
Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
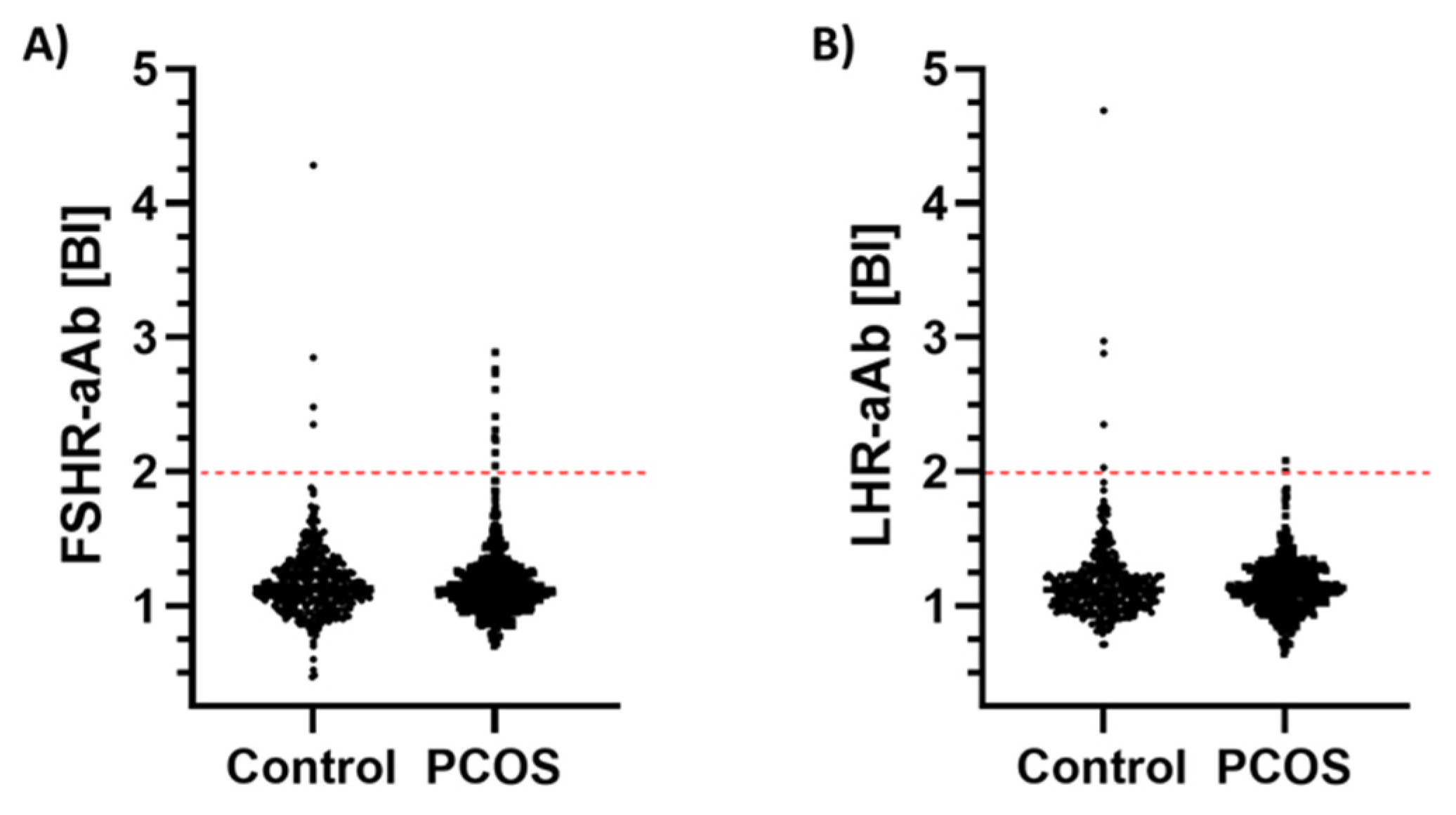
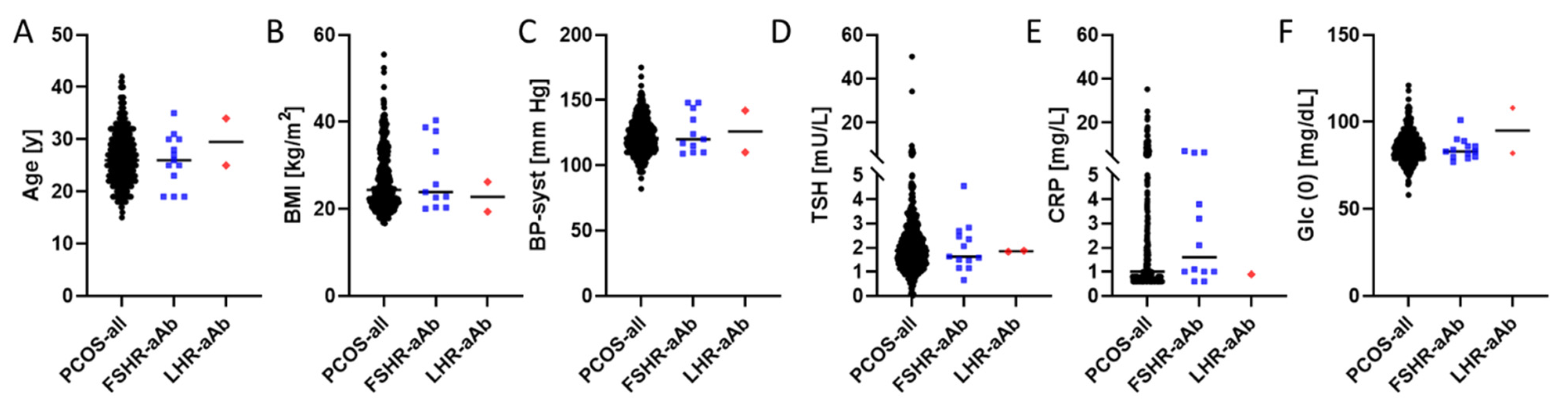
2. Results
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.2. Construction and Expression of GPCR-Luciferase Fusion Proteins
4.3. Quantitative Analysis of FSH-R-aAb and LH-R-aAb in Human Serum Samples
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deswal, R.; Narwal, V.; Dang, A.; Pundir, C.S. The prevalence of polycystic ovary syndrome: A brief systematic review. J. Hum. Reprod. Sci. 2020, 13, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Witchel, S.F.; Teede, H.J.; Pena, A.S. Curtailing PCOS. Pediatr. Res. 2020, 87, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The androgen excess and PCOS society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef]
- Sirmans, S.M.; Pate, K.A. Epidemiology, diagnosis, and management of polycystic ovary syndrome. Clin. Epidemiol. 2013, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rosenfield, R.L.; Ehrmann, D.A. The pathogenesis of polycystic ovary syndrome (PCOS): The hypothesis of PCOS as functional ovarian hyperandrogenism revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef] [PubMed]
- Day, F.; Karaderi, T.; Jones, M.R.; Meun, C.; He, C.; Drong, A.; Kraft, P.; Lin, N.; Huang, H.; Broer, L.; et al. Large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for different diagnosis criteria. PLoS Genet. 2018, 14, e1007813. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Wong, H.; Ahluwalia, N.; Go, R.M.; Guerrero-Go, M.A. Metabolic, hormonal, immunologic, and genetic factors associated with the incidence of thyroid disorders in polycystic ovarian syndrome patients. Cureus 2020, 12, e11681. [Google Scholar] [CrossRef]
- Giampaolino, P.; Foreste, V.; Di Filippo, C.; Gallo, A.; Mercorio, A.; Serafino, P.; Improda, F.P.; Verrazzo, P.; Zara, G.; Buonfantino, C.; et al. Microbiome and PCOS: State-of-art and future aspects. Int. J. Mol. Sci. 2021, 22, 2048. [Google Scholar] [CrossRef]
- Duan, L.Y.; An, X.D.; Zhang, Y.H.; Jin, D.; Zhao, S.H.; Zhou, R.R.; Duan, Y.Y.; Zhang, Y.Q.; Liu, X.M.; Lian, F.M. Gut microbiota as the critical correlation of polycystic ovary syndrome and type 2 diabetes mellitus. Biomed. Pharmacother. 2021, 142, 12094. [Google Scholar] [CrossRef]
- Trummer, C.; Pilz, S.; Schwetz, V.; Obermayer-Pietsch, B.; Lerchbaum, E. Vitamin D, PCOS and androgens in men: A systematic review. Endocr. Connect. 2018, 7, R95–R113. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Y.; Ye, J.; Wei, H.; Huang, Z.; Ning, X.; Fu, X. A comparative study on insulin secretion, insulin resistance and thyroid function in patients with polycystic ovary syndrome with and without Hashimoto’s thyroiditis. Diabetes Metab. Syndr. Obes. 2021, 14, 1817–1821. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Chen, C.C.; Hsieh, M.C.; Ho, C.W.; Hsu, S.P.; Yip, H.T.; Kao, C.H. Graves’ disease could increase polycystic ovary syndrome and comorbidities in Taiwan. Curr. Med. Res. Opin. 2020, 36, 1063–1067. [Google Scholar] [CrossRef]
