney, M.A.; Burnum-Johnson, K.E.; Rabadan, R.; Ansong, C.; Orr, G.; et al. The Long Noncoding RNA Paupar Modulates PAX6 Regulatory Activities to Promote Alpha Cell Development and Function. Cell Metab. 2019, 30, 1091–1106.e8. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, P.; Yang, W.; Ruan, X.; Kiesewetter, K.; Zhu, J.; Cao, H. Integrative Transcriptome Analyses of Metabolic Responses in Mice Define Pivotal LncRNA Metabolic Regulators. Cell Metab. 2016, 24, 627–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akerman, I.; Tu, Z.; Beucher, A.; Rolando, D.M.Y.; Sauty-Colace, C.; Benazra, M.; Nakic, N.; Yang, J.; Wang, H.; Pasquali, L.; et al. Human Pancreatic β Cell LncRNAs Control Cell-Specific Regulatory Networks. Cell Metab. 2017, 25, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Qiu, X.; Chen, J.; Sun, X.; Zheng, X.; Zhao, J.; Li, Q.; Wang, Z. A Feed-Forward Regulatory Network LncPCAT1/MiR-106a-5p/E2F5 Regulates the Osteogenic Differentiation of Periodontal Ligament Stem Cells. J. Cell. Physiol. 2019, 234, 19523–19538. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Xu, X.; Feng, X.; Zhu, E.; Zhou, J.; Wang, G.; Tian, L.; Wang, B. A Novel Long Noncoding RNA PGC1β-OT1 Regulates Adipocyte and Osteoblast Differentiation through Antagonizing MiR-148a-3p. Cell Death Differ. 2019, 26, 2029–2045. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, Y.; Hu, Z.; Zhang, L.; Li, G.; Dang, L.; Tan, Y.; Cao, X.; Shi, F.; Zhang, S.; et al. Bone-Targeted LncRNA OGRU Alleviates Unloading-Induced Bone Loss via MiR-320-3p/Hoxa10 Axis. Cell Death Dis. 2020, 11, 382. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, W.; Tian, Q.-J.; Xin, L.; Cui, M.; Li, Y.-K. Effect of LncRNA AK023948 on Rats with Postmenopausal Osteoporosis via PI3K/AKT Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2181–2188. [Google Scholar] [CrossRef]
- Cao, L.; Liu, W.; Zhong, Y.; Zhang, Y.; Gao, D.; He, T.; Liu, Y.; Zou, Z.; Mo, Y.; Peng, S.; et al. Linc02349 Promotes Osteogenesis of Human Umbilical Cord-Derived Stem Cells by Acting as a Competing Endogenous RNA for MiR-25-3p and MiR-33b-5p. Cell Prolif. 2020, 53, e12814. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xiong, Y.; Yan, C.; Zhou, W.; Endo, Y.; Xue, H.; Hu, Y.; Hu, L.; Leng, X.; Liu, J.; et al. LncRNA KCNQ1OT1 Accelerates Fracture Healing via Modulating MiR-701-3p/FGFR3 Axis. FASEB J. 2020, 34, 5208–5222. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-G.; Liao, Z.; Xiao, H.; Liu, H.; Hu, Y.-H.; Liao, Q.-D.; Zhong, D. LncRNA KCNQ1OT1 Promoted BMP2 Expression to Regulate Osteogenic Differentiation by Sponging MiRNA-214. Exp. Mol. Pathol. 2019, 107, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fang, J.; Liu, Y.; Xiao, L. LncRNA LOC100506178 Promotes Osteogenic Differentiation via Regulating MiR-214-5p-BMP2 Axis in Human Bone Marrow Mesenchymal Stem Cells. PeerJ 2020, 8, e8909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, B.; Wang, Z.; Sun, X.; Chen, J.; Zhao, J.; Qiu, X. Long Noncoding RNA LINC00707 Sponges MiR-370-3p to Promote Osteogenesis of Human Bone Marrow-Derived Mesenchymal Stem Cells through Upregulating WNT2B. Stem Cell Res. Ther. 2019, 10, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-C.; Sun, Q.-Z.; Qiao, X.-F.; Li, X.-G.; Yang, J.-H.; Wang, T.-Q.; Xiao, Y.-J.; Qiao, J.-M. LncRNA TUG1 Influences Osteoblast Proliferation and Differentiation through the Wnt/β-Catenin Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4584–4590. [Google Scholar] [CrossRef]
- Hao, R.; Wang, B.; Wang, H.; Huo, Y.; Lu, Y. LncRNA TUG1 Promotes Proliferation and Differentiation of Osteoblasts by Regulating the MiR-545-3p/CNR2 Axis. Braz. J. Med. Biol. Res. 2020, 53, e9798. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, D.; Zhu, Y.; Dong, Y.; Liu, Y. Long Non-Coding RNA GAS5 Promotes Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells by Regulating the MiR-135a-5p/FOXO1 Pathway. Mol. Cell. Endocrinol. 2019, 496, 110534. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Han, Y.; Liu, P.; Huang, Y.; Li, X.; Jia, L.; Zheng, Y.; Li, W. Long Noncoding RNA GAS5 Promotes Osteogenic Differentiation of Human Periodontal Ligament Stem Cells by Regulating GDF5 and P38/JNK Signaling Pathway. Front. Pharmacol. 2020, 11, 701. [Google Scholar] [CrossRef]
- Yi, J.; Liu, D.; Xiao, J. LncRNA MALAT1 Sponges MiR-30 to Promote Osteoblast Differentiation of Adipose-Derived Mesenchymal Stem Cells by Promotion of Runx2 Expression. Cell Tissue Res. 2019, 376, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Xu, W.; Liu, L.; Zhou, X.; Ye, L.; Song, D.; Zhang, L.; Huang, D. Traumatic Compressive Stress Inhibits Osteoblast Differentiation through Long Chain Non-Coding RNA Dancr. J. Periodontol. 2020, 91, 1532–1540. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, L.; Yan, C.; Endo, Y.; Mi, B.; Liu, G. The LncRNA Rhno1/MiR-6979-5p/BMP2 Axis Modulates Osteoblast Differentiation. Int. J. Biol. Sci. 2020, 16, 1604–1615. [Google Scholar] [CrossRef] [Green Version]
