
Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge

 ,
,
Abstract
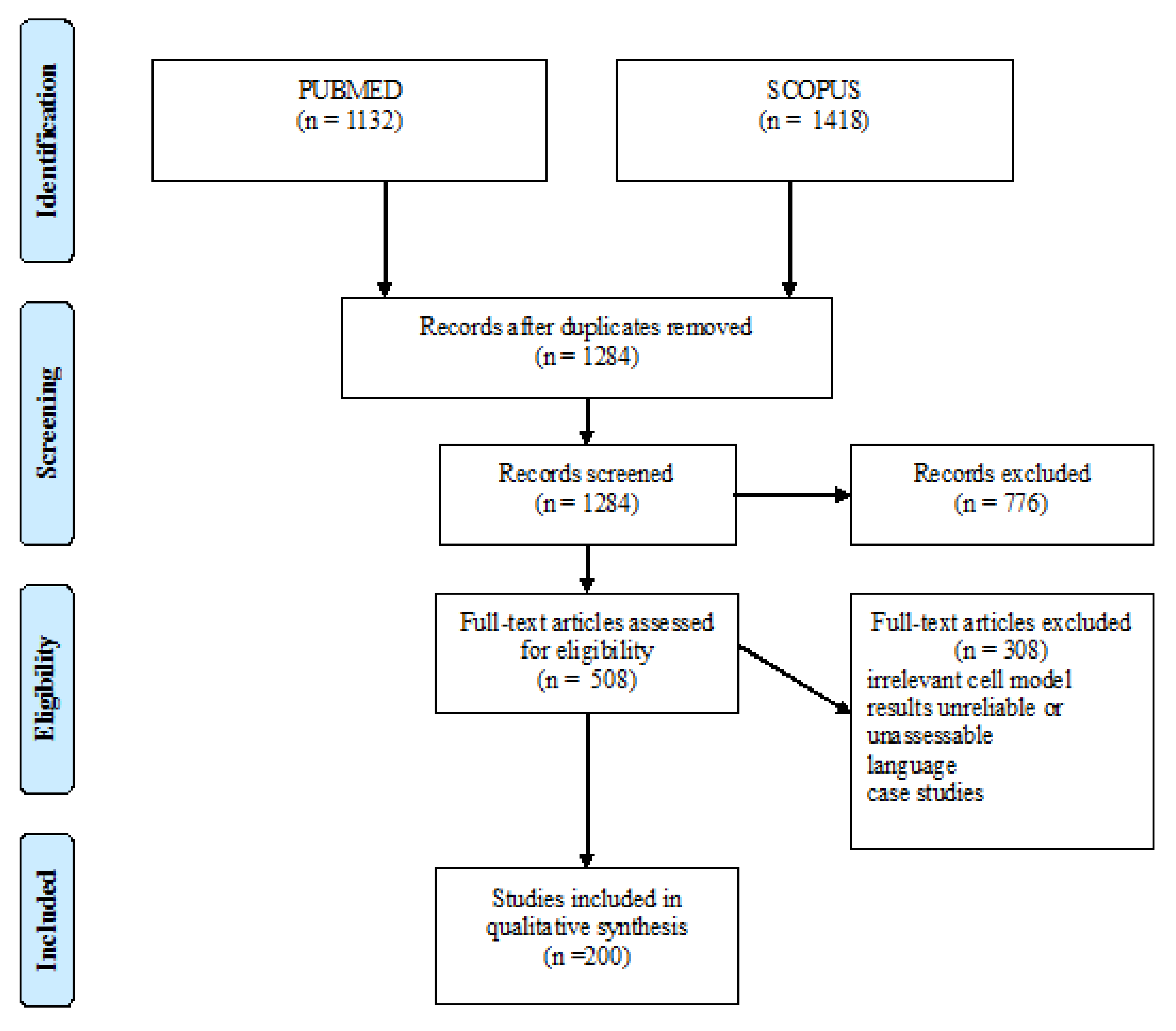
:1. Methods
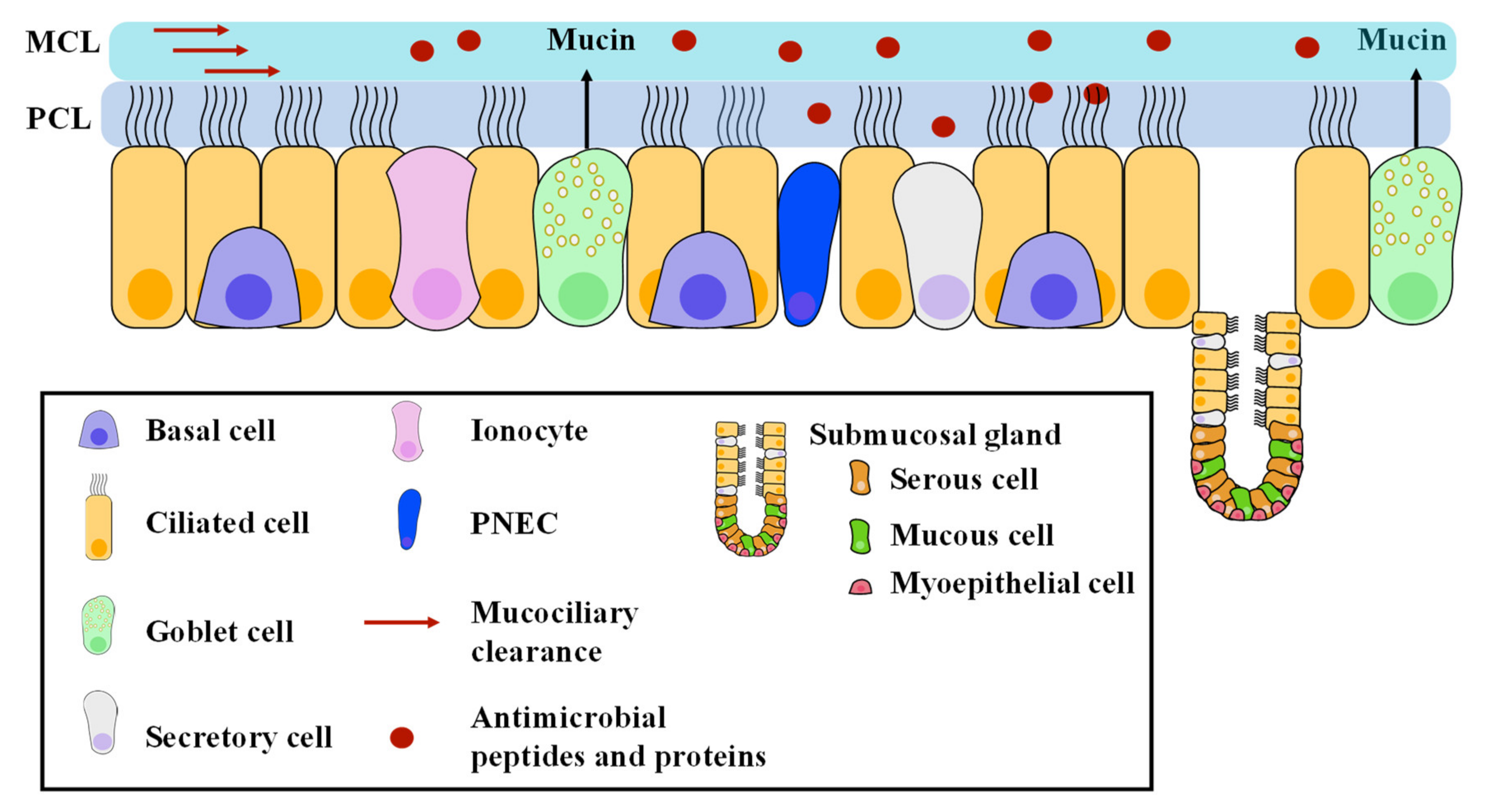
2. Surface Liquid of the Airway Bronchial Epithelium, an Important Player in Airway Physiology
3. ASL pH in Physiology and Disease
3.1. Methods of Measurements
3.2. ASL pH Values in Physiological Conditions
3.3. ASL pH Values in Disease
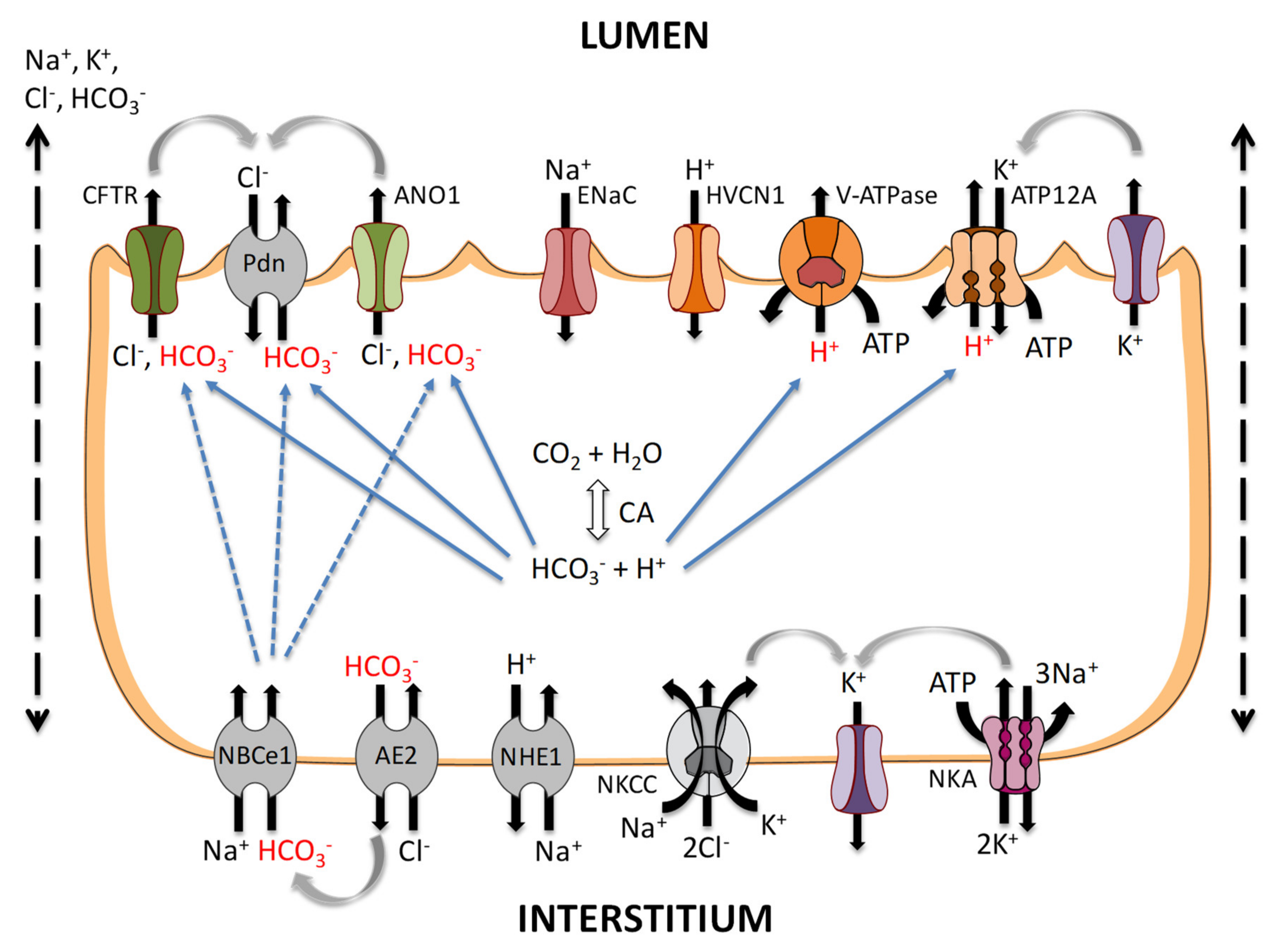
4. Bicarbonate Transport in Airway Cells
4.1. Apical HCO3− Transport
4.1.1. CFTR
4.1.2. ANO1
4.1.3. Apical Cl−/HCO3− Exchangers
4.1.4. SLC26A4 (Pendrin/PDS)
4.1.5. SLC26A9
4.2. Basolateral HCO3− Transport
4.2.1. Cl−/HCO3− Exchanger (AE2)
4.2.2. Electrogenic Na-Coupled Bicarbonate Co-Transport (NBCe, SLC4A4)
4.2.3. Carbonic Anhydrases (CA)
4.3. Paracellular HCO3− Transport
5. Acid Transport in Airway Cells
5.1. Basolateral Secretion: Na+/H+ Exchangers
5.2. Apical Secretion
5.2.1. H+/K+ ATPase
5.2.2. Hydrogen Voltage-Gated Channel 1 (HVCN1)
5.2.3. Vacuolar H+-ATPase
6. Proposed Model
7. Current Gaps in Knowledge
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tam, A.; Wadsworth, S.; Dorscheid, D.; Man, S.F.P.; Sin, D.D. The airway epithelium: More than just a structural barrier. Ther. Adv. Respir. Dis. 2011, 5, 255–273. [Google Scholar] [CrossRef]
- Widdicombe, J.H. Regulation of the depth and composition of airway surface liquid. J. Anat. 2002, 201, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V.; Dickey, B.F. Airway Mucus Function and Dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Widdicombe, J.H.; Wine, J.J. Airway gland structure and function. Physiol. Rev. 2015, 95, 1241–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Scudieri, P.; Musante, I.; Venturini, A.; Guidone, D.; Genovese, M.; Cresta, F.; Caci, E.; Palleschi, A.; Poeta, M.; Santamaria, F.; et al. Ionocytes and CFTR Chloride Channel Expression in Normal and Cystic Fibrosis Nasal and Bronchial Epithelial Cells. Cells 2020, 9, 2090. [Google Scholar] [CrossRef]
- Montoro, D.T.; Haber, A.L.; Biton, M.; Vinarsky, V.; Birket, S.; Yuan, F.; Chen, S.; Leung, H.M.; Villoria, J.; Rogel, N.; et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 2018, 560, 319–324. [Google Scholar] [CrossRef]
- Song, Y.; Thiagarajah, J.; Verkman, A.S. Sodium and Chloride Concentrations, pH, and Depth of Airway Surface Liquid in Distal Airways. J. Gen. Physiol. 2003, 122, 511–519. [Google Scholar] [CrossRef]
- Williams, O.W.; Sharafkhaneh, A.; Kim, V.; Dickey, B.F.; Evans, C.M. Airway mucus: From production to secretion. Am. J. Respir. Cell Mol. Biol. 2006, 34, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Henke, M.O.; John, G.; Germann, M.; Lindemann, H.; Rubin, B.K. MUC5AC and MUC5B mucins increase in cystic fibrosis airway secretions during pulmonary exacerbation. Am. J. Respir. Crit. Care Med. 2007, 175, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef] [Green Version]
- Tarran, R.; Grubb, B.R.; Gatzy, J.T.; Davis, C.W.; Boucher, R.C. The relative roles of passive surface forces and active ion transport in the modulation of airway surface liquid volume and composition. J. Gen. Physiol. 2001, 118, 223–236. [Google Scholar] [CrossRef]
- Korbmacher, J.P.; Michel, C.; Neubauer, D.; Thompson, K.; Mizaikoff, B.; Frick, M.; Dietl, P.; Wittekindt, O.H. Amiloride-sensitive fluid resorption in NCI-H441 lung epithelia depends on an apical Cl- conductance. Physiol. Rep. 2014, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ruffin, M.; Voland, M.; Marie, S.; Bonora, M.; Blanchard, E.; Blouquit-Laye, S.; Naline, E.; Puyo, P.; Le Rouzic, P.; Guillot, L.; et al. Anoctamin 1 dysregulation alters bronchial epithelial repair in cystic fibrosis. Biochim. Biophys. Acta Mol. Basis Dis. 2013, 1832, 2340–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudouin-Legros, M.; Hamdaoui, N.; Borot, F.; Fritsch, J.; Ollero, M.; Planelles, G.; Edelman, A. Control of basal CFTR gene expression by bicarbonate-sensitive adenylyl cyclase in human pulmonary cells. Cell. Physiol. Biochem. 2008, 21, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Garland, A.L.; Walton, W.G.; Coakley, R.D.; Tan, C.D.; Gilmore, R.C.; Hobbs, C.A.; Tripathy, A.; Clunes, L.A.; Bencharit, S.; Stutts, M.J.; et al. Molecular basis for pH-dependent mucosal dehydration in cystic fibrosis airways. Proc. Natl. Acad. Sci. USA 2013, 110, 15973–15978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.S.; Ahmad, S.; Wu, T.; Walton, W.G.; Redinbo, M.R.; Tarran, R. SPLUNC1 is an allosteric modulator of the epithelial sodium channel. FASEB J. 2018, 32, 2478–2491. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Caballero, A.; Rasmussen, J.E.; Gaillard, E.; Watson, M.J.; Olsen, J.C.; Donaldson, S.H.; Stutts, M.J.; Tarran, R. SPLUNC1 regulates airway surface liquid volume by protecting ENaC from proteolytic cleavage. Proc. Natl. Acad. Sci. USA 2009, 106, 11412–11417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bals, R.; Hiemstra, P.S. Innate immunity in the lung: How epithelial cells fight against respiratory pathogens. Eur. Respir. J. 2004, 23, 327–333. [Google Scholar] [CrossRef]
- Ridley, C.; Thornton, D.J. Mucins: The frontline defence of the lung. Biochem. Soc. Trans. 2018, 46, 1099–1106. [Google Scholar] [CrossRef] [Green Version]
- Alaiwa, M.H.A.; Reznikov, L.R.; Gansemer, N.D.; Sheets, K.A.; Horswill, A.R.; Stoltz, D.A.; Zabner, J.; Welsh, M.J. pH modulates the activity and synergism of the airway surface liquid antimicrobials β-defensin-3 and LL-37. Proc. Natl. Acad. Sci. USA 2014, 111, 18703–18708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzulo, A.A.; Tang, X.X.; Hoegger, M.J.; Abou Alaiwa, M.H.; Ramachandran, S.; Moninger, T.O.; Karp, P.H.; Wohlford-Lenane, C.L.; Haagsman, H.P.; Eijk, M.V.; et al. Reduced airway surface pH impairs bacterial killing in the porcine cystic fibrosis lung. Nature 2012, 487, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Dobay, O.; Laub, K.; Stercz, B.; Kéri, A.; Balázs, B.; Tóthpál, A.; Kardos, S.; Jaikumpun, P.; Ruksakiet, K.; Quinton, P.M.; et al. Bicarbonate inhibits bacterial growth and biofilm formation of prevalent cystic fibrosis pathogens. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gawande, P.V.; Lovetri, K.; Yakandawala, N.; Romeo, T.; Zhanel, G.G.; Cvitkovitch, D.G.; Madhyastha, S. Antibiofilm activity of sodium bicarbonate, sodium metaperiodate and SDS combination against dental unit waterline-associated bacteria and yeast. J. Appl. Microbiol. 2008, 105, 986–992. [Google Scholar] [CrossRef]