- Kluzek, S.; Rubin, K.H.; Sanchez-Santos, M.; O’Hanlon, M.S.; Andersen, M.; Glintborg, D.; Abrahamsen, B. Accelerated osteoarthritis in women with polycystic ovary syndrome: A prospective nationwide registry-based cohort study. Arthritis Res. Ther. 2021, 23, 225. [Google Scholar] [CrossRef]
- Busiah, K.; Colmenares, A.; Bidet, M.; Tubiana-Rufi, N.; Levy-Marchal, C.; Delcroix, C.; Jacquin, P.; Martin, D.; Benadjaoud, L.; Jacqz-Aigrain, E.; et al. High prevalence of polycystic ovary syndrome in type 1 diabetes mellitus adolescents: Is there a difference depending on the NIH and rotterdam criteria? Horm. Res. Paediat. 2017, 87, 333–341. [Google Scholar] [CrossRef]
- Ragab, A.; Barakat, R.; Ragheb, M.; State, O.; Badawy, A. Subfertility treatment in women with systemic lupus erythematosus. J. Obstet. Gynaecol. 2012, 32, 569–571. [Google Scholar] [CrossRef]
- De Simone, C.; Caldarola, G.; Corbeddu, M.; Moro, F.; Tropea, A.; Moretta, G.; Apa, R. A possible role of polycystic ovary syndrome for pregnancy complications in women with psoriasis. Drug Dev. Res. 2014, 75, S64–S66. [Google Scholar] [CrossRef] [PubMed]
- Arduc, A.; Dogan, B.A.; Bilmez, S.; Nasiroglu, N.I.; Tuna, M.M.; Isik, S.; Berker, D.; Guler, S. High prevalence of Hashimoto’s thyroiditis in patients with polycystic ovary syndrome: Does the imbalance between estradiol and progesterone play a role? Endocr. Res. 2015, 40, 204–210. [Google Scholar] [CrossRef]
- Janssen, O.E.; Mehlmauer, N.; Hahn, S.; Offner, A.H.; Gartner, R. High prevalence of autoimmune thyroiditis in patients with polycystic ovary syndrome. Eur. J. Endocrinol. 2004, 150, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Thangaratinam, S.; Tan, A.; Knox, E.; Kilby, M.D.; Franklyn, J.; Coomarasamy, A. Association between thyroid autoantibodies and miscarriage and preterm birth: Meta-analysis of evidence. BMJ 2011, 342, d2616. [Google Scholar] [CrossRef] [Green Version]
- Dhillon-Smith, R.K.; Tobias, A.; Smith, P.P.; Middleton, L.J.; Sunner, K.K.; Baker, K.; Farrell-Carver, S.; Bender-Atik, R.; Agrawal, R.; Bhatia, K.; et al. The prevalence of thyroid dysfunction and autoimmunity in women with history of miscarriage or subfertility. J. Clin. Endocrinol. Metab. 2020, 105, 2667–2677. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Polycystic ovary syndrome (PCOS), an inflammatory, systemic, lifestyle endocrinopathy. J. Steroid Biochem. Mol. Biol 2018, 182, 27–36. [Google Scholar] [CrossRef]
- Greenbaum, H.; Galper, B.L.; Decter, D.H.; Eisenberg, V.H. Endometriosis and autoimmunity: Can autoantibodies be used as a non-invasive early diagnostic tool? Autoimmun. Rev. 2021, 20, 102795. [Google Scholar] [CrossRef]