- Mulati, M.; Kobayashi, Y.; Takahashi, A.; Numata, H.; Saito, M.; Hiraoka, Y.; Ochi, H.; Sato, S.; Ezura, Y.; Yuasa, M.; et al. The Long Noncoding RNA Crnde Regulates Osteoblast Proliferation through the Wnt/β-Catenin Signaling Pathway in Mice. Bone 2020, 130, 115076. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, M.; Zhong, J.; Yang, L.; Xiao, J.; Jin, F.; Xue, H.; Liu, X.; Liu, H.; Zhang, Y.; et al. The Long Noncoding RNA Lnc-Ob1 Facilitates Bone Formation by Upregulating Osterix in Osteoblasts. Nat. Metab. 2019, 1, 485–496. [Google Scholar] [CrossRef]
- Arumugam, B.; Vishal, M.; Shreya, S.; Malavika, D.; Rajpriya, V.; He, Z.; Partridge, N.C.; Selvamurugan, N. Parathyroid Hormone-Stimulation of Runx2 during Osteoblast Differentiation via the Regulation of Lnc-SUPT3H-1:16 (RUNX2-AS1:32) and MiR-6797-5p. Biochimie 2019, 158, 43–52. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, K.; Zhang, L.; Tan, Y.; Hu, Z.; Dang, L.; Zhou, H.; Li, G.; Wang, H.; Zhang, S.; et al. Targeted Overexpression of the Long Noncoding RNA ODSM Can Regulate Osteoblast Function in Vitro and in Vivo. Cell Death Dis. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, Z.; Shen, L.; Jiang, H. Long Non-Coding RNA SNHG7 Promotes the Fracture Repair through Negative Modulation of MiR-9. Am. J. Transl. Res. 2019, 11, 974–982. [Google Scholar]
- Del Real, A.; López-Delgado, L.; Sañudo, C.; García-Ibarbia, C.; Laguna, E.; Perez-Campo, F.M.; Menéndez, G.; Alfonso, A.; Fakkas, M.; García-Montesinos, B.; et al. Long Noncoding RNAs as Bone Marrow Stem Cell Regulators in Osteoporosis. DNA Cell Biol. 2020, 39, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, M.; Wu, S. The LncRNA MCF2L-AS1 Controls Osteogenic Differentiation by Regulating MiR-33a. Cell Cycle 2020, 19, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yun, X.; Ye, K.; Zhao, H.; An, J.; Zhang, X.; Han, X.; Li, Y.; Wang, S. Long Non-Coding RNA-H19 Stimulates Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells via the MicroRNA-149/SDF-1 Axis. J. Cell. Mol. Med. 2020, 24, 4944–4955. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Jiang, Y.; Liu, Q.; Liu, C.-Z. LncRNA H19 Promotes Matrix Mineralization through Up-Regulating IGF1 by Sponging MiR-185-5p in Osteoblasts. BMC Mol. Cell Biol. 2019, 20, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yang, L.; Ge, D.; Wang, W.; Yin, Z.; Yan, J.; Cao, X.; Jiang, C.; Zheng, S.; Liang, B. Long Non-Coding RNA XIST Promotes Osteoporosis through Inhibiting Bone Marrow Mesenchymal Stem Cell Differentiation. Exp. Ther. Med. 2019, 17, 803–811. [Google Scholar] [CrossRef]
- Niu, S.; Xiang, F.; Jia, H. Downregulation of LncRNA XIST Promotes Proliferation and Differentiation, Limits Apoptosis of Osteoblasts through Regulating MiR-203-3p/ZFPM2 Axis. Connect. Tissue Res. 2020, 1–12. [Google Scholar] [CrossRef]
- Shen, J.-J.; Zhang, C.-H.; Chen, Z.-W.; Wang, Z.-X.; Yang, D.-C.; Zhang, F.-L.; Feng, K.-H. LncRNA HOTAIR Inhibited Osteogenic Differentiation of BMSCs by Regulating Wnt/β-Catenin Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7232–7246. [Google Scholar] [CrossRef]
- He, S.; Yang, S.; Zhang, Y.; Li, X.; Gao, D.; Zhong, Y.; Cao, L.; Ma, H.; Liu, Y.; Li, G.; et al. LncRNA ODIR1 Inhibits Osteogenic Differentiation of HUC-MSCs through the FBXO25/H2BK120ub/H3K4me3/OSX Axis. Cell Death Dis. 2019, 10, 947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, J.; Fu, H.-Q.; Xu, Z.; Fan, W.-J.; Liu, F.; Chen, B. LncRNA SNHG1 Attenuates Osteogenic Differentiation via the MiR-101/DKK1 Axis in Bone Marrow Mesenchymal Stem Cells. Mol. Med. Rep. 2020, 22, 3715–3722. [Google Scholar] [CrossRef]
- Zhang, R.-F.; Liu, J.-W.; Yu, S.-P.; Sun, D.; Wang, X.-H.; Fu, J.-S.; Xie, Z. LncRNA UCA1 Affects Osteoblast Proliferation and Differentiation by Regulating BMP-2 Expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6774–6782. [Google Scholar] [CrossRef]
- Yin, C.; Tian, Y.; Yu, Y.; Li, D.; Miao, Z.; Su, P.; Zhao, Y.; Wang, X.; Pei, J.; Zhang, K.; et al. Long Noncoding RNA AK039312 and AK079370 Inhibits Bone Formation via MiR-199b-5p. Pharmacol. Res. 2020, 105230. [Google Scholar] [CrossRef]
- Jiang, S.-Y.; Miao, Y.-X.; Hirokazu, T.; Zhu, S.-Z.; Lu, J.-S. Effects of LncRNA DANCR on Proliferation and Differentiation of Osteoblasts by Regulating the Wnt/β-Catenin Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5558–5566. [Google Scholar] [CrossRef]
