
High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology

 , ,
, ,
Abstract
:1. Introduction
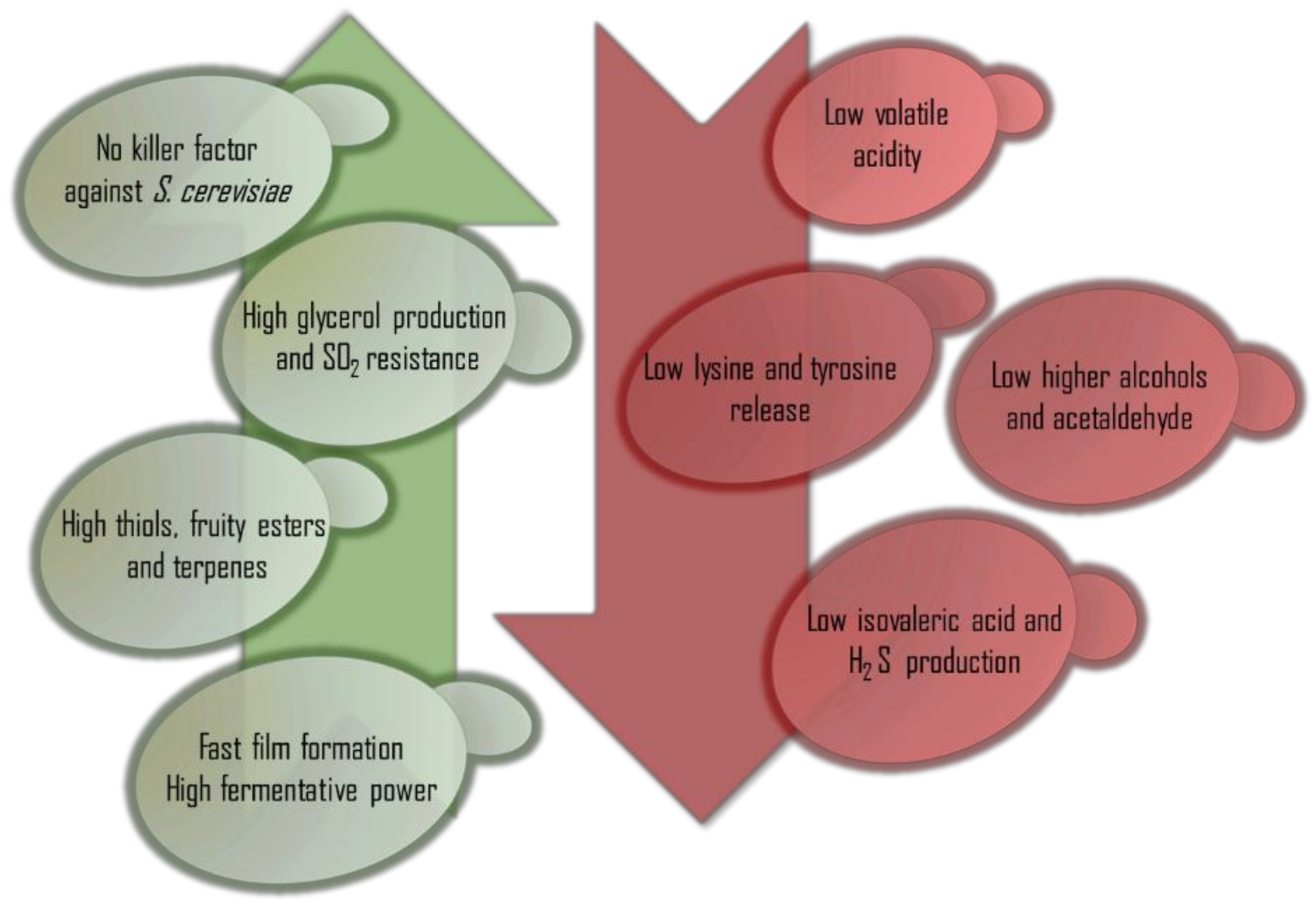
2. P. kluyveri Impact on Different Wine Quality Parameters
2.1. Ethanol
2.2. Glycerol
2.3. Organic Acids
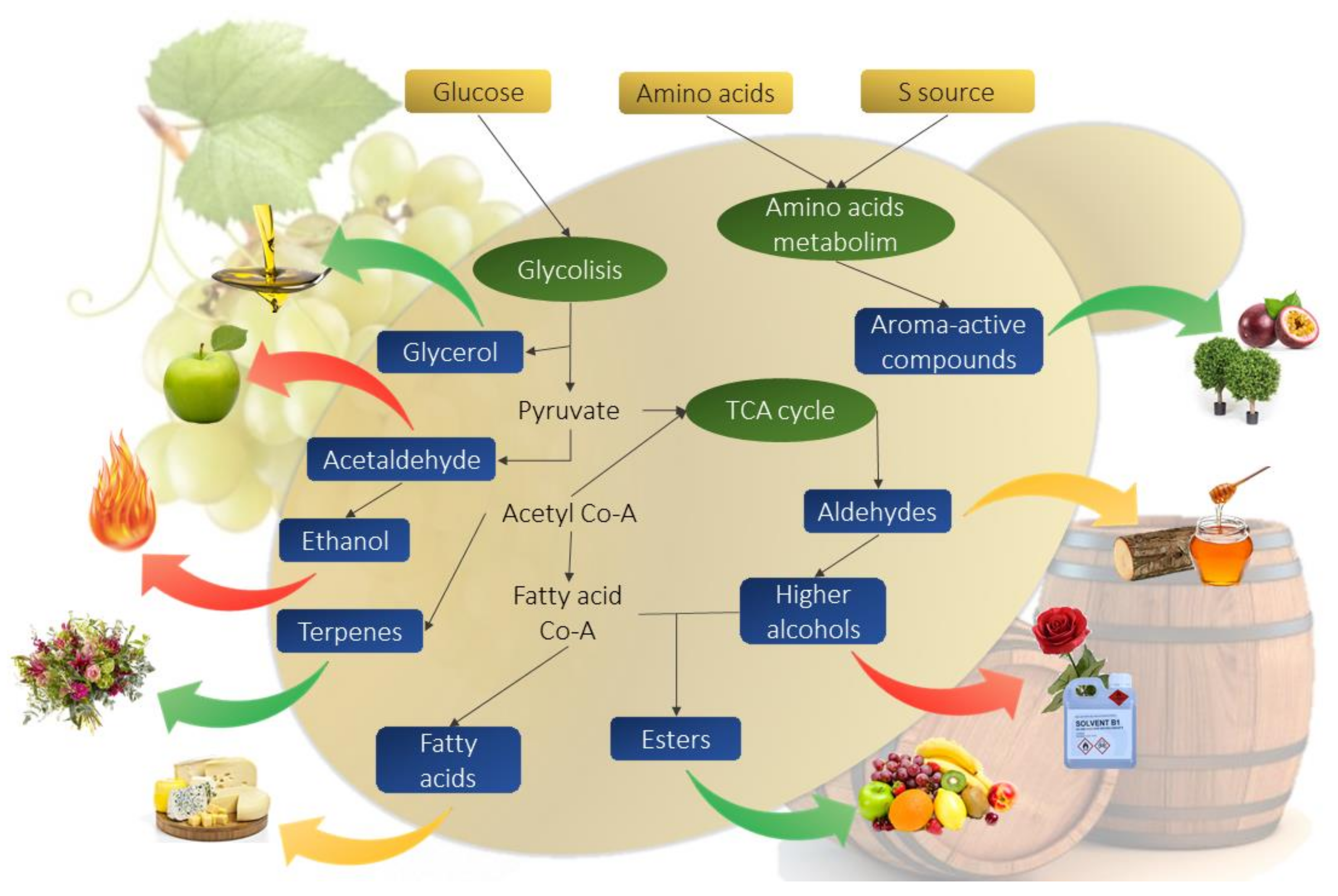
2.4. Aroma Compounds
2.5. Amino Acids
3. Sensory Impact in Regular Wine Fermentation
4. Killer Factor and Its Influence on Wine Ecology
5. Influence of Other Pichia Species on Winemaking
5.1. Pichia Guilliermondii
5.2. Pichia Kudriavzevii
5.3. Pichia fermentans
5.4. Pichia anomala
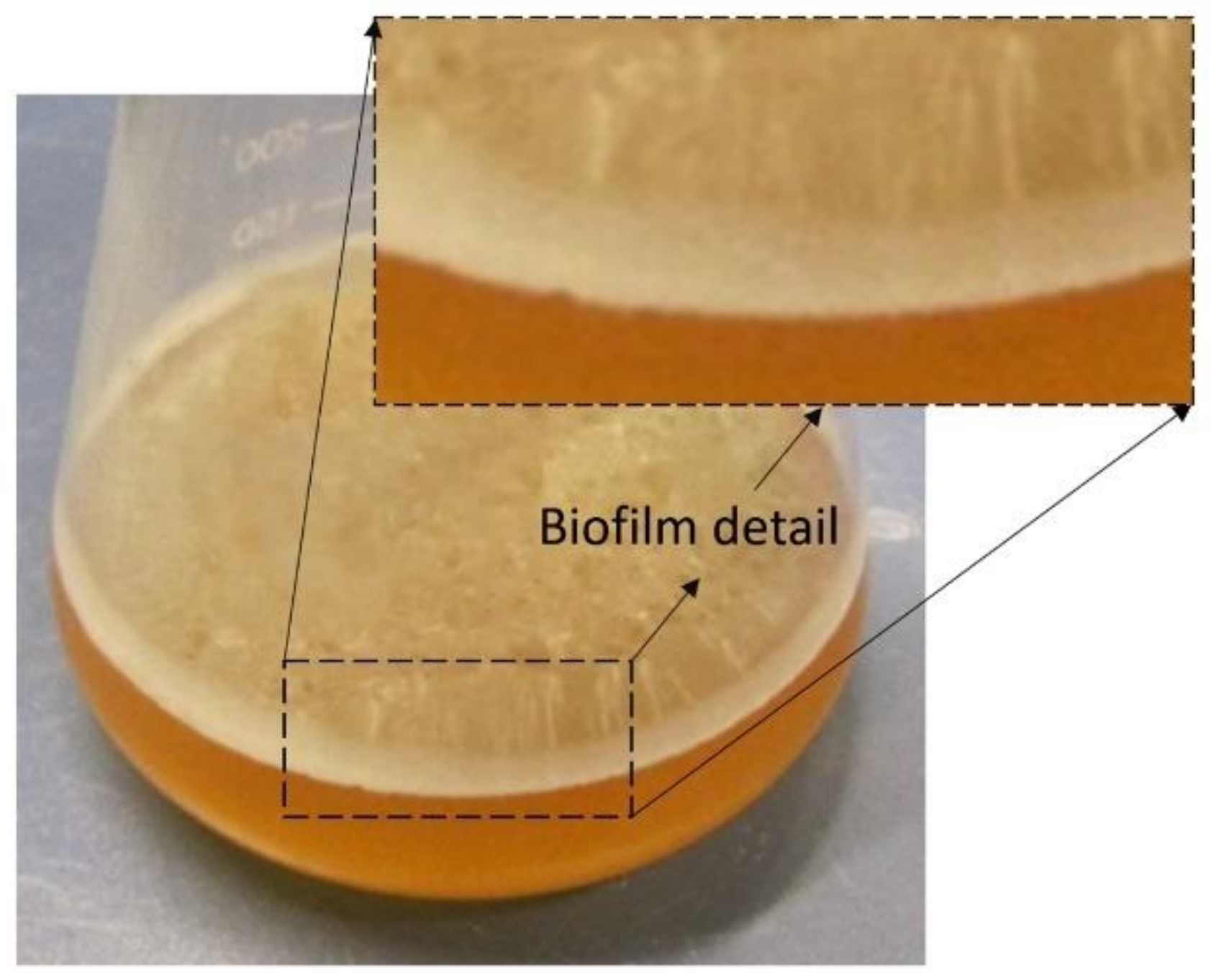
5.5. Pichia membranifaciens

{kind=link}
{kind=link}
{kind=link}
| Teleomorph | Synonym | Impact on Wine Fermentation |
|---|---|---|
| Pichia kluyveri | Hansenula kluyveri | Increased levels of varietal thiols (3-MHA) [2] |
| Pichia fermentans | Candida lambica | Increased levels of higher alcohols, glycerol, and polysaccharides [2] |
| Pichia membranifaciens | Candida valida | Increased levels of esters [4] |
| Pichia terricola | Issatchenkia terricola | Increased β-glucosidase activity [63]; malic acid degradation [64] |
| Pichia kudriavzevii | Candida krusei. Issatchenkia orientalis | Malic acid degradation [17]; increased 2-pheyl-ethanol production [65] |
| Pichia manshurica | - | Spoilage yeast; increased volatile phenols and other off-odors [66] |
| Pichia guilliermondi | Candida guilliermondi, Meyerozyma guilliermondi | Increased color stability [29] |
| Pichia anomala | Candida anomala, Hansenula anomala, Wickerhanomyces anomalus | Increased levels of higher alcohols, acetates, and ethyl esters [2] |
| Pichia pastoris | Komagataella pastoris, K. pseudopastoris, K. phafii | Inhibition of protein haze formation in white wines (genetically modified strain) [67] |
6. Proposed Selection Parameters for P. kluyveri
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascues, E.; Marquina, D.; Santos, A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans Population in Monoculture and Mixed Fermentations: Impact on Wine Characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Int. Food Res. J. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The Genus Metschnikowia in Enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms 2020, 8, 655. [Google Scholar] [CrossRef] [PubMed]