- Seppa, S.; Kuiri-Hanninen, T.; Holopainen, E.; Voutilainen, R. Management of endocrine disease: Diagnosis and management of primary amenorrhea and female delayed puberty. Eur. J. Endocrinol. 2021, 184, R225–R242. [Google Scholar] [CrossRef]
- Liao, B.; Qiao, J.; Pang, Y. Central regulation of PCOS: Abnormal neuronal-reproductive-metabolic circuits in PCOS pathophysiology. Front. Endocrinol. 2021, 12, 667422. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Ren, Y.A.; Candelaria, N.; Adams, J.E.; Rajkovic, A. Ovarian follicular theca cell recruitment, differentiation, and impact on fertility: 2017 update. Endocr. Rev. 2018, 39, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Recchia, K.; Jorge, A.S.; Pessoa, L.V.F.; Botigelli, R.C.; Zugaib, V.C.; de Souza, A.F.; Martins, D.D.S.; Ambrosio, C.E.; Bressan, F.F.; Pieri, N.C.G. Actions and roles of FSH in germinative cells. Int. J. Mol. Sci. 2021, 22, 10110. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, S.M.; Rapoport, B. Thyrotropin-blocking autoantibodies and thyroid-stimulating autoantibodies: Potential mechanisms involved in the pendulum swinging from hypothyroidism to hyperthyroidism or vice versa. Thyroid 2013, 23, 14–24. [Google Scholar] [CrossRef]
- Querat, B.; Sellouk, A.; Salmon, C. Phylogenetic analysis of the vertebrate glycoprotein hormone family including new sequences of sturgeon (Acipenser baeri) beta subunits of the two gonadotropins and the thyroid-stimulating hormone. Biol. Reprod. 2000, 63, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Vassart, G.; Pardo, L.; Costagliola, S. A molecular dissection of the glycoprotein hormone receptors. Trends Biochem. Sci. 2004, 29, 119–126. [Google Scholar] [CrossRef]
- Dias, J.A.; Gates, S.A.; Reichert, L.E. Evidence for the presence of follicle-stimulating-hormone receptor antibody in human-serum. Fertil. Steril. 1982, 38, 330–338. [Google Scholar] [CrossRef]
- Tang, V.W.; Faiman, C. Premature ovarian failure—A search for circulating factors against gonadotropin receptors. Am. J. Obstet. Gynecol. 1983, 146, 816–821. [Google Scholar] [CrossRef]
- Chiauzzi, V.A.; Bussmann, L.; Calvo, J.C.; Sundblad, V.; Charreau, E.H. Circulating immunoglobulins that inhibit the binding of follicle-stimulating hormone to its receptor: A putative diagnostic role in resistant ovary syndrome? Clin. Endocrinol. 2004, 61, 46–54. [Google Scholar] [CrossRef]
- Simoni, M.; Paschke, R.; Nieschlag, E. A search for circulating immunoglobulins blocking follicle-stimulating hormone action in male idiopathic infertility. Int. J. Androl. 1993, 16, 129–135. [Google Scholar] [CrossRef]
- Mobeen, H.; Afzal, N.; Kashif, M. Polycystic ovary syndrome may be an autoimmune disorder. Scientifica 2016, 2016, 4071735. [Google Scholar] [CrossRef] [Green Version]