- Cai, N.; Li, C.; Wang, F. Silencing of LncRNA-ANCR Promotes the Osteogenesis of Osteoblast Cells in Postmenopausal Osteoporosis via Targeting EZH2 and RUNX2. Yonsei Med. J. 2019, 60, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tian, Y.; Yin, C.; Huai, Y.; Zhao, Y.; Su, P.; Wang, X.; Pei, J.; Zhang, K.; Yang, C.; et al. Silencing of LncRNA AK045490 Promotes Osteoblast Differentiation and Bone Formation via β-Catenin/TCF1/Runx2 Signaling Axis. Int J. Mol. Sci. 2019, 20, 6229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-G.; Liu, S.-C.; Qiao, X.-F.; Kong, Y.; Liu, J.-G.; Peng, X.-M.; Wang, Y.-X.; Abdulkarim Mohammed Al-Mohana, R.A. LncRNA MEG3 Promotes Proliferation and Differentiation of Osteoblasts through Wnt/β-Catenin Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4521–4529. [Google Scholar] [CrossRef] [PubMed]
- Xiaoling, G.; Shuaibin, L.; Kailu, L. MicroRNA-19b-3p Promotes Cell Proliferation and Osteogenic Differentiation of BMSCs by Interacting with LncRNA H19. BMC Med. Genet. 2020, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Hu, S.; Kang, Z.; Liu, Z.; Yang, W.; Zhang, Y.; Yang, D.; Ruan, W.; Yu, G.; Sun, L.; et al. Identification of Functional LncRNAs Based on Competing Endogenous RNA Network in Osteoblast Differentiation. J. Cell. Physiol. 2020, 235, 2232–2244. [Google Scholar] [CrossRef]
- Cui, Y.; Fu, S.; Sun, D.; Xing, J.; Hou, T.; Wu, X. EPC-Derived Exosomes Promote Osteoclastogenesis through LncRNA-MALAT1. J. Cell. Mol. Med. 2019, 23, 3843–3854. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, Y.; Zhao, Z.; Han, X.; Chen, Y. Knockdown of DANCR Reduces Osteoclastogenesis and Root Resorption Induced by Compression Force via Jagged1. Cell Cycle 2019, 18, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Gong, Z.; Sun, X. LncRNA DANCR Involved Osteolysis after Total Hip Arthroplasty by Regulating FOXO1 Expression to Inhibit Osteoblast Differentiation. J. Biomed. Sci. 2018, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Tao, Z.; Wang, Y. Long Non-Coding RNA DANCR Regulates the Proliferation and Osteogenic Differentiation of Human Bone-Derived Marrow Mesenchymal Stem Cells via the P38 MAPK Pathway. Int. J. Mol. Med. 2018, 41, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Kajiya, H.; Fukushima, H.; Fukushima, A.; Anan, H.; Ozeki, S.; Okabe, K. PTHrP Induces Notch Signaling in Periodontal Ligament Cells. J. Dent. Res. 2009, 88, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Irwandi, R.A.; Vacharaksa, A. The Role of MicroRNA in Periodontal Tissue: A Review of the Literature. Arch. Oral Biol. 2016, 72, 66–74. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.-F.; Li, J.; He, F.; Li, X.; Guo, Y. LncRNA Neat1 Stimulates Osteoclastogenesis Via Sponging MiR-7. J. Bone Miner. Res. 2020, 35, 1772–1781. [Google Scholar] [CrossRef]
- Du, Y.-J.; Yu, Q.-Q.; Zheng, X.-F.; Wang, S.-P. LncRNA TUG1 Positively Regulates Osteoclast Differentiation by Targeting V-Maf Musculoaponeurotic Fibrosarcoma Oncogene Homolog B. Autoimmunity 2020, 1–7. [Google Scholar] [CrossRef]
- Ling, L.; Hu, H.-L.; Liu, K.-Y.; Ram, Y.-I.; Gao, J.-L.; Cao, Y.-M. Long Noncoding RNA MIRG Induces Osteoclastogenesis and Bone Resorption in Osteoporosis through Negative Regulation of MiR-1897. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10195–10203. [Google Scholar] [CrossRef]
- Chen, R.-S.; Zhang, X.-B.; Zhu, X.-T.; Wang, C.-S. LncRNA Bmncr Alleviates the Progression of Osteoporosis by Inhibiting RANML-Induced Osteoclast Differentiation. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9199–9206. [Google Scholar] [CrossRef]
- Zhang, R.; Li, J.; Li, G.; Jin, F.; Wang, Z.; Yue, R.; Wang, Y.; Wang, X.; Sun, Y. LncRNA Nron Regulates Osteoclastogenesis during Orthodontic Bone Resorption. Int. J. Oral Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Wang, X.Q.; Liu, X.T.; Guo, R.; Zhang, R.D. Integrative Analysis Reveals Key MRNAs and LncRNAs in Monocytes of Osteoporotic Patients. Math. Biosci. Eng. 2019, 16, 5947–5971. [Google Scholar] [CrossRef]
- Liu, W.; Li, Z.; Cai, Z.; Xie, Z.; Li, J.; Li, M.; Cen, S.; Tang, S.; Zheng, G.; Ye, G.; et al. LncRNA-mRNA Expression Profiles and Functional Networks in Osteoclast Differentiation. J. Cell. Mol. Med. 2020, 24, 9786–9797. [Google Scholar] [CrossRef]
- Ji, J.; Dai, X.; Yeung, S.-C.J.; He, X. The Role of Long Non-Coding RNA GAS5 in Cancers. Cancer Manag. Res. 2019, 11, 2729–2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Chen, M.; Ma, L.; Dang, X.; Du, G. LncRNA GAS5 Suppresses the Proliferation and Invasion of Osteosarcoma Cells via the MiR-23a-3p/PTEN/PI3K/AKT Pathway. Cell Transplant. 2020, 29. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, D. LncRNA GAS5 Represses Osteosarcoma Cells Growth and Metastasis via Sponging MiR-203a. Cell. Physiol. Biochem. 2018, 45, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Ye, K.; Wang, S.; Zhang, H.; Han, H.; Ma, B.; Nan, W. Long Noncoding RNA GAS5 Suppresses Cell Growth and Epithelial-Mesenchymal Transition in Osteosarcoma by Regulating the MiR-221/ARHI Pathway. J. Cell. Biochem. 2017, 118, 4772–4781. [Google Scholar] [CrossRef]