- Puig-Pujol, A.; Ferrando, N.; Capdevila, F.; Ocete, R.; Revilla, E. Yeast biodiversity from Vitis vinifera L., subsp. sylvestris (Gmelin) Hegi to face up the oenological consequences of climate change. BIO Web Conf. EDP Sci. 2016, 7, 02026. [Google Scholar] [CrossRef] [Green Version]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces commercial starter cultures: Scientific trends, recent patents and innovation in the wine sector. Recent Pat. Food Nutr. Agric. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Ciani, M.; Canonico, L.; Oro, L.; Comitini, F. Footprint of nonconventional yeasts and their contribution in alcoholic fermentations. In Biotechnological Progress and Beverage Consumption; Elsevier: Amsterdam, The Netherlands, 2020; pp. 435–465. [Google Scholar]
- Pretorius, I.S. Conducting wine symphonics with the aid of yeast genomics. Beverages 2016, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Amaya-Delgado, L.; Herrera-López, E.; Arrizon, J.; Arellano-Plaza, M.; Gschaedler, A. Performance evaluation of Pichia kluyveri, Kluyveromyces marxianus and Saccharomyces cerevisiae in industrial tequila fermentation. World J. Microb. Biotechnol. 2013, 29, 875–881. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of Sequential Inoculation with Non-Saccharomyces and Saccharomyces Yeasts on Riesling Wine Chemical Composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Varela, J.; Varela, C. Microbiological strategies to produce beer and wine with reduced ethanol concentration. Curr. Opin. Biotechnol. 2019, 56, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Röcker, J.; Strub, S.; Ebert, K.; Grossmann, M. Usage of different aerobic non-Saccharomyces yeasts and experimental conditions as a tool for reducing the potential ethanol content in wines. Eur. Food Res. Technol. 2016, 242, 2051–2070. [Google Scholar] [CrossRef]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Whitener, M.E.B.; Stanstrup, J.; Panzeri, V.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Untangling the wine metabolome by combining untargeted SPME–GCxGC-TOF-MS and sensory analysis to profile Sauvignon blanc co-fermented with seven different yeasts. Metabolomics 2016, 12, 53. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Voon, M.K.W.; Chua, J.-Y.; Huang, D.; Lee, P.-R.; Liu, S.-Q. The effects of co-and sequential inoculation of Torulaspora delbrueckii and Pichia kluyveri on chemical compositions of durian wine. Appl. Microbiol. Biotechnol. 2017, 101, 7853–7863. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased β-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- Whitener, M.B.; Stanstrup, J.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Effect of non-Saccharomyces yeasts on the volatile chemical profile of Shiraz wine. Aust. J. Grape Wine Res. 2017, 23, 179–192. [Google Scholar] [CrossRef]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascues, E.; Calderon, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on varietal aromas during wine making: A review of the impact of varietal aromas on the flavor of wine. Appl. Microbiol. Biotechnol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, A.M.; Guadalupe, V.; Varela, C.; Swiegers, J.H.; Pretorius, I.S.; Agosin, E. Differential synthesis of fermentative aroma compounds of two related commercial wine yeast strains. Food Chem. 2009, 117, 189–195. [Google Scholar] [CrossRef]
- Landete, J.; Ferrer, S.; Pardo, I. Biogenic amine production by lactic acid bacteria, acetic bacteria and yeast isolated from wine. Food Control 2007, 18, 1569–1574. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romano, P. Biotechnological Approach Based on Selected Saccharomyces cerevisiae Starters for Reducing the Use of Sulfur Dioxide in Wine. Microorganisms 2020, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The biology of Pichia membranifaciens killer toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middelbeek, E.; Stumm, C.; Vogels, G. Effects of Pichia kluyveri killer toxin on sensitive cells. Antonie Van Leeuwenhoek 1980, 46, 205–220. [Google Scholar] [CrossRef]
- Labbani, F.-Z.K.; Turchetti, B.; Bennamoun, L.; Dakhmouche, S.; Roberti, R.; Corazzi, L.; Meraihi, Z.; Buzzini, P. A novel killer protein from Pichia kluyveri isolated from an Algerian soil: Purification and characterization of its in vitro activity against food and beverage spoilage yeasts. Antonie van Leeuwenhoek 2015, 107, 961–970. [Google Scholar] [CrossRef]