- Beermann, S.; Seifert, R.; Neumann, D. Commercially available antibodies against human and murine histamine H-4-receptor lack specificity. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 125–135. [Google Scholar] [CrossRef]
- Marchalant, Y.; Brownjohn, P.W.; Bonnet, A.; Kleffmann, T.; Ashton, J.C. Validating antibodies to the cannabinoid CB2 Receptor: Antibody sensitivity is not evidence of antibody specificity. J. Histochem. CytoChem. 2014, 62, 395–404. [Google Scholar] [CrossRef]
- Ju, M.S.; Jung, S.T. Antigen design for successful isolation of highly challenging therapeutic anti-GPCR antibodies. Int. J. Mol. Sci. 2020, 21, 8240. [Google Scholar] [CrossRef] [PubMed]
- Bizzaro, N. Autoantibodies as predictors of disease: The clinical and experimental evidence. Autoimmun. Rev. 2007, 6, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Adamus, G. Current techniques to accurately measure anti-retinal autoantibodies. Expert Rev. Ophthalmol 2020, 15, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Carmona, S.J.; Nielsen, M.; Schafer-Nielsen, C.; Mucci, J.; Altcheh, J.; Balouz, V.; Tekiel, V.; Frasch, A.C.; Campetella, O.; Buscaglia, C.A.; et al. Towards high-throughput immunomics for infectious diseases: Use of next-generation peptide microarrays for rapid discovery and mapping of antigenic determinants. Mol. Cell Proteom. 2015, 14, 1871–1884. [Google Scholar] [CrossRef] [Green Version]
- Zambrano-Mila, M.S.; Blacio, K.E.S.; Vispo, N.S. Peptide phage display: Molecular principles and biomedical applications. Ther. Innov. Regul. Sci. 2020, 54, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Futami, J.; Nonomura, H.; Kido, M.; Niidoi, N.; Fujieda, N.; Hosoi, A.; Fujita, K.; Mandai, K.; Atago, Y.; Kinoshita, R.; et al. Sensitive multiplexed quantitative analysis of autoantibodies to cancer antigens with chemically s-cationized full-length and water-soluble denatured proteins. Bioconjug. Chem. 2015, 26, 2076–2084. [Google Scholar] [CrossRef]
- Deutschmann, C.; Roggenbuck, D.; Schierack, P.; Rodiger, S. Autoantibody testing by enzyme-linked immunosorbent assay-a case in which the solid phase decides on success and failure. Heliyon 2020, 6, e03270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burbelo, P.D.; Beck, L.H.; Waldman, M. Detection and monitoring PLA(2)R autoantibodies by LIPS in membranous nephropathy. J. Immunol. Methods 2017, 444, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minich, W.B.; Loos, U. Detection of functionally different types of pathological autoantibodies against thyrotropin receptor in Graves’ patients sera by luminescent immunoprecipitation analysis. Exp. Clin. Endocr. Diab 2000, 108, 110–119. [Google Scholar] [CrossRef]