- Yao, X.; Li, X.; Luo, Y.; Xu, X.; Liu, J.; Bu, J. LncRNA GAS5 Regulates Osteosarcoma Cell Proliferation, Migration, and Invasion by Regulating RHOB via Sponging MiR-663a. Cancer Manag. Res. 2020, 12, 8253–8261. [Google Scholar] [CrossRef]
- Zhao, S.; Xiong, W.; Xu, K. MiR-663a, Regulated by LncRNA GAS5, Contributes to Osteosarcoma Development through Targeting MYL9. Hum. Exp. Toxicol. 2020, 39, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xie, Z.; Li, J.; Lin, J.; Zheng, G.; Liu, W.; Tang, S.; Cen, S.; Ye, G.; Li, Z.; et al. GAS5 Protects against Osteoporosis by Targeting UPF1/SMAD7 Axis in Osteoblast Differentiation. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xie, Z.; Wang, P.; Li, J.; Liu, W.; Tang, S.; Liu, Z.; Wu, X.; Wu, Y.; Shen, H. The Long Noncoding RNA GAS5 Negatively Regulates the Adipogenic Differentiation of MSCs by Modulating the MiR-18a/CTGF Axis as a CeRNA. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Wang, J.-X.; Li, C.-H. LncRNA GAS5 Overexpression Alleviates the Development of Osteoporosis through Promoting Osteogenic Differentiation of MSCs via Targeting MicroRNA-498 to Regulate RUNX2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7757–7765. [Google Scholar] [CrossRef]
- Visconti, V.V.; Fittipaldi, S.; Ciuffi, S.; Marini, F.; Isaia, G.; D’Amelio, P.; Migliaccio, S.; Marcocci, C.; Minisola, S.; Nuti, R.; et al. Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility. Int. J. Mol. Sci. 2020, 21, 6930. [Google Scholar] [CrossRef] [PubMed]
- Cong, C.; Tian, J.; Gao, T.; Zhou, C.; Wang, Y.; Cui, X.; Zhu, L. LncRNA GAS5 Is Upregulated in Osteoporosis and Downregulates MiR-21 to Promote Apoptosis of Osteoclasts. Clin. Interv. Aging 2020, 15, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Young, T.L.; Matsuda, T.; Cepko, C.L. The Noncoding RNA Taurine Upregulated Gene 1 Is Required for Differentiation of the Murine Retina. Curr. Biol. 2005, 15, 501–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wang, J.; Qiu, M.; Xu, L.; Li, M.; Jiang, F.; Yin, R.; Xu, L. Upregulation of the Long Noncoding RNA TUG1 Promotes Proliferation and Migration of Esophageal Squamous Cell Carcinoma. Tumour Biol. 2015, 36, 1643–1651. [Google Scholar] [CrossRef]
- Huang, M.-D.; Chen, W.-M.; Qi, F.-Z.; Sun, M.; Xu, T.-P.; Ma, P.; Shu, Y.-Q. Long Non-Coding RNA TUG1 Is up-Regulated in Hepatocellular Carcinoma and Promotes Cell Growth and Apoptosis by Epigenetically Silencing of KLF2. Mol. Cancer 2015, 14, 165. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, Y.; Gui, Y.; Cai, Z. Long Intergenic Non-Coding RNA TUG1 Is Overexpressed in Urothelial Carcinoma of the Bladder. J. Surg. Oncol. 2013, 107, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Kuang, D.; Zhang, X.; Hua, S.; Dong, W.; Li, Z. Long Non-Coding RNA TUG1 Regulates Ovarian Cancer Proliferation and Metastasis via Affecting Epithelial-Mesenchymal Transition. Exp. Mol. Pathol. 2016, 101, 267–273. [Google Scholar] [CrossRef]
- Isin, M.; Ozgur, E.; Cetin, G.; Erten, N.; Aktan, M.; Gezer, U.; Dalay, N. Investigation of Circulating LncRNAs in B-Cell Neoplasms. Clin. Chim. Acta 2014, 431, 255–259. [Google Scholar] [CrossRef]
- Zhang, E.-b; Yin, D.-d; Sun, M.; Kong, R.; Liu, X.-h; You, L.-h; Han, L.; Xia, R.; Wang, K.-m; Yang, J.-s.; et al. P53-Regulated Long Non-Coding RNA TUG1 Affects Cell Proliferation in Human Non-Small Cell Lung Cancer, Partly through Epigenetically Regulating HOXB7 Expression. Cell Death Dis. 2014, 5, e1243. [Google Scholar] [CrossRef]
- Ma, B.; Li, M.; Zhang, L.; Huang, M.; Lei, J.-B.; Fu, G.-H.; Liu, C.-X.; Lai, Q.-W.; Chen, Q.-Q.; Wang, Y.-L. Upregulation of Long Non-Coding RNA TUG1 Correlates with Poor Prognosis and Disease Status in Osteosarcoma. Tumor Biol. 2016, 37, 4445–4455. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, Y.; Fan, S.; Luo, L. Knockdown of Long Noncoding RNA TUG1 Inhibits the Proliferation and Cellular Invasion of Osteosarcoma Cells by Sponging MiR-153. Oncol. Res. 2018, 26, 665–673. [Google Scholar] [CrossRef]
- Yu, X.; Hu, L.; Li, S.; Shen, J.; Wang, D.; Xu, R.; Yang, H. Long Non-Coding RNA Taurine Upregulated Gene 1 Promotes Osteosarcoma Cell Metastasis by Mediating HIF-1α via MiR-143-5p. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-H.; Cao, Y.-M.; Huang, Y.; Shi, Q.-W.; Guo, J.-H.; Fan, Z.-W.; Li, J.-G.; Chen, B.-W.; Wu, B.-Y. Long Non-Coding RNA TUG1 Contributes to Tumorigenesis of Human Osteosarcoma by Sponging MiR-9-5p and Regulating POU2F1 Expression. Tumour Biol. 2016, 37, 15031–15041. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, T.; Zhang, Z.; Lu, M.; Zhao, W.; Zeng, X.; Zhang, W. Long Non-Coding RNA TUG1 Promotes Migration and Invasion by Acting as a CeRNA of MiR-335-5p in Osteosarcoma Cells. Cancer Sci. 2017, 108, 859–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Tian, G.; Tian, F.; Shao, L. Long Non-Coding RNA TUG1 Promotes Osteosarcoma Cell Proliferation and Invasion through Inhibition of MicroRNA-212-3p Expression. Exp. Ther. Med. 2018, 16, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, K.; Du, X. Long Non-Coding RNA TUG1 Promotes Proliferation and Inhibits Apoptosis of Osteosarcoma Cells by Sponging MiR-132-3p and Upregulating SOX4 Expression. Yonsei Med. J. 2018, 59, 226–235. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Zhao, Y.-C.; Liu, W. Long Non-Coding RNA TUG1 Regulates the Progression and Metastasis of Osteosarcoma Cells via MiR-140-5p/PFN2 Axis. Eur Rev. Med. Pharmacol. Sci. 2019, 23, 9781–9792. [Google Scholar] [CrossRef]