- Kaushik, K.S.; Stolhandske, J.; Shindell, O.; Smyth, H.D.; Gordon, V.D. Tobramycin and bicarbonate synergise to kill planktonic Pseudomonas aeruginosa, but antagonise to promote biofilm Survival. NPJ Biofilms Microbiomes 2016, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratten, J.; Wiecek, J.; Mordan, N.; Lomax, A.; Patel, N.; Spratt, D.; Middleton, A.M. Physical disruption of oral biofilms by sodium bicarbonate: An in vitro study. Int. J. Dent. Hyg. 2016, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Tang, X.; Xu, W.; Diao, R.; Cai, Z.; Chan, H.C. A host defense mechanism involving CFTR-mediated bicarbonate secretion in bacterial prostatitis. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.S.; Ernst, S.; Tang, X.X.; Karp, P.H.; Parker, C.P.; Ostedgaard, L.S.; Welsh, M.J. Relationships among CFTR expression, HCO3− secretion, and host defense may inform gene- and cell-based cystic fibrosis therapies. Proc. Natl. Acad. Sci. USA 2016, 113, 5382–5387. [Google Scholar] [CrossRef] [Green Version]
- Knowles, M.R.; Boucher, R.C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Investig. 2002, 109, 571–577. [Google Scholar] [CrossRef]
- Verdugo, P. Goblet Cells Secretion And Mucogenesis. Annu. Rev. Physiol. 1990, 52, 157–176. [Google Scholar] [CrossRef]
- Verdugo, P. Mucin Exocytosis. Am. Rev. Respir. Dis. 1991, 144, S33–S37. [Google Scholar] [CrossRef] [PubMed]
- Quinton, P.M. Cystic fibrosis: Impaired bicarbonate secretion and mucoviscidosis. Lancet 2008, 372, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.T.; Yang, N.; Quinton, P.M.; Chin, W.C. A new role for bicarbonate in mucus formation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, L.H.; Evans, J.R.; Wang, T.T.; Ford, A.A.; Makhov, A.M.; Nguyen, K.; Coakley, R.D.; Griffith, J.D.; Davis, C.W.; Ballard, S.T.; et al. Defective postsecretory maturation of MUC5B mucin in cystic fibrosis airways. JCI Insight 2017, 2, e89752. [Google Scholar] [CrossRef] [Green Version]
- Clary-Meinesz, C.; Mouroux, J.; Cosson, J.; Huitorel, P.; Blaive, B. Influence of external pH on ciliary beat frequency in human bronchi and bronchioles. Eur. Respir. J. 1998, 11, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Schmid, A.; Sutto, Z.; Nlend, M.C.; Horvath, G.; Schmid, N.; Buck, J.; Levin, L.R.; Conner, G.E.; Fregien, N.; Salathe, M. Soluble adenylyl cyclase is localized to cilia and contributes to ciliary beat frequency regulation via production of cAMP. J. Gen. Physiol. 2007, 130, 99–109. [Google Scholar] [CrossRef]
- Schmid, A.; Sutto, Z.; Schmid, N.; Novak, L.; Ivonnet, P.; Horvath, G.; Conner, G.; Fregien, N.; Salathe, M. Decreased soluble adenylyl cyclase activity in cystic fibrosis is related to defective apical bicarbonate exchange and affects ciliary beat frequency regulation. J. Biol. Chem. 2010, 285, 29998–30007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delpiano, L.; Thomas, J.J.; Yates, A.R.; Rice, S.J.; Gray, M.A.; Saint-Criq, V. Esomeprazole increases airway surface liquid ph in primary cystic fibrosis epithelial cells. Front. Pharmacol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.X.; Ostedgaard, L.S.; Hoegger, M.J.; Moninger, T.O.; Karp, P.H.; McMenimen, J.D.; Choudhury, B.; Varki, A.; Stoltz, D.A.; Welsh, M.J. Acidic pH increases airway surface liquid viscosity in cystic fibrosis. J. Clin. Investig. 2016, 126, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.S.; Meyerholz, D.K.; Tang, X.X.; Reznikov, L.; Abou Alaiwa, M.; Ernst, S.E.; Karp, P.H.; Wohlford-Lenane, C.L.; Heilmann, K.P.; Leidinger, M.R.; et al. Airway acidification initiates host defense abnormalities in cystic fibrosis mice. Science 2016, 351, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Lennox, A.T.; Coburn, S.L.; Leech, J.A.; Heidrich, E.M.; Kleyman, T.R.; Wenzel, S.E.; Pilewski, J.M.; Corcoran, T.E.; Myerburg, M.M. ATP12A promotes mucus dysfunction during Type 2 airway inflammation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Coakley, R.D.; Grubb, B.R.; Paradiso, A.M.; Gatzy, J.T.; Johnson, L.G.; Kreda, S.M.; O’Neal, W.K.; Boucher, R.C. Abnormal surface liquid pH regulation by cultured cystic fibrosis bronchial epithelium. Proc. Natl. Acad. Sci. USA 2003, 100, 16083–16088. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, R.; Benton, D.C.H.; Munye, M.M.; Rangseesorranan, S.; Hart, S.L.; Moss, G.W.J. A Nanosensor Toolbox for Rapid, Label-Free Measurement of Airway Surface Liquid and Epithelial Cell Function. ACS Appl. Mater. Interfaces 2019, 11, 8731–8739. [Google Scholar] [CrossRef]
- Abou Alaiwa, M.H.; Beer, A.M.; Pezzulo, A.A.; Launspach, J.L.; Horan, R.A.; Stoltz, D.A.; Starner, T.D.; Welsh, M.J.; Zabner, J. Neonates with cystic fibrosis have a reduced nasal liquid pH; A small pilot study. J. Cyst. Fibros. 2014, 13, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Abou Alaiwa, M.H.; Launspach, J.L.; Grogan, B.; Carter, S.; Zabner, J.; Stoltz, D.A.; Singh, P.K.; McKone, E.F.; Welsh, M.J. Ivacaftor-induced sweat chloride reductions correlate with increases in airway surface liquid pH in cystic fibrosis. JCI Insight 2018, 3, 10–13. [Google Scholar] [CrossRef]
- Alaiwa, M.H.A.; Launspach, J.L.; Sheets, K.A.; Rivera, J.A.; Gansemer, N.D.; Taft, P.J.; Thorne, P.S.; Welsh, M.J.; Stoltz, D.A.; Zabner, J. Repurposing tromethamine as inhaled therapy to treat CF airway disease. JCI Insight 2016, 1, 1–11. [Google Scholar] [CrossRef]
- McShane, D.; Davies, J.C.; Davies, M.G.; Bush, A.; Geddes, D.M.; Alton, E.W.F.W. Airway surface pH in subjects with cystic fibrosis. Eur. Respir. J. 2003, 21, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Schultz, A.; Puvvadi, R.; Borisov, S.M.; Shaw, N.C.; Klimant, I.; Berry, L.J.; Montgomery, S.T.; Nguyen, T.; Kreda, S.M.; Kicic, A.; et al. Airway surface liquid pH is not acidic in children with cystic fibrosis. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrin, F.; Voisin, C.; Macquet, V.; Robin, H.; Lequien, P. Apport de la pH metrie bronchique in situ. Progr. Resp. Res. 1971, 6, 372–383. [Google Scholar]
- Benedetto, R.; Centeio, R.; Ousingsawat, J.; Schreiber, R.; Janda, M.; Kunzelmann, K. Transport properties in CFTR−/− knockout piglets suggest normal airway surface liquid pH and enhanced amiloride-sensitive Na+ absorption. Pflugers Arch. Eur. J. Physiol. 2020, 472, 1507–1519. [Google Scholar] [CrossRef]
- Jayaraman, S.; Song, Y.; Vetrivel, L.; Shankar, L.; Verkman, A.S. Noninvasive in vivo fluorescence measurement of airway-surface liquid depth, salt concentration, and pH. J. Clin. Investig. 2001, 107, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Jayaraman, S.; Song, Y.; Verkman, A.S. Airway surface liquid pH in well-differentiated airway epithelial cell cultures and mouse trachea. Am. J. Physiol. Cell Physiol. 2001, 281. [Google Scholar] [CrossRef]
- Birket, S.; Tuggle, K.; Oden, A.; Fernandez, C.; Chu, K.; Tearney, G.; Fanucchi, M.; Sorscher, E.; Rowe, S. Poster Session Abstracts. Pediatr. Pulmonol. 2016, 51, S194–S485. [Google Scholar] [CrossRef] [Green Version]
- Kyle, H.; Ward, J.P.T.; Widdicombe, J.G. Control of pH of airway surface liquid of the ferret trachea in vitro. J. Appl. Physiol. 1990, 68, 135–140. [Google Scholar] [CrossRef]
