
Regulatory Peptides in Asthma


Abstract
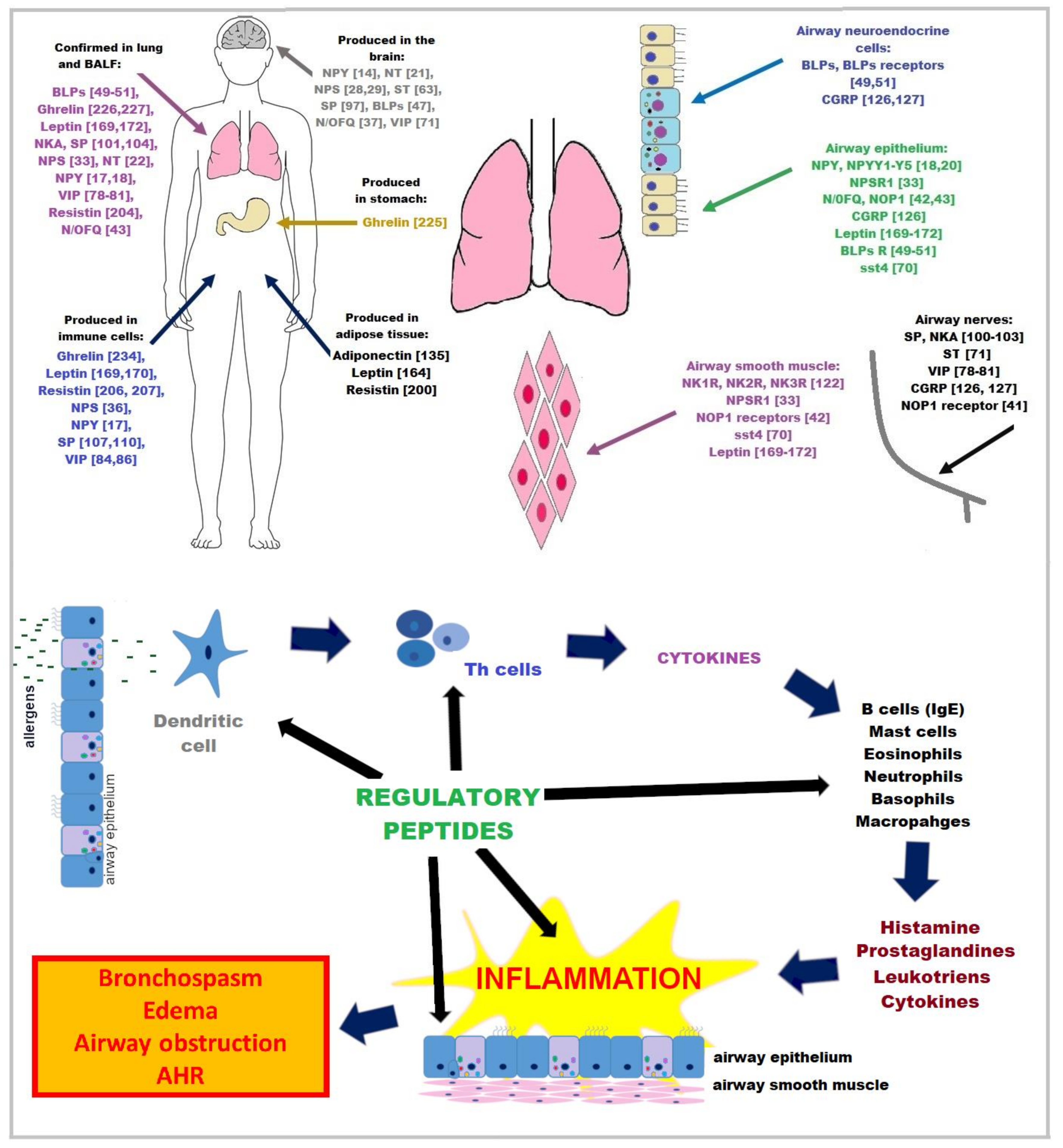
:1. Introduction
2. Neuropeptides
2.1. Neuropeptide Y (NPY)
2.2. Neurotensin (NT)
2.3. Neuropeptide S (NPS)
2.4. Nociceptin (N)/Orphanin (OFQ)
2.5. Bombesin (BN)
2.6. Somatostatin (ST)
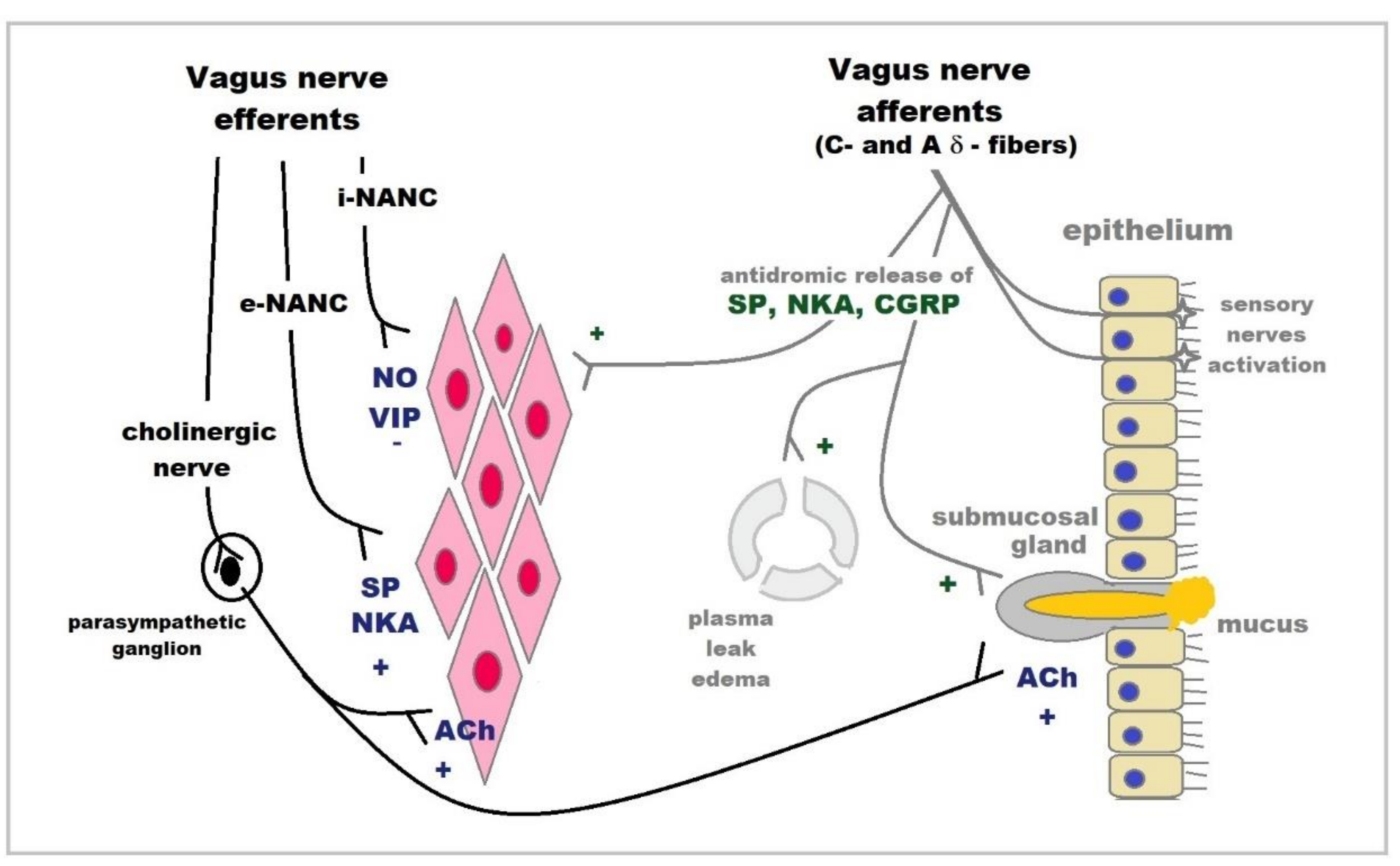
2.7. Vasoactive Intestinal Polypeptide (VIP)
2.8. Tachykinins
2.9. CGRP
3. Adipokines
3.1. Adiponectin
3.2. Leptin
3.3. Resistin and Resistin-like Molecule Family (RELM) Proteins
4. Stomach Peptide
Ghrelin
5. Antimicrobial Peptides (AMPs)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Enilari, O.; Sinha, S. The global impact of asthma in adult populations. Ann. Glob. Health 2019, 85, 2. [Google Scholar] [CrossRef] [Green Version]
- GINA 2021. Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. 2021. Available online: https://ginasthma.org/gina-reports/ (accessed on 2 June 2021).