- Cao, J.; Han, X.; Qi, X.; Jin, X.; Li, X. TUG1 Promotes Osteosarcoma Tumorigenesis by Upregulating EZH2 Expression via MiR-144-3p. Int. J. Oncol. 2017, 51, 1115–1123. [Google Scholar] [CrossRef]
- Zhou, Q.; Hu, T.; Xu, Y. Anticancer Potential of TUG1 Knockdown in Cisplatin-Resistant Osteosarcoma through Inhibition of MET/Akt Signalling. J. Drug Target. 2020, 28, 204–211. [Google Scholar] [CrossRef]
- Han, X.; Yang, Y.; Sun, Y.; Qin, L.; Yang, Y. LncRNA TUG1 Affects Cell Viability by Regulating Glycolysis in Osteosarcoma Cells. Gene 2018, 674, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Yun-Bo, F.; Xiao-Po, L.; Xiao-Li, L.; Guo-Long, C.; Pei, Z.; Fa-Ming, T. LncRNA TUG1 Is Upregulated and Promotes Cell Proliferation in Osteosarcoma. Open Med. 2016, 11, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Li, Y. LncRNA TUG1 Promotes the Development of Osteosarcoma through RUNX2. Exp. Ther. Med. 2019, 18, 3002–3008. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-X.; Zhu, Q.-N.; Zhang, H.-B.; Hu, Y.; Wang, G.; Zhu, Y.-S. MALAT1: A Potential Biomarker in Cancer. Cancer Manag. Res. 2018, 10, 6757–6768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sun, G. FOXO1-MALAT1-MiR-26a-5p Feedback Loop Mediates Proliferation and Migration in Osteosarcoma Cells. Oncol. Res. 2017, 25, 1517–1527. [Google Scholar] [CrossRef]
- Duan, G.; Zhang, C.; Xu, C.; Xu, C.; Zhang, L.; Zhang, Y. Knockdown of MALAT1 Inhibits Osteosarcoma Progression via Regulating the MiR-34a/Cyclin D1 Axis. Int. J. Oncol. 2019, 54, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Piao, C.-D.; Ding, J.; Li, Z.-W. LncRNA MALAT1 Facilitates Lung Metastasis of Osteosarcomas through MiR-202 Sponging. Sci. Rep. 2020, 10, 12757. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, W.; Sun, W.; Zheng, B.; Wang, C.; Luo, Z.; Wang, J.; Yan, W. LncRNA MALAT1 Promotes Cancer Metastasis in Osteosarcoma via Activation of the PI3K-Akt Signaling Pathway. Cell. Physiol. Biochem. 2018, 51, 1313–1326. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Yang, T.; Zhao, W.; Wang, N.; Li, P.; Zeng, X.; Zhang, W. Long Non-Coding RNA MALAT1 for Promoting Metastasis and Proliferation by Acting as a CeRNA of MiR-144-3p in Osteosarcoma Cells. Oncotarget 2017, 8, 59417–59434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Pan, X.; Wang, X.; Jiao, X.; Zheng, J.; Li, Z.; Huo, Y. Long Noncoding RNA MALAT1 Promotes Cell Proliferation through Suppressing MiR-205 and Promoting SMAD4 Expression in Osteosarcoma. Oncotarget 2017, 8, 106648–106660. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Qin, B. Long Noncoding RNA MALAT1 Regulates HDAC4-Mediated Proliferation and Apoptosis via Decoying of MiR-140-5p in Osteosarcoma Cells. Cancer Med. 2018, 7, 4584–4597. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Huang, J.; Ni, J.; Song, D.; Ding, M.; Wang, J.; Huang, X.; Li, W. MALAT1 Promotes Osteosarcoma Development by Regulation of HMGB1 via MiR-142-3p and MiR-129-5p. Cell Cycle 2017, 16, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; He, H.; Xiao, W.; Liu, Q.; Deng, Z.; Lu, Y.; Wang, Q.; Zheng, Q.; Li, Y. MALAT1 Promotes Osteosarcoma Development by Targeting TGFA via MIR376A. Oncotarget 2016, 7, 54733–54743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Zhang, Y.; Liu, L.; Yuan, Q. MALAT1 Promotes Proliferation, Migration, and Invasion of MG63 Cells by Upregulation of TGIF2 via Negatively Regulating MiR-129. OncoTargets Ther. 2018, 11, 8729–8740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Dai, Q.; Zeng, F.; Liu, H. MALAT1 Promotes the Proliferation and Metastasis of Osteosarcoma Cells By Activating the Rac1/JNK Pathway Via Targeting MiR-509. Oncol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Liu, Y.; Yang, W.; Xia, Y.; Yang, C.; Yang, S.; Liu, X. Long Noncoding RNA MALAT1 as a Potential Therapeutic Target in Osteosarcoma. J. Orthop. Res. 2016, 34, 932–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Li, Q.; Wang, X.; Jiao, X.; Zheng, J.; Li, Z.; Pan, X. MALAT1 Predicts Poor Survival in Osteosarcoma Patients and Promotes Cell Metastasis through Associating with EZH2. Oncotarget 2017, 8, 46993–47006. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-C.; Tang, C.; Dong, Y.; Zhang, J.; Yuan, T.; Li, X.-L. Targeting LncRNA-MALAT1 Suppresses the Progression of Osteosarcoma by Altering the Expression and Localization of β-Catenin. J. Cancer 2018, 9, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Liang, G.; Yuan, B.; Yang, C.; Gao, R.; Zhou, X. MALAT1 Promotes the Proliferation and Metastasis of Osteosarcoma Cells by Activating the PI3K/Akt Pathway. Tumour Biol. 2015, 36, 1477–1486. [Google Scholar] [CrossRef]