- Nielson, D.W.; Goerke, J.; Clements, J.A. Alveolar subphase pH in the lungs of anesthetized rabbits. Proc. Natl. Acad. Sci. USA 1981, 78, 7119–7123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inglis, S.K.; Wilson, S.M.; Olver, R.E. Secretion of acid and base equivalents by intact distal airways. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Thiagarajah, J.R.; Song, Y.; Haggie, P.M.; Verkman, A.S. A small molecule CFTR inhibitor produces cystic fibrosis-like submucosal gland fluid secretions in normal airways. FASEB J. 2004, 18, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Simonin, J.; Bille, E.; Crambert, G.; Noel, S.; Dreano, E.; Edwards, A.; Hatton, A.; Pranke, I.; Villeret, B.; Cottart, C.H.; et al. Airway surface liquid acidification initiates host defense abnormalities in Cystic Fibrosis. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, I.; Nilsson, H.; Henriksnäs, J.; Roomans, G.M. X-ray microanalysis of apical fluid in cystic fibrosis airway epithelial cell lines. Cell. Physiol. Biochem. 2006, 17, 13–20. [Google Scholar] [CrossRef]
- Zając, M.; Lewenstam, A.; Stobiecka, M.; Dołowy, K. New ISE-Based Apparatus for Na+, K+, Cl−, pH and Transepithelial Potential Difference Real-Time Simultaneous Measurements of Ion Transport across Epithelial Cells Monolayer–Advantages and Pitfalls. Sensors 2019, 19, 1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.B.; Moody, M.; Koh, D.S. Epithelial monolayer culture system for real-time single-cell analyses. Physiol. Rep. 2014, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Liao, J.; Huang, J.; Robert, R.; Palmer, M.L.; Fahrenkrug, S.C.; O’Grady, S.M.; Hanrahan, J.W. Bicarbonate-dependent chloride transport drives fluid secretion by the human airway epithelial cell line Calu-3. J. Physiol. 2012, 590, 5273–5297. [Google Scholar] [CrossRef]
- Muraglia, K.A.; Chorghade, R.S.; Kim, B.R.; Tang, X.X.; Shah, V.S.; Grillo, A.S.; Daniels, P.N.; Cioffi, A.G.; Karp, P.H.; Zhu, L.; et al. Small-molecule ion channels increase host defences in cystic fibrosis airway epithelia. Nature 2019, 567, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, Á.G.; Clauzure, M.; Massip-Copiz, M.M.; Cancio, C.E.; Asensio, C.J.A.; Mori, C.; Santa-Coloma, T.A. Impairment of CFTR activity in cultured epithelial cells upregulates the expression and activity of LDH resulting in lactic acid hypersecretion. Cell. Mol. Life Sci. 2019, 76, 1579–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, S.; Joo, N.S.; Reitz, B.; Wine, J.J.; Verkman, A.S. Submucosal gland secretions in airways from cystic fibrosis patients have normal [Na+] and pH but elevated viscosity. Proc. Natl. Acad. Sci. USA 2001, 98, 8119–8123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreindler, J.L.; Bertrand, C.A.; Lee, R.J.; Karasic, T.; Aujla, S.; Pilewski, J.M.; Frizzell, R.A.; Kolls, J.K. Interleukin-17A induces bicarbonate secretion in nrmal human bronchial epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Haggie, P.M.; Phuan, P.W.; Tan, J.A.; Zlock, L.; Finkbeiner, W.E.; Verkman, A.S. Inhibitors of pendrin anion exchange identified in a small molecule screen increase airway surface liquid volume in cystic fibrosis. FASEB J. 2016, 30, 2187–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudieri, P.; Musante, I.; Caci, E.; Venturini, A.; Morelli, P.; Walter, C.; Tosi, D.; Palleschi, A.; Martin-Vasallo, P.; Sermet-Gaudelus, I.; et al. Increased expression of ATP12A proton pump in cystic fibrosis airways. JCI Insight 2018, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gianotti, A.; Capurro, V.; Delpiano, L.; Mielczarek, M.; García-valverde, M.; Carreira-barral, I.; Ludovico, A.; Fiore, M.; Baroni, D.; Moran, O.; et al. Small molecule anion carriers correct abnormal airway surface liquid properties in cystic fibrosis airway epithelia. Int. J. Mol. Sci. 2020, 21, 1488. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Salinas, D.; Nielson, D.W.; Verkman, A.S. Hyperacidity of secreted fluid from submucosal glands in early cystic fibrosis. Am. J. Physiol. Cell Physiol. 2006, 290, 669–672. [Google Scholar] [CrossRef]
- Bodem, C.R.; Lampton, L.M.; Miller, D.P.; Tarka, E.F.; Everett, E.D. Endobronchial pH. Relevance to aminoglycoside activity in gram-negative bacillary pneumonia. Am. Rev. Respir. Dis. 1983, 127, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Karnad, D.R.; Mhaisekar, D.G.; Moralwar, K.V. Respiratory mucus pH in tracheostomized intensive care unit patients. Crit. Care Med. 1990, 18, 699–701. [Google Scholar] [CrossRef]
- Lozo Vukovac, E.; Miše, K.; Gudelj, I.; Perić, I.; Duplančić, D.; Vuković, I.; Vučinović, Z.; Lozo, M. Bronchoalveolar pH and inflammatory biomarkers in patients with acute exacerbation of chronic obstructive pulmonary disease. J. Int. Med. Res. 2019, 47, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kim, J.H.; Kim, S.W.; Kim, S.W.; Jin, K.S.; Cho, J.H.; Kang, J.M.; Park, S.Y. Nasal pH in patients with chronic rhinosinusitis before and after endoscopic sinus surgery. Am. J. Otolaryngol. Head Neck Med. Surg. 2013, 34, 505–507. [Google Scholar] [CrossRef]
- Masuda, M.; Sato, T.; Sakamaki, K.; Kudo, M.; Kaneko, T.; Ishigatsubo, Y. The effectiveness of sputum pH analysis in the prediction of response to therapy in patients with pulmonary tuberculosis. PeerJ 2015, 2015, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Adler, K.; Wooten, O.; Philippoff, W.; Lerner, E.; Dulfano, M.J. Physical Properties of Sputum. Am. Rev. Respir. Dis. 1972, 106, 86–96. [Google Scholar] [CrossRef] [PubMed]
- England, R.J.A.; Homer, J.J.; Knight, L.C.; Ell, S.R. Nasal pH measurement: A reliable and repeatable parameter. Clin. Otolaryngol. Allied Sci. 1999, 24, 67–68. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Huang, J.; Liao, J.; Robert, R.; Hanrahan, J.W. Anion secretion by a model epithelium: More lessons from Calu-3. Acta Physiol. 2011, 202, 523–531. [Google Scholar] [CrossRef]
- Saint-Criq, V.; Delpiano, L.; Casement, J.; Onuora, J.C.; Lin, J.H.; Gray, M.A. Choice of Differentiation Media Significantly Impacts Cell Lineage and Response to CFTR Modulators in Fully Differentiated Primary Cultures of Cystic Fibrosis Human Airway Epithelial Cells. Cells 2020, 9, 2137. [Google Scholar] [CrossRef] [PubMed]
- Hilding, A. The Common Cold. Arch. Otolaryngol. Head Neck Surg. 1930, 12, 133–150. [Google Scholar] [CrossRef]
- Steinmann, E. La secretion bronchique et le pH. Bronches 1956, 6, 126–129. [Google Scholar]
- Hunt, J.F.; Fang, K.; Malik, R.; Snyder, A.; Malhotra, N.; Platts-Mills, T.A.E.; Gaston, B. Endogenous Airway Acidification. Am. J. Respir. Crit. Care Med. 2000, 161, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Carraro, S.; Folesani, G.; Corradi, M.; Zanconato, S.; Gaston, B.; Baraldi, E. Acid-base equilibrium in exhaled breath condensate of allergic asthmatic children. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 476–481. [Google Scholar] [CrossRef]
- Kostikas, K.; Papatheodorou, G.; Ganas, K.; Psathakis, K.; Panagou, P.; Loukides, S. pH in expired breath condensate of patients with inflammatory airway diseases. Am. J. Respir. Crit. Care Med. 2002, 165, 1364–1370. [Google Scholar] [CrossRef]