- Pintar, J.; Starmer, W.T. The costs and benefits of killer toxin production by the yeast Pichia kluyveri. Antonie van Leeuwenhoek 2003, 83, 89–97. [Google Scholar] [CrossRef]
- Sangorrín, M.P.; Lopes, C.A.; Jofré, V.; Querol, A.; Caballero, A.C. Spoilage yeasts from Patagonian cellars: Characterization and potential biocontrol based on killer interactions. World J. Microbiol. Biotechnol. 2008, 24, 945–953. [Google Scholar] [CrossRef]
- Alonso, A.; Belda, I.; Santos, A.; Navascués, E.; Marquina, D. Advances in the control of the spoilage caused by Zygosaccharomyces species on sweet wines and concentrated grape musts. Food Control 2015, 51, 129–134. [Google Scholar] [CrossRef]
- Benito, S.; Morata, A.; Palomero, F.; González, M.; Suárez-Lepe, J. Formation of vinylphenolic pyranoanthocyanins by Saccharomyces cerevisiae and Pichia guillermondii in red wines produced following different fermentation strategies. Food Chem. 2011, 124, 15–23. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The combined use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the anthocyanin wine composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, S.; Palomero, F.; Morata, A.; Calderon, F.; Suarez-Lepe, J.A. A method for estimating Dekkera/Brettanomyces populations in wines. J. Appl. Microbiol. 2009, 106, 1743–1751. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential Non-Saccharomyces and Saccharomyces cerevisiae Fermentations to Reduce the Alcohol Content in Wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Del Monaco, S.M.; Barda, N.B.; Rubio, N.C.; Caballero, A.C. Selection and characterization of a Patagonian Pichia kudriavzevii for wine deacidification. J. Appl. Microbiol. 2014, 117, 451–464. [Google Scholar] [CrossRef]
- Del Mónaco, S.M.; Rodriguez, M.E.; Lopes, C.A. Pichia kudriavzevii as a representative yeast of North Patagonian winemaking terroir. Int. J. Food Microbiol. 2016, 230, 31–39. [Google Scholar] [CrossRef]