- Eleftheriadou, A.M.; Mehl, S.; Renko, K.; Kasim, R.H.; Schaefer, J.A.; Minich, W.B.; Schomburg, L. Re-visiting autoimmunity to sodium-iodide symporter and pendrin in thyroid disease. Eur. J. Endocrinol. 2020, 183, 571–580. [Google Scholar] [CrossRef]
- Ajjan, R.A.; Kemp, E.H.; Waterman, E.A.; Watson, P.F.; Endo, T.; Onaya, T.; Weetman, A.P. Detection of binding and blocking autoantibodies to the human sodium-iodide symporter in patients with autoimmune thyroid disease. J. Clin. Endocrinol. Metab. 2000, 85, 2020–2027. [Google Scholar] [CrossRef] [Green Version]
- Seissler, J.; Wagner, S.; Schott, M.; Lettmann, M.; Feldkamp, J.; Scherbaum, W.A.; Morgenthaler, M.G. Low frequency of autoantibodies to the human Na+/I− symporter in patients with autoimmune thyroid disease. J. Clin. Endocr. Metab. 2000, 85, 4630–4634. [Google Scholar] [CrossRef]
- Boege, F.; Westenfeld, R.; Jahns, R. β1AAb determined by peptide ELISA: A signal in the noise? J. Am. Coll. Cardiol. 2017, 70, 807–808. [Google Scholar] [CrossRef]
- Bornholz, B.; Roggenbuck, D.; Jahns, R.; Boege, F. Diagnostic and therapeutic aspects of beta1-adrenergic receptor autoantibodies in human heart disease. Autoimmun. Rev. 2014, 13, 954–962. [Google Scholar] [CrossRef] [Green Version]
- Haberland, A.; Muller, J.; Wallukat, G.; Wenzel, K. Antigen-free control wells in an ELISA set-up for the determination of autoantibodies against G protein-coupled receptors-a requisite for correct data evaluation. Anal. Bioanal. Chem. 2018, 410, 5101–5105. [Google Scholar] [CrossRef] [Green Version]
- Kem, D.C.; Li, H.L.; Yu, X.C.; Weedin, E.; Reynolds, A.C.; Forsythe, E.; Beel, M.; Fischer, H.; Hines, B.; Guo, Y.K.; et al. The role of GnRH receptor autoantibodies in polycystic ovary syndrome. J. Endocr. Soc. 2020, 4, bvaa078. [Google Scholar] [CrossRef]
- Sattler, L.M.; Schniewind, H.A.; Minich, W.B.; Haudum, C.W.; Niklowitz, P.; Munzker, J.; Kovacs, G.L.; Reinehr, T.; Obermayer-Pietsch, B.; Schomburg, L. Natural autoantibodies to the gonadotropin-releasing hormone receptor in polycystic ovarian syndrome. PLoS ONE 2021, 16, e0249639. [Google Scholar] [CrossRef] [PubMed]
- Mora, M.; Angelini, C.; Bignami, F.; Bodin, A.M.; Crimi, M.; Di Donato, J.H.; Felice, A.; Jaeger, C.; Karcagi, V.; LeCam, Y.; et al. The EuroBioBank Network: 10 years of hands-on experience of collaborative, transnational biobanking for rare diseases. Eur. J. Hum. Genet. 2015, 23, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Sargsyan, K.; Macheiner, T.; Story, P.; Strahlhofer-Augsten, M.; Plattner, K.; Riegler, S.; Granitz, G.; Bayer, M.; Huppertz, B. Sustainability in biobanking: Model of biobank graz. Biopreserv. Biobank. 2015, 13, 410–420. [Google Scholar] [CrossRef]