- Antus, B.; Barta, I.; Kullmann, T.; Lazar, Z.; Valyon, M.; Horvath, I.; Csiszer, E. Assessment of exhaled breath condensate ph in exacerbations of asthma and chronic obstructive pulmonary disease: A longitudinal study. Am. J. Respir. Crit. Care Med. 2010, 182, 1492–1497. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, A.I.; Loukides, S.; Minas, M.; Kontogianni, K.; Bakakos, P.; Gourgoulianis, K.I.; Alchanatis, M.; Papiris, S.; Kostikas, K. Exhaled breath condensate pH as a biomarker of COPD severity in ex-smokers. Respir. Res. 2011, 12, 67. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.Y.; Hwang, P.H.; Illek, B.; Fischer, H. Acid and base secretion in freshly excised nasal tissue from cystic fibrosis patients with ΔF508 mutation. Int. Forum Allergy Rhinol. 2011, 1, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, P.S.; McCray, P.B.; Bals, R. The innate immune function of airway epithelial cells in inflammatory lung disease. Eur. Respir. J. 2015, 45, 1150–1162. [Google Scholar] [CrossRef] [Green Version]
- Ng, A.W.; Bidani, A.; Heming, T.A. Innate host defense of the lung: Effects of lung-lining fluid pH. Lung 2004, 182, 297–317. [Google Scholar] [CrossRef] [PubMed]
- Devor, D.C.; Singh, A.K.; Lambert, L.C.; DeLuca, A.; Frizzell, R.A.; Bridges, R.J. Bicarbonate and chloride secretion in Calu-3 human airway epithelial cells. J. Gen. Physiol. 1999, 113, 743–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, F.; Chen, J. Molecular structure of the ATP-bound, phosphorylated human CFTR. Proc. Natl. Acad. Sci. USA 2018, 115, 12757–12762. [Google Scholar] [CrossRef] [Green Version]
- Tabcharani, J.A.; Linsdell, P.; Hanrahan, J.W. Halide permeation in wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride channels. J. Gen. Physiol. 1997, 110, 341–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linsdell, P.; Evagelidis, A.; Hanrahan, J.W. Molecular determinants of anion selectivity in the cystic fibrosis transmembrane conductance regulator chloride channel pore. Biophys. J. 2000, 78, 2973–2982. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, J.H.; Fischer, H.; Illek, B.; Machen, T.E. Bicarbonate conductance and pH regulatory capability of cystic fibrosis transmembrane conductance regulator. Proc. Natl. Acad. Sci. USA 1994, 91, 5340–5344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linsdell, P.; Hanrahan, J.W. Glutathione permeability of CFTR. Am. J. Physiol. Cell Physiol. 1998, 275, 323–326. [Google Scholar] [CrossRef]
- Kreda, S.M.; Mall, M.; Mengos, A.; Rochelle, L.; Yankaskas, J.; Riordan, J.R.; Boucher, R.C. Characterization of wild-type and ΔF508 cystic fibrosis transmembrane regulator in human respiratory epithelia. Mol. Biol. Cell 2005, 16, 2154–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plasschaert, L.W.; Žilionis, R.; Choo-Wing, R.; Savova, V.; Knehr, J.; Roma, G.; Klein, A.M.; Jaffe, A.B. A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte. Nature 2018, 560, 377–381. [Google Scholar] [CrossRef]
- Okuda, K.; Dang, H.; Kobayashi, Y.; Carraro, G.; Nakano, S.; Chen, G.; Kato, T.; Asakura, T.; Gilmore, R.C.; Morton, L.C.; et al. Secretory Cells Dominate Airway CFTR Expression and Function in Human Airway Superficial Epithelia. Am. J. Respir. Crit. Care Med. 2020. [Google Scholar] [CrossRef]
- Bertrand, C.A.; Zhang, R.; Pilewski, J.M.; Frizzell, R.A. SLC26A9 is a constitutively active, CFTR-regulated anion conductance in human bronchial epithelia. J. Gen. Physiol. 2009, 133, 421–438. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Nam, J.H.; Park, H.W.; Oh, U.; Yoon, J.-H.; Lee, M.G. Dynamic modulation of ANO1/TMEM16A HCO3- permeability by Ca2+/calmodulin. Proc. Natl. Acad. Sci. USA 2013, 110, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Garnett, J.P.; Hickman, E.; Burrows, R.; Hegyi, P.; Tiszlavicz, L.; Cuthbert, A.W.; Fong, P.; Gray, M.A. Novel role for pendrin in orchestrating bicarbonate secretion in cystic fibrosis transmembrane conductance regulator (CFTR)-expressing airway serous cells. J. Biol. Chem. 2011, 286, 41069–41082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, S.B.H.; Zeng, W.; Dorwart, M.R.; Luo, X.; Kim, K.H.; Millen, L.; Goto, H.; Naruse, S.; Soyombo, A.; Thomas, P.J.; et al. Gating of CFTR by the STAS domain of SLC26 transporters. Nat. Cell Biol. 2004, 6, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Bakouh, N.; Bienvenu, T.; Thomas, A.; Ehrenfeld, J.; Liote, H.; Roussel, D.; Duquesnoy, P.; Farman, N.; Viel, M.; Cherif-Zahar, B.; et al. Characterization of SLC26A9 in patients with CF-like lung disease. Hum. Mutat. 2013, 34, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Rode, B.; Dirami, T.; Bakouh, N.; Rizk-rabin, M.; Norez, C.; Lhuillier, P.; Lorès, P.; Jollivet, M.; Melin, P.; Zvetkova, I.; et al. The testis anion transporter TAT1 (SLC26A8) physically and functionally interacts with the cystic fibrosis transmembrane conductance regulator channel: A potential role during sperm capacitation. Hum. Mol. Genet. 2012, 21, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Lee, M.G.; Ko, S.; Muallem, S. Cl--dependent HCO3- transport by cystic fibrosis transmembrane conductance regulator. J. Pancreas 2001, 2, 243–246. [Google Scholar]
- Massey, M.K.; Reiterman, M.J.; Mourad, J.; Luckie, D.B. Is CFTR an exchanger?: Regulation of HCO3−Transport and extracellular pH by CFTR. Biochem. Biophys. Rep. 2021, 25, 100863. [Google Scholar] [CrossRef]
- Chen, J.H.; Cai, Z.; Sheppard, D.N. Direct sensing of intracellular pH by the cystic fibrosis transmembrane conductance regulator (CFTR) Cl- channel. J. Biol. Chem. 2009, 284, 35495–35506. [Google Scholar] [CrossRef] [Green Version]
- Park, H.W.; Nam, J.H.; Kim, J.Y.; Namkung, W.; Yoon, J.S.; Lee, J.S.; Kim, K.S.; Venglovecz, V.; Gray, M.A.; Kim, K.H.; et al. Dynamic regulation of CFTR bicarbonate permeability by [Cl−]i and its role in pancreatic bicarbonate secretion. Gastroenterology 2010, 139, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Jun, I.; Shin, D.H.; Yoon, J.G.; Piao, H.; Jung, J.; Park, H.W.; Cheng, M.H.; Bahar, I.; Whitcomb, D.C.; et al. Regulation of CFTR Bicarbonate Channel Activity by WNK1: Implications for Pancreatitis and CFTR-Related Disorders. Cmgh 2020, 9, 79–103. [Google Scholar] [CrossRef] [Green Version]
- The Human Protein Atlas. Available online: v14.proteinatlas.org/ENSG00000060237-WNK1/tissue/lung (accessed on 13 January 2021).
- Choi, J.Y.; Muallem, D.; Kiselyov, K.; Lee, M.G.; Thomas, P.J.; Muallem, S. Aberrant CFTR-dependent HCO3- transport in mutations associated with cystic fibrosis. Nature 2001, 410, 94–97. [Google Scholar] [CrossRef] [Green Version]