- Barnes, P.J. The cytokine network in asthma and chronic obstructive pulmonary disease. J. Clin. Investig. 2008, 118, 3546–3556. [Google Scholar] [CrossRef] [Green Version]
- Kaczyńska, K.; Zając, D.; Wojciechowski, P.; Kogut, E.; Szereda-Przestaszewska, M. Neuropeptides and breathing in health and disease. Pulm. Pharmacol. Ther. 2018, 48, 217–224. [Google Scholar] [CrossRef]
- Pavón-Romero, G.F.; Serrano-Pérez, N.H.; García-Sánchez, L.; Ramírez-Jiménez, F.; Terán, L.M. Neuroimmune Pathophysiology in Asthma. Front. Cell Dev. Biol. 2021, 9, 663535. [Google Scholar] [CrossRef]
- Van Lommel, A. Pulmonary neuroendocrine cells (PNEC) and neuroepithelial bodies (NEB): Chemoreceptors and regulators of lung development. Paediatr. Respir. Rev. 2001, 2, 171–176. [Google Scholar] [CrossRef]
- Underner, M.; Millet, C.; Charrière, V.; Dore, P.; Meurice, J.C.; Patte, F. Neuropeptides and respiratory diseases: Prospects in the treatment of asthma. Rev. Pneumol. Clin. 1989, 45, 144–151. [Google Scholar] [PubMed]
- Barnes, P.J. Neuropeptides as modulators of airway function. Agents Actions Suppl. 1990, 31, 175–196. [Google Scholar]
- Saaresranta, T.; Polo, O. Hormones and breathing. Chest 2002, 122, 2165–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russjan, E.; Kaczyńska, K. Beneficial Effects of Neurotensin in Murine Model of Hapten-Induced Asthma. Int. J. Mol. Sci. 2019, 20, 5025. [Google Scholar] [CrossRef] [Green Version]
- Oda, N.; Miyahara, N.; Taniguchi, A.; Morichika, D.; Senoo, S.; Fujii, U.; Itano, J.; Gion, Y.; Kiura, K.; Kanehiro, A.; et al. Requirement for neuropeptide Y in the development of type 2 responses and allergen-induced airway hyperresponsiveness and inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L407–L417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hersoug, L.G.; Linneberg, A. The link between the epidemics of obesity and allergic diseases: Does obesity induce decreased immune tolerance? Allergy 2007, 62, 1205–1213. [Google Scholar] [CrossRef]
- Garcia, P.; Sood, A. Adiponectin in Pulmonary Disease and Critically Ill Patients. Curr. Med. Chem. 2012, 19, 5493–5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehlert, D.R. Introduction to the reviews on neuropeptide Y. Neuropeptides 2004, 38, 135–140. [Google Scholar] [CrossRef]
- Kuo, L.E.; Kitlinska, J.B.; Tilan, J.U.; Li, L.; Baker, S.B.; Johnson, M.D.; Lee, E.W.; Burnett, M.S.; Fricke, S.T.; Kvetnansky, R.; et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nat. Med. 2007, 13, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Buttari, B.; Profumo, E.; Domenici, G.; Tagliani, A.; Ippoliti, F.; Bonini, S.; Businaro, R.; Elenkov, I.; Riganò, R. Neuropeptide Y induces potent migration of human immature dendritic cells and promotes a Th2 polarization. FASEB J. 2014, 28, 3038–3049. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Rao, P.T.; Wheway, J.; Sierro, F.; Mackay, F.; Herzog, H. Y1 signaling has a critical role in allergic airway inflammation. Immun. Cell Biol. 2011, 89, 882–888. [Google Scholar] [CrossRef]
- Makinde, T.O.; Steininger, R.; Agrawal, D.K. NPY and NPY receptors in airway structural and inflammatory cells in allergic asthma. Exp. Mol. Pathol. 2013, 94, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Cadieux, A.; Benchekroun, M.T.; St-Pierre, S.; Fournier, A. Bronchoconstrictor action of neuropeptide Y (NPY) in isolated guinea pig airways. Neuropeptides 1989, 13, 215–219. [Google Scholar] [CrossRef]