- Zhang, Z.-C.; Tang, C.; Dong, Y.; Zhang, J.; Yuan, T.; Tao, S.-C.; Li, X.-L. Targeting the Long Noncoding RNA MALAT1 Blocks the Pro-Angiogenic Effects of Osteosarcoma and Suppresses Tumour Growth. Int. J. Biol. Sci. 2017, 13, 1398–1408. [Google Scholar] [CrossRef]
- Sun, H.; Lin, D.-C.; Cao, Q.; Pang, B.; Gae, D.D.; Lee, V.K.M.; Lim, H.J.; Doan, N.; Said, J.W.; Gery, S.; et al. Identification of a Novel SYK/c-MYC/MALAT1 Signaling Pathway and Its Potential Therapeutic Value in Ewing Sarcoma. Clin. Cancer Res. 2017, 23, 4376–4387. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Huang, Q.; Hu, J.; Li, L.; Xiao, Y.; Yu, H.; Han, Z.; Wang, T.; Zhou, W.; Wei, H.; et al. EWS-FLI1-Mediated Tenascin-C Expression Promotes Tumour Progression by Targeting MALAT1 through Integrin Α5β1-Mediated YAP Activation in Ewing Sarcoma. Br. J. Cancer 2019, 121, 922–933. [Google Scholar] [CrossRef]
- Hong, J.; Ye, F.; Yu, B.; Gao, J.; Qi, F.; Wang, W. Identification of the Specific MicroRNAs and Competitive Endogenous RNA Mechanisms in Osteoporosis. J. Int. Med. Res. 2020, 48. [Google Scholar] [CrossRef]
- Zheng, S.; Wang, Y.-B.; Yang, Y.-L.; Chen, B.-P.; Wang, C.-X.; Li, R.-H.; Huang, D. LncRNA MALAT1 Inhibits Osteogenic Differentiation of Mesenchymal Stem Cells in Osteoporosis Rats through MAPK Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4609–4617. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, J.; Lei, P.; Wen, T. LncRNA MALAT1 Shuttled by Bone Marrow-Derived Mesenchymal Stem Cells-Secreted Exosomes Alleviates Osteoporosis through Mediating MicroRNA-34c/SATB2 Axis. Aging 2019, 11, 8777–8791. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Yang, Z.; Huang, Z.; Zhou, Y.; Cui, Q.; Dong, D. LncRNADisease 2.0: An Updated Database of Long Non-Coding RNA-Associated Diseases. Nucleic Acids Res. 2019, 47, D1034–D1037. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Kraus, W.L. From Discovery to Function: The Expanding Roles of Long NonCoding RNAs in Physiology and Disease. Endocr. Rev. 2015, 36, 25–64. [Google Scholar] [CrossRef] [Green Version]
- Lassere, M.N. The Biomarker-Surrogacy Evaluation Schema: A Review of the Biomarker-Surrogate Literature and a Proposal for a Criterion-Based, Quantitative, Multidimensional Hierarchical Levels of Evidence Schema for Evaluating the Status of Biomarkers as Surrogate Endpoints. Stat. Methods Med. Res. 2008, 17, 303–340. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of Bone Remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [Green Version]
- Zang, J.; Lu, L.; Yaszemski, M.J. Chapter 3—Bone Disorders. In Materials for Bone Disorders; Bose, S., Bandyopadhyay, A., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 83–118. ISBN 978-0-12-802792-9. [Google Scholar]
- Ko, N.-Y.; Chen, L.-R.; Chen, K.-H. The Role of Micro RNA and Long-Non-Coding RNA in Osteoporosis. Int. J. Mol. Sci. 2020, 21, 4886. [Google Scholar] [CrossRef]
- Hackl, M.; Heilmeier, U.; Weilner, S.; Grillari, J. Circulating MicroRNAs as Novel Biomarkers for Bone Diseases—Complex Signatures for Multifactorial Diseases? Mol. Cell. Endocrinol. 2016, 432, 83–95. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sources | lncRNAs | Regulation Mechanism | Potential Role in Osteoblastogenesis | Study |
|---|---|---|---|---|
| Cells | PCAT1 | Sponging miR-106a-5p (BMP2) | Regulates osteogenic differentiation | [34] |
| Cells | PGC1β-OT1 | Sponging miR-148a-3p (KDM6B) | Regulates osteogenic differentiation | [35] |
| Cells and animals | OGRU | Sponging miR-320-3p (HOXA10) | Regulates bone formation | [36] |
| Animals | AK023948 | Regulates PI3K/AKT pathway | Role in OP occurrence and development | [37] |
| Cells | LINC02349 | Sponging miR-25-3p and miR-33b-5p (SMAD5 and Wnt10b) | Regulates osteogenic differentiation | [38] |
| Cells | KCNQ10T1 | Sponging miR-701-3p (FGFR3) | Regulates osteoblast survival, migration, and proliferation | [39] |
| Cells | KCNQ10T1 | Sponging miR-214 (BMP2) | Regulates osteoblast differentiation | [40] |
| Cells | LOC100506178 | Sponging miR-214-5p (BMP2) | Regulates osteoblast differentiation | [41] |
| Cells | LINC00707 | Sponging miR-370-3p (Wnt2B) | Regulates osteogenic differentiation | [42] |
| Cells | TUG1 | Regulates Wnt/β-catenin pathway | Regulates bone formation | [43] |