- Demircan, K.; Bengtsson, Y.; Sun, Q.; Brange, A.; Vallon-Christersson, J.; Rijntjes, E.; Malmberg, M.; Saal, L.H.; Ryden, L.; Borg, A.; et al. Serum selenium, selenoprotein P and glutathione peroxidase 3 as predictors of mortality and recurrence following breast cancer diagnosis: A multicentre cohort study. Redox. Biol. 2021, 47, 102145. [Google Scholar] [CrossRef]
- Sun, Q.; Mehl, S.; Renko, K.; Seemann, P.; Gorlich, C.L.; Hackler, J.; Minich, W.B.; Kahaly, G.J.; Schomburg, L. Natural autoimmunity to selenoprotein P impairs selenium transport in Hashimoto’s thyroiditis. Int. J. Mol. Sci. 2021, 22, 13088. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Helm, M.; Teede, H.; Dunaif, A.; Dokras, A. Delayed diagnosis and a lack of information associated with dissatisfaction in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2017, 102, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Franks, S. Controversy in clinical endocrinology: Diagnosis of polycystic ovarian syndrome: In defense of the Rotterdam criteria. J. Clin. Endocrinol. Metab. 2006, 91, 786–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzan, V.; Lerchbaum, E.; Missbrenner, C.; Heijboer, A.C.; Goschnik, M.; Trummer, C.; Theiler-Schwetz, V.; Haudum, C.; Gumpold, R.; Schweighofer, N.; et al. Risk of insulin resistance and metabolic syndrome in women with hyperandrogenemia: A comparison between PCOS phenotypes and beyond. J. Clin. Med. 2021, 10, 829. [Google Scholar] [CrossRef]
- Munzker, J.; Lindheim, L.; Adaway, J.; Trummer, C.; Lerchbaum, E.; Pieber, T.R.; Keevil, B.; Obermayer-Pietsch, B. High salivary testosterone-to-androstenedione ratio and adverse metabolic phenotypes in women with polycystic ovary syndrome. Clin. Endocrinol. 2017, 86, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Munzker, J.; Hofer, D.; Trummer, C.; Ulbing, M.; Harger, A.; Pieber, T.; Owen, L.; Keevil, B.; Brabant, G.; Lerchbaum, E.; et al. Testosterone to dihydrotestosterone ratio as a new biomarker for an adverse metabolic phenotype in the polycystic ovary syndrome. J. Clin. Endocr. Metab. 2015, 100, 653–660. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schniewind, H.A.; Sattler, L.-M.; Haudum, C.W.; Münzker, J.; Minich, W.B.; Obermayer-Pietsch, B.; Schomburg, L. Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome. Int. J. Mol. Sci. 2021, 22, 13667. https://doi.org/10.3390/ijms222413667
Schniewind HA, Sattler L-M, Haudum CW, Münzker J, Minich WB, Obermayer-Pietsch B, Schomburg L. Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome. International Journal of Molecular Sciences. 2021; 22(24):13667. https://doi.org/10.3390/ijms222413667
Chicago/Turabian StyleSchniewind, Hanna A., Lisa-Marie Sattler, Christoph W. Haudum, Julia Münzker, Waldemar B. Minich, Barbara Obermayer-Pietsch, and Lutz Schomburg. 2021. "Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome" International Journal of Molecular Sciences 22, no. 24: 13667. https://doi.org/10.3390/ijms222413667