- Luan, Y.; Zhang, B.-Q.; Duan, C.-Q.; Yan, G.-L. Effects of different pre-fermentation cold maceration time on aroma compounds of Saccharomyces cerevisiae co-fermentation with Hanseniaspora opuntiae or Pichia kudriavzevii. LWT 2018, 92, 177–186. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.-L.; Li, A.-H.; Su, J.; Wang, X.-C.; Chen, C.-Q.; Tao, Y.-S. Flavor modification of dry red wine from Chinese spine grape by mixed fermentation with Pichia fermentans and S. cerevisiae. LWT 2019, 109, 83–92. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Heras-Vázquez, F.L.; Rodríguez-Vico, F. Influence of sequential yeast mixtures on wine fermentation. Int. J. Food Microbiol. 2005, 98, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Acuña-Fontecilla, A.; Catrileo, D. Formation of aromatic and flavor compounds in wine: A perspective of positive and negative contributions of non-Saccharomyces yeasts. In Winemaking-Stabilization, Aging Chemistry and Biochemistry; IntechOpen: London, UK, 2020. [Google Scholar]
- Romani, C.; Lencioni, L.; Bartolini, A.B.; Ciani, M.; Mannazzu, I.; Domizio, P. Pilot Scale Fermentations of Sangiovese: An Overview on the Impact of Saccharomyces and Non-Saccharomyces Wine Yeasts. Fermentation 2020, 6, 63. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Cañas, P.M.I.; García-Romero, E.; Manso, J.M.H.; Fernández-González, M. Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- Laitila, A.; Sarlin, T.; Raulio, M.; Wilhelmson, A.; Kotaviita, E.; Huttunen, T.; Juvonen, R. Yeasts in malting, with special emphasis on Wickerhamomyces anomalus (synonym Pichia anomala). Antonie van Leeuwenhoek 2011, 99, 75–84. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Santos, A.; Marquina, D. Killer toxin of Pichia membranifaciens and its possible use as a biocontrol agent against grey mould disease of grapevine. Microbiology 2004, 150, 2527–2534. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.; Mauro, M.S.; Bravo, E.; Marquina, D. PMKT2, a new killer toxin from Pichia membranifaciens, and its promising biotechnological properties for control of the spoilage yeast Brettanomyces bruxellensis. Microbiology 2009, 155, 624–634. [Google Scholar] [CrossRef] [Green Version]
- González-Pombo, P.; Fariña, L.; Carrau, F.; Batista-Viera, F.; Brena, B.M. A novel extracellular β-glucosidase from Issatchenkia terricola: Isolation, immobilization and application for aroma enhancement of white Muscat wine. Process Biochem. 2011, 46, 385–389. [Google Scholar] [CrossRef]