| Cells | TUG1 | Sponging miR-545-3p (CNR2) | Regulates osteoblast proliferation, and differentiation | [44] |
| Cells | GAS5 | Sponging miR-135a-3p (FOXO1) | Regulates osteoblast differentiation | [45] |
| Cells | GAS5 | Regulates MAPK signaling pathway | Regulates osteogenic differentiation | [46] |
| Cells | MALAT1 | Sponging miR-30 (RUNX2) | Regulates osteogenic differentiation | [47] |
| Tissue and cells | DANCR | Regulates NF-kB pathway (ALP and RUNX2) | Regulates osteogenic differentiation | [48] |
| Cells and animals | Rhno1 | Sponging miR-6979-5p (BMP2) | Regulates osteoblast differentiation | [49] |
| Cells and animals | Crnde | Regulates Wnt/β-catenin pathway | Regulates osteoblast proliferation and differentiation | [50] |
| Tissue, cells and animals | lnc-ob1 | Regulates OSX expression | Regulates osteoblast differentiation | [51] |
| Cells | lnc-SUPT3H-1:16 | Sponging miR-6797-5p (RUNX2) | Regulates osteoblast differentiation | [52] |
| Cells and animals | ODSM | Sponging miR-139-3p (ELK1) | Regulates bone formation | [53] |
| Tissue and cells | SNHG7 | Sponging miR-9 (TGFBR2, p-smad2 and p-smad3) | Regulates osteoblast activity, migration, and proliferation | [54] |
| Cells | LINC00341 and PACERR | Regulate osteoblast specific marker genes | Role in OP occurrence | [55] |
| Cells | MCF2L-AS1 | Sponging miR-33a (RUNX2) | Regulates osteogenic differentiation | [56] |
| Cells | H19 | Sponging miR-149 (SDF-1) | Regulates osteoblast differentiation | [57] |
| Cells | H19 | Sponging miR-185-5p (IGF-1) | Regulates the mineralization in osteoblasts | [58] |
| Cells | XIST | / | Regulates OP occurrence | [59] |
| Cells and plasma | XIST | Sponging miR-203-3p (ZFPM2) | Regulates osteoblast proliferation and differentiation | [60] |
| Cells and serum | HOTAIR | Regulates Wnt/β-catenin pathway | Regulates osteoblast differentiation | [61] |
| Cells | ODIR1 | Regulates OSX expression | Regulates osteogenic differentiation | [62] |
| Cells | SNHG1 | Sponging miR-101 (DKK1) | Regulates osteogenic differentiation | [63] |
| Cells and plasma | UCA1 | Regulates BMP2/Smad1/5/8 pathway | Regulates osteoblast proliferation and differentiation | [64] |
| Cells and animals | AK079370 and AK039312 | Sponging miR-199b-5p (Wnt/β-catenin pathway) | Regulate bone formation | [65] |
| Cells and Serum | DANCR | Regulates Wnt/β-catenin pathway | Regulates osteoblast proliferation and differentiation | [66] |
| Cells | ANCR | Interacts with EZH2 (RUNX2) | Regulates osteoblast proliferation and differentiation | [67] |
| Cells and animals | AK045490 | Regulates Wnt/β-catenin pathway (RUNX2) | Regulates bone formation | [68] |
| Cells and Serum | MEG3 | Regulates Wnt/β-catenin pathway | Regulates osteoblast proliferation and differentiation | [69] |
| Cells and Serum | H19 | Sponging miR-19b-3p | Role in OP occurrence | [70] |
| Bioinformatic analysis | 662 differentially expressed lncRNAs | Interact with different mRNAs | Regulates osteogenic differentiation | [71] |
| Sources | lncRNAs | Regulation Mechanism | Potential Role in Osteoclastogenesis | Study |
|---|---|---|---|---|
| EPC-derived exosomes | MALAT1 | Sponging miR-124 (ITGB1) | Regulates osteoclast migration and differentiation | [72] |
| Cells | DANCR | Regulates Jagged1 and RANKL expression | Regulates osteoclast formation | [73] |
| Cells | NEAT1 | Sponging miR-7 (PTK2) | Regulates osteoclast formation | [78] |
| Cells | TUG1 | Regulates V-MafB | Regulates osteoclast differentiation | [79] |
| Cells and animals | MIRG | Sponging miR-1897 (NFATC1) | Regulates osteoclast formation | [80] |
| Cells | Bmncr | Downregulates osteoclast-related marker genes | Regulates osteoclast differentiation | [81] |
| Cells and animals | NRON | Regulates NFATC1 expression | Regulates osteoclastogenesis during orthodontic bone resorption | [82] |
| Bioinformatic analysis | RP11-498C9.17 | Regulates genes mainly enriched in pathways regulating bone mineral density | Regulates osteoclastogenesis | [83] |
| Bioinformatic analysis | ENSG00000257764.2 | Sponging miR-106a-5p (TIMP2) | Regulates osteoclast differentiation | [84] |
| Disease | Sources | Pattern Expression Profile | Regulation Mechanism | Related Effects | Study |
|---|---|---|---|---|---|
| OS | Tissues and cells | ↓ | Sponging miR-23a-3p (PTEN/PI3K/AKT) | Regulates OS cell proliferation and invasion | [86] |
| OS | Cells | ↓ | Sponging miR-203a (TIMP2) | Regulates OS cell growth and metastasis | [87] |
| OS | Tissues and cells | ↓ | Sponging miR-221 (ARHI) | Regulates OS cell growth and EMT | [88] |
| OS | Tissues and cells | ↓ | Sponging miR-663a (RHOB) | Regulates OS progression | [89] |
| OS | Tissues and cells | ↓ | Sponging miR-663a (MYL9) | Regulates OS progression | [90] |
| OP | Tissues, cells, and animals | ↓ | Regulation of UPF1/SMAD7 axis | Regulates osteoblast differentiation | [91] |