- LaRusch, J.; Jung, J.; General, I.J.; Lewis, M.D.; Park, H.W.; Brand, R.E.; Gelrud, A.; Anderson, M.A.; Banks, P.A.; Conwell, D.; et al. Mechanisms of CFTR Functional Variants That Impair Regulated Bicarbonate Permeation and Increase Risk for Pancreatitis but Not for Cystic Fibrosis. PLoS Genet. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, M.J.; Clarke, L.L.; Gray, M.A. How to measure CFTR-dependent bicarbonate transport: From single channels to the intact epithelium. Methods Mol. Biol. 2011, 741, 489–509. [Google Scholar] [CrossRef]
- Fiore, M.; Picco, C.; Moran, O. Correctors modify the bicarbonate permeability of F508del-CFTR. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lérias, J.; Pinto, M.; Benedetto, R.; Schreiber, R.; Amaral, M.; Aureli, M.; Kunzelmann, K. Compartmentalized crosstalk of CFTR and TMEM16A (ANO1) through EPAC1 and ADCY1. Cell. Signal. 2018, 44, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Geertsma, E.R.; Chang, Y.N.; Shaik, F.R.; Neldner, Y.; Pardon, E.; Steyaert, J.; Dutzler, R. Structure of a prokaryotic fumarate transporter reveals the architecture of the SLC26 family. Nat. Struct. Mol. Biol. 2015, 22, 803–808. [Google Scholar] [CrossRef]
- Gray, M.A. Bicarbonate secretion: It takes two to tango. Nat. Cell Biol. 2004, 6, 292–294. [Google Scholar] [CrossRef]
- Shcheynikov, N.; Wang, Y.; Park, M.; Ko, S.B.H.; Dorwart, M.; Naruse, S.; Thomas, P.J.; Muallem, S. Coupling modes and stoichiometry of Cl−/HCO3− exchange by slc26a3 and slc26a6. J. Gen. Physiol. 2006, 127, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheat, V.J.; Shumaker, H.; Burnham, C.; Shull, G.E.; Yankaskas, J.R.; Soleimani, M. CFTR induces the expression of DRA along with Cl-/HCO3- exchange activity in tracheal epithelial cells. Am. J. Physiol. Cell Physiol. 2000, 279, 62–71. [Google Scholar] [CrossRef]
- Rehman, T.; Thornell, I.M.; Pezzulo, A.A.; Thurman, A.L.; Romano Ibarra, G.S.; Karp, P.H.; Tan, P.; Duffey, M.E.; Welsh, M.J. TNFα and IL-17 Alkalinize Airway Surface Liquid through CFTR and Pendrin. Am. J. Physiol. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Pedemonte, N.; Caci, E.; Sondo, E.; Caputo, A.; Rhoden, K.; Pfeffer, U.; Di Candia, M.; Bandettini, R.; Ravazzolo, R.; Zegarra-Moran, O.; et al. Thiocyanate Transport in Resting and IL-4-Stimulated Human Bronchial Epithelial Cells: Role of Pendrin and Anion Channels. J. Immunol. 2007, 178, 5144–5153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, K.M.; Abraham, V.; Spielman, D.; Kolls, J.K.; Rubenstein, R.C.; Conner, G.E.; Cohen, N.A.; Kreindler, J.L. IL-17A induces Pendrin expression and chloride-bicarbonate exchange in human bronchial epithelial cells. PLoS ONE 2014, 9, e103263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnett, J.P.; Turner, M.J. Controversies surrounding the role of CFTR in airway bicarbonate secretion. J. Physiol. 2013, 591, 2241–2242. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Soyombo, A.A.; Shcheynikov, N.; Zeng, W.; Dorwart, M.; Marino, C.R.; Thomas, P.J.; Muallem, S. Slc26a6 regulates CFTR activity in vivo to determine pancreatic duct HCO3- secretion: Relevance to cystic fibrosis. EMBO J. 2006, 25, 5049–5057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Valentin, E.; Crahay, C.; Garbacki, N.; Hennuy, B.; Guéders, M.; Noël, A.; Foidart, J.M.; Grooten, J.; Colige, A.; Piette, J.; et al. New asthma biomarkers: Lessons from murine models of acute and chronic asthma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Ohta, N.; Suzuki, Y.; Kakehata, S.; Okubo, K.; Ikeda, H.; Shiraishi, H.; Izuhara, K. Expression of pendrin and periostin in allergic rhinitis and chronic rhinosinusitis. Allergol. Int. 2012, 61, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Nakagami, Y.; Favoreto, S.; Zhen, G.; Park, S.-W.; Nguyenvu, L.T.; Kuperman, D.A.; Dolganov, G.M.; Huang, X.; Boushey, H.A.; Avila, P.C.; et al. The Epithelial Anion Transporter Pendrin Is Induced by Allergy and Rhinovirus Infection, Regulates Airway Surface Liquid, and Increases Airway Reactivity and Inflammation in an Asthma Model. J. Immunol. 2008, 181, 2203–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajko, J.; Duguid, M.; Altmann, S.; Hurlbut, G.D.; Kaczmarek, J.S. Pendrin stimulates a chloride absorption pathway to increase CFTR-mediated chloride secretion from Cystic Fibrosis airway epithelia. FASEB BioAdvances 2020. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Huang, J.; Billet, A.; Abu-Arish, A.; Goepp, J.; Matthes, E.; Tewfik, M.A.; Frenkiel, S.; Hanrahan, J.W. Pendrin mediates bicarbonate secretion and enhances cystic fibrosis transmembrane conductance regulator function in airway surface epithelia. Am. J. Respir. Cell Mol. Biol. 2019, 60, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, C.A.; Mitra, S.; Mishra, S.K.; Wang, X.; Zhao, Y.; Pilewski, J.M.; Madden, D.R.; Frizzell, R.A. The CFTR trafficking mutation F508del inhibits the constitutive activity of SLC26A9. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L12–L925. [Google Scholar] [CrossRef] [PubMed]
- Lohi, H.; Kujala, M.; Mäkelä, S.; Lehtonen, E.; Kestilä, M.; Saarialho-Kere, U.; Markovichand, D.; Kere, J. Functional characterization of three novel tissue-specific anion exchangers SLC26A7, -A8, and -A9. J. Biol. Chem. 2002, 277, 14246–14254. [Google Scholar] [CrossRef] [Green Version]
- Salomon, J.J.; Spahn, S.; Wang, X.; Füllekrug, J.; Bertrand, C.A.; Mall, M.A. Generation and functional characterization of epithelial cells with stable expression of SLC26A9 Cl- channels. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L593–L602. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Henriksnäs, J.; Barone, S.; Witte, D.; Shull, G.E.; Forte, J.G.; Holm, L.; Soleimani, M. SLC26A9 is expressed in gastric surface epithelial cells, mediates Cl-/HCO3- exchange, and is inhibited by NH4+. Am. J. Physiol. Cell Physiol. 2005, 289, C493–C505. [Google Scholar] [CrossRef]
- Demitrack, E.S.; Soleimani, M.; Montrose, M.H. Damage to the gastric epithelium activates cellular bicarbonate secretion via SLC26A9 Cl-/HCO3- exchange. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G255–G264. [Google Scholar] [CrossRef] [PubMed]
- Dorwart, M.R.; Shcheynikov, N.; Wang, Y.; Stippec, S.; Muallem, S. SLC26A9 is a Cl- channel regulated by the WNK kinases. J. Physiol. 2007, 584, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-H.H.; Plata, C.; Zandi-Nejad, K.; Sinđić, A.; Sussman, C.R.; Mercado, A.; Broumand, V.; Raghuram, V.; Mount, D.B.; Romero, M.F.; et al. Slc26a9-Anion exchanger, channel and Na+ transporter. J. Membr. Biol. 2009, 228, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Alper, S.L. Molecular physiology and genetics of Na+-independent SLC4 anion exchangers. J. Exp. Biol. 2009, 212, 1672–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, A.K.; Chernova, M.N.; Shmukler, B.E.; Wilhelm, S.; Alper, S.L. Regulation of AE2-mediated Cl- transport by intracellular or by extracellular pH requires highly conserved amino acid residues of the AE2 NH2-terminal cytoplasmic domain. J. Gen. Physiol. 2002, 120, 707–722. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Shan, J.; Kim, D.; Liao, J.; Evagelidis, A.; Alper, S.L.; Hanrahan, J.W. Basolateral chloride loading by the anion exchanger type 2: Role in fluid secretion by the human airway epithelial cell line Calu-3. J. Physiol. 2012, 590, 5299–5316. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.H.; Turner, M.J.; Saint-Criq, V.; Garnett, J.; Haq, I.J.; Brodlie, M.; Ward, C.; Borgo, C.; Salvi, M.; Venerando, A.; et al. CK2 is a key regulator of SLC4A2-mediated Cl−/HCO3− exchange in human airway epithelia. Pflugers Arch. Eur. J. Physiol. 2017, 469, 1073–1091. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.D.; Boron, W.F. The divergence, actions, roles, and relatives of sodium-coupled bicarbonate transporters. Physiol. Rev. 2013, 93, 803–959. [Google Scholar] [CrossRef] [Green Version]