- Wen, L.; Wang, L.; Wang, G. Degradation of L-malic and citric acids by Issatchenkia terricola. J. Food Sci. 2011, 32, 220–223. [Google Scholar]
- Martínez-Avila, O.; Sánchez, A.; Font, X.; Barrena, R. 2-Phenylethanol (rose aroma) production potential of an isolated Pichia kudriavzevii through solid-state fermentation. Process Biochem. 2020, 93, 94–103. [Google Scholar] [CrossRef]
- Perpetuini, G.; Tittarelli, F.; Battistelli, N.; Suzzi, G.; Tofalo, R. Contribution of Pichia manshurica strains to aroma profile of organic wines. Eur. Food Res. Technol. 2020, 246, 1405–1417. [Google Scholar] [CrossRef]
- Schmidt, S.A.; Tan, E.L.; Brown, S.; Nasution, U.J.; Pettolino, F.; Macintyre, O.J.; Lopes, M.D.B.; Waters, E.J.; Anderson, P.A. Hpf2 glycan structure is critical for protection against protein haze formation in white wine. J. Agric. Food Chem. 2009, 57, 3308–3315. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascues, E.; Marquina, D.; Calderon, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guijo, S.; Mauricio, J.; Salmon, J.-M.; Ortega, J. Determination of the relative ploidy in different Saccharomyces cerevisiae strains used for fermentation and “flor” film ageing of dry sherry-type wines. Yeast 1997, 13, 101–117. [Google Scholar] [CrossRef]
- Ruiz-Muñoz, M.; Cordero-Bueso, G.; Benítez-Trujillo, F.; Martínez, S.; Pérez, F.; Cantoral, J.M. Rethinking about flor yeast diversity and its dynamic in the “criaderas and soleras” biological aging system. Food Microbiol. 2020, 92, 103553. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Ruiz-Muñoz, M.; González-Moreno, M.; Chirino, S.; Bernal-Grande, M.D.C.; Cantoral, J.M. The Microbial Diversity of Sherry Wines. Fermentation 2018, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Peinado, R.A.; Mauricio, J.C. Biologically aged wines. In Wine Chemistry and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2009; pp. 81–101. [Google Scholar]
- Benito, A.; Calderon, F.; Benito, S. Combined Use of S. pombe and L. thermotolerans in Winemaking. Beneficial Effects Determined Through the Study of Wines’ Analytical Characteristics. Molecules 2016, 21, 1744. [Google Scholar] [CrossRef] [Green Version]



| Species | Oenological Impact |
|---|---|
| Torulospora delbrueckii | Increased esters and thiols; acetic acid consumption [7,8] |