| OP | Cells | ↓ | Sponging miR-498 (RUNX2) | Regulates osteogenic differentiation | [93] |
| OP | Serum | ↑ | / | / | [94] |
| OP | Plasma | ↑ | Downregulation of miR-21 | Regulates apoptosis of osteoclasts | [95] |
| Disease | Sources | Pattern Expression Profile | Regulation Mechanism | Related Effects | Study |
|---|---|---|---|---|---|
| OS | Tissues, cells, and animals | ↑ | Sponging miR-153 | Contributes to OS development | [104] |
| OS | Tissues, cells, and animals | ↑ | Sponging miR-143-5p (HIF-1α) | Regulates OS cell proliferation, metastasis, and angiogenesis | [105] |
| OS | Cells and animals | ↑ | Sponging miR-9-5p (POU2F1) | Facilitates OS tumorigenesis | [106] |
| OS | Tissues and cells | ↑ | Sponging miR-335-5p (ROCK1) | Regulates OS cell migration and invasion | [107] |
| OS | Tissues and cells | ↑ | Sponging miR-212-3p | Regulates OS cell proliferation and invasion | [108] |
| OS | Tissues and cells | ↑ | Sponging miR-132-3p (SOX4) | Regulates OS cell proliferation and apoptosis | [109] |
| OS | Tissues and cells | ↑ | Sponging miR-140-5p (PFN2) | Regulates OS cell proliferation, migration, and invasion | [110] |
| OS | Tissues and cells | ↑ | Sponging miR-144-3p (EZH2) | Regulates OS cell migration and EMT | [111] |
| OS | Cells and animals | ↑ | Regulation of MET/Akt signaling | Regulates cell growth and apoptosis under DDP treatment | [112] |
| OS | Cells | ↑ | Regulation of HK2 | Regulates OS cell viability | [113] |
| OS | Cells | ↑ | Regulation of AKT signaling pathway | Regulates OS cell proliferation | [114] |
| OS | Tissues, cells, and plasma | ↑ | Regulation of RUNX2 expression | Regulates OS development | [115] |
| Disease | Sources | Pattern Expression Profile | Regulation Mechanism | Related Effects | Study |
|---|---|---|---|---|---|
| OS | Tissues and, cells | ↑ | Sponging miR-26a-5p (FOXO1) | Mediates OS cells proliferation and migration | [117] |
| OS | Tissues and cells | ↑ | Sponging miR-34a (CCND1) | Promotes OS progression | [118] |
| OS | Tissues, cells, and animals | ↑ | Sponging miR-202 | Facilitates lung metastasis of OS | [119] |
| OS | Tissues, cells, and animals | ↑ | Sponging miR-129-5p (RET-Akt pathway) | Promotes OS metastasis | [120] |
| OS | Tissues, cells, and animals | ↑ | Sponging mir-144-3p (ROCK1/ROCK2) | Promotes OS cells metastasis and proliferation | [121] |
| OS | Tissues and cells | ↑ | Sponging miR-205 (SMAD4) | Promotes OS cells proliferation | [122] |
| OS | Tissues and cells | ↑ | Sponging miR-140-5p (HDAC4) | Regulates OS cells proliferation and apoptosis | [123] |
| OS | Tissues and cells | ↑ | Sponging miR-142-3p and miR-129-5p (HMGB1) | Promotes OS development | [124] |
| OS | Tissues and cells | ↑ | Sponging miR-376a (TGFA) | Promotes OS development | [125] |
| OS | Tissues and cells | ↑ | Sponging miR-129-5p (TGIF2) | Promotes proliferation, migration, and invasion of OS cells | [126] |
| OS | Tissues, cells, and animals | ↑ | Sponging miR-509 (Rac1/JNK Pathway) | Promotes the proliferation and metastasis of OS Cells | [127] |
| OS | Tissues and cells | ↑ | Regulating RhoA/ROCK pathway | Promotes OS development | [128] |
| OS | Tissues, cells, and animals | ↑ | Regulating E-cadherin and β-catenin expression | Supports OS metastasis | [129,130] |
| OS | Tissues and cell | ↑ | Regulating of PI3K/AKT signaling pathway | Regulates proliferation and metastasis of OS | [131] |
| OS | Cells and animals | ↑ | Regulating mTOR and HIF-1α | Induces pro-angiogenic effects in OS | [132] |
| ES | Cells and animals | ↑ | Regulated by SYK/cMYC | Contributes to the ES malignancy | [133] |
| ES | Cells and animals | ↑ | Regulated by TNC/Integrin α5β1/YAP axis | Regulates ES progression | [134] |
| OP | Bioinformatic studies | / | Regulating miR-22-3p-PTEN/ESR1/ERBB3/CSF1R/CDK6 and miR-32-3p-SP1/FZD6 axis | Possible role in OP biology | [135] |
| OP | Cells and animals | ↓ | Promoting the activation of the MAPK signaling pathway | Contributes to the OS onset and progression | [136] |
| OP | Cells and animals | ↑ | Sponging miR-34c (SATB2) | Mitigates the symptoms of OP | [137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aurilia, C.; Donati, S.; Palmini, G.; Miglietta, F.; Iantomasi, T.; Brandi, M.L. The Involvement of Long Non-Coding RNAs in Bone. Int. J. Mol. Sci. 2021, 22, 3909. https://doi.org/10.3390/ijms22083909
Aurilia C, Donati S, Palmini G, Miglietta F, Iantomasi T, Brandi ML. The Involvement of Long Non-Coding RNAs in Bone. International Journal of Molecular Sciences. 2021; 22(8):3909. https://doi.org/10.3390/ijms22083909
Chicago/Turabian StyleAurilia, Cinzia, Simone Donati, Gaia Palmini, Francesca Miglietta, Teresa Iantomasi, and Maria Luisa Brandi. 2021. "The Involvement of Long Non-Coding RNAs in Bone" International Journal of Molecular Sciences 22, no. 8: 3909. https://doi.org/10.3390/ijms22083909