- Kreindler, J.L.; Peters, K.W.; Frizzell, R.A.; Bridges, R.J. Identification and membrane localization of electrogenic sodium bicarbonate cotransporters in Calu-3 cells. Biochim. Biophys. Acta Mol. Basis Dis. 2006, 1762, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Pushkin, A.; Abuladze, N.; Newman, D.; Lee, I.; Xu, G.; Kurtz, I. Two C-terminal variants of NBC4, a new member of the sodium bicarbonate cotransporter family: Cloning, characterization, and localization. IUBMB Life 2000, 50, 13–19. [Google Scholar] [CrossRef]
- Barbuskaite, D.; Pedersen, F.D.; Christensen, H.L.; Johnsen, L.; Praetorius, J.; Damkier, H.H. NBCe2 (Slc4a5) Is Expressed in the Renal Connecting Tubules and Cortical Collecting Ducts and Mediates Base Extrusion. Front. Physiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Shirakabe, K.; Priori, G.; Yamada, H.; Ando, H.; Horita, S.; Fujita, T.; Fujimoto, I.; Mizutani, A.; Seki, G.; Mikoshiba, K. IRBIT, an inositol 1,4,5-trisphosphate receptor-binding protein, specifically binds to and activates pancreas-type Na+/HCO3- cotransporter 1 (pNBC1). Proc. Natl. Acad. Sci. USA 2006, 103, 9542–9547. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Shcheynikov, N.; Zeng, W.; Ohana, E.; So, I.; Ando, H.; Mizutani, A.; Mikoshiba, K.; Muallem, S. IRBIT coordinates epithelial fluid and HCO3- secretion by stimulating the transporters pNBC1 and CFTR in the murine pancreatic duct. J. Clin. Investig. 2009, 119, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shcheynikov, N.; Son, A.; Hong, J.H.; Yamazaki, O.; Ohana, E.; Kurtz, I.; Shin, D.M.; Muallem, S. Intracellular Cl-as a signaling ion that potently regulates Na+/Hco3—Transporters. Proc. Natl. Acad. Sci. USA 2015, 112, E329–E337. [Google Scholar] [CrossRef] [Green Version]
- Effros, R.; Chang, R.; Silverman, P. Acceleration of plasma bicarbonate conversion to carbon dioxide by pulmonary carbonic anhydrase. Science 1978, 199, 427–429. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.P.; Alt, M.D.; Klier, M.; Spiess, A.; Andes, F.T.; Waheed, A.; Sly, W.S.; Becker, H.M.; Deitmer, J.W. GPI-anchored carbonic anhydrase IV displays both intra- and extracellular activity in cRNA-injected oocytes and in mouse neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 1494–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vince, J.W.; Reithmeier, R.A.F. Carbonic anhydrase II binds to the carboxyl terminus of human band 3, the erythrocyte Cl-/HCO3- exchanger. J. Biol. Chem. 1998, 273, 28430–28437. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Alvarez, B.; Casey, J.R.; Reithmeier, R.A.F.; Fliegel, L. Carbonic anhydrase II binds to and enhances activity of the Na+/H+ exchanger. J. Biol. Chem. 2002, 277, 36085–36091. [Google Scholar] [CrossRef] [Green Version]
- Pushkin, A.; Kurtz, I. SLC4 base (HCO3−, CO32−) transporters: Classification, function, structure, genetic diseases, and knockout models. Am. J. Physiol. Ren. Physiol. 2006, 290, 580–599. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.M.; Deitmer, J.W. Carbonic anhydrase II increases the activity of the human electrogenic Na+/HCO3- cotransporter. J. Biol. Chem. 2007, 282, 13508–13521. [Google Scholar] [CrossRef] [Green Version]
- Inglis, S.K.; Finlay, L.; Ramminger, S.J.; Richard, K.; Ward, M.R.; Wilson, S.M.; Olver, R.E. Regulation of intracellular pH in Calu-3 human airway cells. J. Physiol. 2002, 538, 527–539. [Google Scholar] [CrossRef]
- Effros, R.M. Carbonic anhydrase and alveolar fluid absorption. Am. J. Respir. Cell Mol. Biol. 2008, 39, 124. [Google Scholar] [CrossRef]
- Thornell, I.M.; Li, X.; Tang, X.X.; Brommel, C.M.; Karp, P.H.; Welsh, M.J.; Zabner, J. Nominal carbonic anhydrase activity minimizes airway-surface liquid pH changes during breathing. Physiol. Rep. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanjul, M.; Salvador, C.; Alvarez, L.; Cantet, S.; Hollande, E. Targeting of carbonic anhydrase IV to plasma membrane is altered in cultured human pancreatic duct cells expressing a mutated (ΔF508) CFTR. Eur. J. Cell Biol. 2002, 81, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Vecchio-Pagán, B.; Sharma, N.; Waheed, A.; Li, X.; Raraigh, K.S.; Robbins, S.; Han, S.T.; Franca, A.L.; Pellicore, M.J.; et al. Loss of carbonic anhydrase XII function in individuals with elevated sweat chloride concentration and pulmonary airway disease. Hum. Mol. Genet. 2016, 25, 1923–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.X.; Fok, K.L.; Chen, H.; Chan, K.S.; Tsang, L.L.; Rowlands, D.K.; Zhang, X.H.; Da Dong, J.; Ruan, Y.C.; Jiang, X.; et al. Lymphocyte CFTR promotes epithelial bicarbonate secretion for bacterial killing. J. Cell. Physiol. 2012, 227, 3887–3894. [Google Scholar] [CrossRef] [PubMed]
- Coyne, C.B.; Vanhook, M.K.; Gambling, T.M.; Carson, J.L.; Boucher, R.C.; Johnson, L.G. Regulation of Airway Tight Junctions by Proinflammatory Cytokines. Mol. Biol. Cell 2002, 13, 3218–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinhas, R.; Cortes, L.; Cardoso, I.; Mendes, V.M.; Manadas, B.; Todo-Bom, A.; Pires, E.; Veríssimo, P. Pollen proteases compromise the airway epithelial barrier through degradation of transmembrane adhesion proteins and lung bioactive peptides. Allergy Eur. J. Allergy Clin. Immunol. 2011, 66, 1088–1098. [Google Scholar] [CrossRef]
- Günzel, D.; Yu, A.S.L. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, A.N.; Itani, O.A.; Moninger, T.O.; Welsh, M.J. Acute regulation of tight junction ion selectivity in human airway epithelia. Proc. Natl. Acad. Sci. USA 2009, 106, 3591–3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornell, I.M.; Rehman, T.; Pezzulo, A.A.; Welsh, M.J. Paracellular bicarbonate flux across human cystic fibrosis airway epithelia tempers changes in airway surface liquid pH. J. Physiol. 2020, JP280120. [Google Scholar] [CrossRef]