| Lachancea thermotolerans | Increased L-lactic acid, glycerol, and 2-phenyl-ethanol [9,10,11] |
| Metschnikowia pulcherrima | Increased esters, terpenes, thiols, and aromatic complexity [12] |
| Schizosaccharomyces pombe | Increased deacidification by L-malic acid degradation [13] |
| Candida zemplinina | Increased glycerol and succinic acid; decreased acetic acid and higher alcohols [2] |
| Hanseniaspora spp. | Increased acetate esters and terpenes; biogenic amine adsorption [2] |
| Hansenula anomala | Low C6 alcohols; increased higher alcohols and acetate and ethyl esters [2] |
| Zygosaccharomyces bailii | Increased polysaccharides; acetic acid [2] |
| Pichia guillermondii | Increased color stability and 4-ethyl-phenol production [2] |
| Pichia kluyveri | Increased varietal thiols and esters [2] |
| Group | Aroma Compound | Structure | Odor Descriptor | Perception Threshold (ng/L) | Reference |
|---|---|---|---|---|---|
| Higher alcohols | 2-methyl butanol |  | Harsh, nail polish remover | 30,000 | [36] |
| 3-methyl butanol |  | Harsh, nail polish remover | 30,000 | [36] | |
| 2-phenyl ethanol |  | Rose | 10,000 | [36] | |
| Esters | 2-phenyl-ethyl acetate |  | Rose, raspberry | 250 | [36] |
| 2-methyl-butyl acetate |  | Banana | 5 | [36] | |
| Terpenes | Linalool |  | Flowery, fruity | 6 for white varieties 15 for red varieties | [34] |
| Hotrienol |  | Faintly flowery, elderflower | 110 | [34] | |
| Thiols | 3-SHA |  | Passionfruit, box tree | 4 | [34] |
| 3-SH |  | Grapefruit, citrus peel | 60 | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente, J.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology. Int. J. Mol. Sci. 2021, 22, 1196. https://doi.org/10.3390/ijms22031196
Vicente J, Calderón F, Santos A, Marquina D, Benito S. High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology. International Journal of Molecular Sciences. 2021; 22(3):1196. https://doi.org/10.3390/ijms22031196
Chicago/Turabian StyleVicente, Javier, Fernando Calderón, Antonio Santos, Domingo Marquina, and Santiago Benito. 2021. "High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology" International Journal of Molecular Sciences 22, no. 3: 1196. https://doi.org/10.3390/ijms22031196