- Fischer, H.; Widdicombe, J.H. Mechanisms of acid and base secretion by the airway epithelium. J. Membr. Biol. 2006, 211, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Aronson, P.S.; Nee, J.; Suhm, M.A. Modifier role of internal H+ in activating the Na +-H+ exchanger in renal microvillus membrane vesicles. Nature 1982, 299, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Al-Bazzaz, F.J.; Hafez, N.; Tyagi, S.; Gailey, C.A.; Toofanfard, M.; Alrefai, W.A.; Nazir, T.M.; Ramaswamy, K.; Dudeja, P.K. Detection of Cl--HCO3- and Na+-H+ exchangers in human airways epithelium. J. Pancreas 2001, 2, 285–290. [Google Scholar]
- Paradiso, A.M. ATP-activated baselateral Na+/H+ exchange in human normal and cystic fibrosis airway epithelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 1997, 273. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, M.; Steele, L. Na(+)−H+ exchanger in isolated epithelial tracheal cells from sheep. Involvement in tracheal proton secretion. Exp. Physiol. 1993, 78, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Willumsen, N.J.; Boucher, R.C. Intracellular pH and its relationship to regulation of ion transport in normal and cystic fibrosis human nasal epithelia. J. Physiol. 1992, 455, 247–269. [Google Scholar] [CrossRef]
- Fischer, H.; Widdicombe, J.H.; Illek, B. Acid secretion and proton conductance in human airway epithelium. Am. J. Physiol. Cell Physiol. 2002, 282, 736–743. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.V.N.; Ribeiro, J.D.; Bertuzzo, C.S.; Marson, F.A.L. Association of clinical severity of cystic fibrosis with variants in the SLC gene family (SLC6A14, SLC26A9, SLC11A1 and SLC9A3). Gene 2017, 629, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, R.; Taylor, C.; Lin, F.; Sun, L.; Sandford, A.; Paré, P.; Berthiaume, Y.; Corey, M.; Durie, P.; Zielenski, J. Modulatory effect of the SLC9A3 gene on susceptibility to infections and pulmonary function in children with cystic fibrosis. Pediatr. Pulmonol. 2011, 46, 385–392. [Google Scholar] [CrossRef]
- Corvol, H.; Blackman, S.M.; Boëlle, P.Y.; Gallins, P.J.; Pace, R.G.; Stonebraker, J.R.; Accurso, F.J.; Clement, A.; Collaco, J.M.; Dang, H.; et al. Genome-wide association meta-analysis identifies five modifier loci of lung disease severity in cystic fibrosis. Nat. Commun. 2015, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Soave, D.; Miller, M.R.; Keenan, K.; Lin, F.; Gong, J.; Chiang, T.; Stephenson, A.L.; Durie, P.; Rommens, J.; et al. Unraveling the complex genetic model for cystic fibrosis: Pleiotropic effects of modifier genes on early cystic fibrosis-related morbidities. Hum. Genet. 2014, 133, 151–161. [Google Scholar] [CrossRef]
- Cougnon, M.; Bouyer, P.; Planelles, G.; Jaisser, F. Does the colonic H,K-ATPase also act as an Na,K-ATPase? Proc. Natl. Acad. Sci. 1998, 95, 6516–6520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnay, M.; Crambert, G.; Kharoubi-Hess, S.; Geering, K.; Horisberger, J.D. Bufo marinus bladder H-K-ATPase carries out electroneutral ion transport. Am. J. Physiol. Ren. Physiol. 2001, 281, 869–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishin, A.V.; Caplan, M.J. ATP1AL1, a member of the non-gastric H,K-ATPase family, functions as a sodium pump. J. Biol. Chem. 1998, 273, 27772–27778. [Google Scholar] [CrossRef] [Green Version]
- Gorrieri, G.; Scudieri, P.; Caci, E.; Schiavon, M.; Tomati, V.; Sirci, F.; Napolitano, F.; Carrella, D.; Gianotti, A.; Musante, I.; et al. Goblet Cell Hyperplasia Requires High Bicarbonate Transport to Support Mucin Release. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroche-Joubert, N.; Marsy, S.; Luriau, S.; Imbert-Teboul, M.; Doucet, A. Mechanism of activation of ERK and H-K-ATPase by isoproterenol in rat cortical collecting duct. Am. J. Physiol. Ren. Physiol. 2003, 284, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherny, V.V.; Markin, V.S.; Decoursey, T.E. The Voltage-activated hydrogen ion conductance in rat alveolar epithelial cells is determined by the pH gradient. J. Gen. Physiol. 1995, 105, 861–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, C.; Machen, T.E.; Illek, B.; Fischer, H. NADPH oxidase-dependent acid production in airway epithelial cells. J. Biol. Chem. 2004, 279, 36454–36461. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.Y.; Hajighasemi, M.; Hwang, P.H.; Illek, B.; Fischer, H. Proton secretion in freshly excised sinonasal mucosa from asthma and sinusitis patients. Am. J. Rhinol. Allergy 2009, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Iovannisci, D.; Illek, B.; Fischer, H. Function of the HVCN1 proton channel in airway epithelia and a naturally occurring mutation, M91T. J. Gen. Physiol. 2010, 136, 35–46. [Google Scholar] [CrossRef]
- Ribeiro, C.M.P.; Paradiso, A.M.; Livraghi, A.; Boucher, R.C. The mitochondrial barriers segregate agonist-induced calcium-dependent functions in human airway epithelia. J. Gen. Physiol. 2003, 122, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Fischer, H. Function of proton channels in lung epithelia. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.D.; Abdul Rahman, R.; Khalid, F.; Cotton, C.; Smith, L.; Boyne, K.; Marozkina, N.; Ramsey, I.S.; Chmiel, J.; Gaston, B.M. Airway Epithelial Voltage-Gated Proton Channel Expression and pH in Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2020, 201, A7452. [Google Scholar] [CrossRef]
- Futai, M.; Nakanishi-Matsui, M.; Okamoto, H.; Sekiya, M.; Nakamoto, R.K. Rotational catalysis in proton pumping ATPases: From E. coli F-ATPase to mammalian V-ATPase. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1711–1721. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Thornell, I.; Villacreses Rada, R.; Brommel, C.; Lu, L.; Mather, S.; Ehler, A.; Karp, P.; Welsh, M.; Zabner, J. V-Type ATPase Mediates Airway Surface Liquid Acidification in Cultured Pig Small Airway Epithelial Cells. Am. J. Respir. Crit. Care Med. 2019, 199, A2573. [Google Scholar] [CrossRef]
- Poulsen, J.H.; Machen, T.E. HCO3-dependent pH(i) regulation in tracheal epithelial cells. Pflugers Arch. Eur. J. Physiol. 1996, 432, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Young, L.; Chen, Y.; Ran, H.; Meyers, M.; Joseph, P.; Cho, Y.H.; Hassett, D.J.; Lau, G.W. Pseudomonas aeruginosa pyocyanin inactivates lung epithelial vacuolar ATPase-dependent cystic fibrosis transmembrane conductance regulator expression and localization. Cell. Microbiol. 2006, 8, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.; Flemström, G. Gastroduodenal mucus bicarbonate barrier: Protection against acid and pepsin. Am. J. Physiol. Cell Physiol. 2005, 288, C1–C19. [Google Scholar] [CrossRef] [Green Version]
- Holma, B.; Hegg, P.O. pH- and protein-dependent buffer capacity and viscosity of respiratory mucus. Their interrelationships and influence of health. Sci. Total Environ. 1989, 84, 71–82. [Google Scholar] [CrossRef]
- Holma, B. Influence of buffer capacity and pH-dependent rheological properties of respiratory mucus on health effects due to acidic pollution. Sci. Total Environ. 1985, 41, 101–123. [Google Scholar] [CrossRef]
- Ballard, S.T.; Trout, L.; Bebök, Z.; Sorscher, E.J.; Crews, A. CFTR involvement in chloride, bicarbonate, and liquid secretion by airway submucosal glands. Am. J. Physiol. Lung Cell. Mol. Physiol. 1999, 277, 3–8. [Google Scholar] [CrossRef]
- Cho, H.J.; Joo, N.S.; Wine, J.J. Mucus secretion from individual submucosal glands of the ferret trachea. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, N.S.; Cho, H.J.; Khansaheb, M.; Wine, J.J. Hyposecretion of fluid from tracheal submucosal glands of CFTR-deficient pigs. J. Clin. Investig. 2010, 120, 3161–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, J.F.; Yankaskas, J.R.; Ernst, S.A.; Yang, Y.; Marino, C.R.; Boucher, R.C.; Cohn, J.A.; Wilson, J.M. Submucosal glands are the predominant site of CFTR expression in the human bronchus. Nat. Genet. 1992, 2, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Ratzke, C.; Gore, J. Modifying and reacting to the environmental pH can drive bacterial interactions. PLoS Biol. 2018, 16, e2004248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, R.A.; Comstock, W.; Zhang, T.; Morton, J.T.; Da Silva, R.; Tran, A.; Aksenov, A.; Nothias, L.F.; Wangpraseurt, D.; Melnik, A.V.; et al. Niche partitioning of a pathogenic microbiome driven by chemical gradients. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.J.; Welsh, M.J. cAMP stimulates bicarbonate secretion across normal, but not cystic fibrosis airway epithelia. J. Clin. Investig. 1992, 89, 1148–1153. [Google Scholar] [CrossRef]
- Liu, X.; Li, T.; Tuo, B. Physiological and pathophysiological relevance of the anion transporter Slc26a9 in multiple organs. Front. Physiol. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Strug, L.J.; Gonska, T.; He, G.; Keenan, K.; Ip, W.; Böelle, P.Y.; Lin, F.; Panjwani, N.; Gong, J.; Li, W.; et al. Cystic fibrosis gene modifier SLC26A9 modulates airway response to CFTR-directed therapeutics. Hum. Mol. Genet. 2016, 25, 4590–4600. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, I.; Petrasek, D.; Tatishchev, S. Molecular Mechanisms of Electrogenic Sodium Bicarbonate Cotransport: Structural and Equilibrium Thermodynamic Considerations. J. Membr. Biol. 2004, 197, 77–90. [Google Scholar] [CrossRef]
- Morla, L.; Doucet, A.; Lamouroux, C.; Crambert, G.; Edwards, A. The renal cortical collecting duct: A secreting epithelium? J. Physiol. 2016, 594, 5991–6008. [Google Scholar] [CrossRef] [Green Version]
- Cheval, L.; Bakouh, N.; Walter, C.; Tembely, D.; Morla, L.; Escher, G.; Vogt, B.; Crambert, G.; Planelles, G.; Doucet, A. ANP-stimulated Na+ secretion in the collecting duct prevents Na+ retention in the renal adaptation to acid load. Am. J. Physiol. Ren. Physiol. 2019, 317, F435–F443. [Google Scholar] [CrossRef] [PubMed]
- Crambert, G. H-K-ATpase type 2: Relevance for renal physiology and beyond. Am. J. Physiol. Ren. Physiol. 2014, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, F.J.; Boron, W.F. Carbonic anhydrases enhance activity of endogenous Na-H exchangers and not the electrogenic Na/HCO3 cotransporter NBCe1-A, expressed in Xenopus oocytes. J. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample/Model | pH Value | Method Reference | ||
|---|---|---|---|---|
| WT | CF | |||
| Mouse | 6.95 ± 0.03 | 6.84 ± 0.07 | BCECF-dextran | [51] |
| 7.14 ± 0.01 (in vivo) | BCECF-dextran | [52] | ||
| 6.98 ± 0.16 | pH electrode | [50] | ||
| 7.28 | BCECF-dextran | [8] | ||
| Rat | 7.25 ± 0.05 | 6.42 ± 0.12 | Not specified | [53] |
| Ferret | 6.84 ± 0.03 | pH electrode | [54] | |
| Rabbit * | 6.92 ± 0.01 | pH electrode | [55] | |
| Cow | 6.81 ± 0.04 (25 mM bicarbonate) | BCECF-dextran | [52] | |
| 6.98 ± 0.05 (no bicarbonate) | BCECF-dextran | [52] | ||
| Pig | 7.14 ± 0.04 | 6.94 ± 0.05 | Optode (in vivo) | [22] |
| Primary Bronchial epithelia | 7.37 ± 0.05 | 7.05 ± 0.03 | SNARF pH indicator | [22] |
| 6.93 ± 0.04 | pH electrode | [56] | ||
| ~7.1 | 7.2 | pH electrode | [50] | |
| Gland fluid | 6.9 ± 0.06 | BCECF-dextran | [57] | |
| Sample/Model | pH Value | Method | Reference | |
|---|---|---|---|---|
| WT | CF | |||
| Cell lines | ||||
| CFBE41σ | 7.42 ± 0.02 * | 7.15 ± 0.01 * | pH electrode | [58] |
| 7.24 ** | X Ray microanalysis | [59] | ||
| 16HBE14 σ | 7.14 ± 0.02 | pH electrode | [60] | |
| 7.16 ** | pH electrode | [59] | ||
| Calu3 | ~7.2 | BCECF-dextran | [61] | |
| 7.55 ± 0.04 | 7.28 ± 0.02 | pH electrode | [62] | |
| NuLi-1/CuFi-1 | 7.52 ± 0.07 | 6.88 ± 0.02 | SNARF pH indicator | [63] |
| C38/IB3-1/ | 7.32 ± 0.08 | 7.02 ± 0.04 | pH electrode | [64] |
| Primary cells (bronchi) | ||||
| 6.81 ± 0.20 | BCECF-dextran | [65] | ||
| 6.6 ± 0.1 | SNARF-1 | [66] | ||
| ΔpH = −0.096 ± 0.029 * | ΔpH = −0.146 ± 0.011 * | pH electrode | [42] | |
| ~7.4 | ~7.1 | BCECF-dextran | [67] | |
| 7.77 | 7.31 | pH electrode | [68] | |
| 7.35 ± 0.09 | pH electrode | [43] | ||
| 7.43 ± 0.06 * | 7.26 ± 0.02 * | pH electrode | [58] | |
| 7.35 ± 0.05 | 6.70 ± 0.03 | pH electrode | [69] | |
| Submucosal gland secretions | ||||
| 7.18 ± 0.06 | 6.57 ± 0.09 | BCECF-dextran | [70] | |
| 6.97 ± 0.06 | BCECF-dextran | [65] | ||
| Sample/Model | pH Value | Method | Reference | |
|---|---|---|---|---|
| WT | CF | |||
| Nose | ||||
| Edge of nostril/adults | 5.5 ± 0.1 | 5.6 ± 0.1 | Monocrystalline antimony catheter | [47] |
| 4 cm from nares/adults | 6.7 ± 0.13 | 6.2 ± 0.1 | Monocrystalline antimony catheter | [47] |
| 6.6 ± 0.1 | 6.8 ± 0.10 | Gold probe | [47] | |
| 5 to 7.2 * | Mobidium pH probe | [44] | ||
| Neonates | 6.4 ± 0.2 | 5.2 ± 0.3 (4.5–6.9) | Mobidium pH probe | [44,45] |
| Lower airway/children | ||||
| 7.1 ± 0.1 | 7.1 ± 0.2 | Gold probe | [47] | |
| 7.00 ± 0.12 | 6.98 ± 0.15 | Fiberoptic probe | [48] | |
| pH Value | Method | Reference | |
|---|---|---|---|
| Pneumonia | 6.62 ± 0.07 | pH electrode | [71] |
| 6.72 | pH electrode | [72] | |
| Chronic lung diseases | 6.64 ± 0.08 | pH electrode | [71] |
| COPD | 6.21 ± 0.37 | pH test strip | [73] |
| Acute exacerbation of COPD (AECOPD) | 6.89 ± 0.53 | pH test strip | [73] |
| Chronic rhinosinusitis | 6.7 ± 0.6 | pH electrode | [74] |
| Pulmonary tuberculosis (sputum) | 7.00 (range 5.50–8.37) | pH electrode | [75] |
| Chronic bronchitis | 7.59 (mucoid) | pH electrode | [76] |
| 7.83 (purulent) | pH electrode | [76] | |
| Rhinitis | 7.2–8.3 | pH electrode | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajac, M.; Dreano, E.; Edwards, A.; Planelles, G.; Sermet-Gaudelus, I. Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge. Int. J. Mol. Sci. 2021, 22, 3384. https://doi.org/10.3390/ijms22073384
Zajac M, Dreano E, Edwards A, Planelles G, Sermet-Gaudelus I. Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge. International Journal of Molecular Sciences. 2021; 22(7):3384. https://doi.org/10.3390/ijms22073384
Chicago/Turabian StyleZajac, Miroslaw, Elise Dreano, Aurelie Edwards, Gabrielle Planelles, and Isabelle Sermet-Gaudelus. 2021. "Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge" International Journal of Molecular Sciences 22, no. 7: 3384. https://doi.org/10.3390/ijms22073384