- Li, S.; Koziol-White, C.; Jude, J.; Jiang, M.; Zhao, H.; Cao, G.; Yoo, E.; Jester, W.; Morley, M.P.; Zhou, S.; et al. Epithelium-generated neuropeptide Y induces smooth muscle contraction to promote airway hyperresponsiveness. J. Clin. Investig. 2016, 126, 1978–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, M.R.; Kunos, G. Therapeutic approaches targeting the neurotensin receptors. Expert Opin. Ther. Pat. 2021, 31, 361–386. [Google Scholar] [CrossRef]
- Vacca, P.; Carbone, R.; Monselise, A.; Grosso, M.; Bottino, G. Neurotensin pulmonary metabolism in normal and asthmatic subjects. Eur. Rev. Med. Pharmacol. Sci. 2003, 7, 75–80. [Google Scholar] [PubMed]
- Aas, P.; Helle, K.B. Neurotensin receptors in the rat bronchi. Regul. Pept. 1982, 3, 405–413. [Google Scholar] [CrossRef]
- Djokic, T.D.; Dusser, D.J.; Borson, D.B.; Nadel, J.A. Neutral endopeptidase modulates neurotensin-induced airway contraction. J. Appl. Physiol. 1989, 66, 2338–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.A.; Gully, D.; Naline, E.; Advenier, C. Neurotensin modulates cholinergic and noncholinergic neurotransmission in guinea-pig main bronchi in vitro. Neuropeptides 1994, 26, 159–166. [Google Scholar] [CrossRef]
- Russjan, E.; Andrzejewski, K.; Sulejczak, D.; Kleczkowska, P.; Kaczyńska, K. Endomorphin-2- and Neurotensin- Based Chimeric Peptide Attenuates Airway Inflammation in Mouse Model of Nonallergic Asthma. Int. J. Mol. Sci. 2019, 20, 5935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russjan, E.; Zając, D.; Sulejczak, D.; Kleczkowska, P.; Kaczyńska, K. Contribution of opioid and neurotensin receptors in the anti-inflammatory activity of PK20 hybrid compound in murine airways. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1162–1170. [Google Scholar] [CrossRef]
- Reinscheid, R.K. Neuropeptide S: Anatomy, pharmacology, genetics and physiological functions. Results Probl. Cell Differ. 2008, 46, 145–158. [Google Scholar] [CrossRef]
- Kushikata, T.; Hirota, K.; Saito, J.; Takekawa, D. Roles of Neuropeptide S in Anesthesia, Analgesia, and Sleep. Pharmaceuticals 2021, 14, 483. [Google Scholar] [CrossRef]
- Castro-Giner, F.; de Cid, R.; Gonzalez, J.R.; Jarvis, D.; Heinrich, J.; Janson, C.; Omenaas, E.R.; Matheson, M.C.; Pin, I.; Anto, J.M.; et al. Positionally cloned genes and age-specific effects in asthma and atopy: An international population-based cohort study (ECRHS). Thorax 2010, 65, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, N.; Ezer, S.; Kebede Merid, S.; Gaertner, V.D.; Söderhäll, C.; D’Amato, M.; Kabesch, M.; Melén, E.; Kere, J.; Pulkkinen, V. Neuropeptide S (NPS) variants modify the signaling and risk effects of NPS Receptor 1 (NPSR1) variants in asthma. PLoS ONE 2017, 12, e0176568. [Google Scholar] [CrossRef]
- Laitinen, T.; Polvi, A.; Rydman, P.; Vendelin, J.; Pulkkinen, V.; Salmikangas, P.; Mäkelä, S.; Rehn, M.; Pirskanen, A.; Rautanen, A.; et al. Characterization of a common susceptibility locus for asthma-related traits. Science 2004, 304, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Vendelin, J.; Pulkkinen, V.; Rehn, M.; Pirskanen, A.; Räisänen-Sokolowski, A.; Laitinen, A.; Laitinen, L.A.; Kere, J.; Laitinen, T. Characterization of GPRA, a novel G protein-coupled receptor related to asthma. Am. J. Respir. Cell. Mol. Biol. 2005, 33, 262–270. [Google Scholar] [CrossRef]
- Hamsten, C.; Häggmark, A.; Grundström, J.; Mikus, M.; Lindskog, C.; Konradsen, J.R.; Eklund, A.; Pershagen, G.; Wickman, M.; Grunewald, J.; et al. Protein profiles of CCL5, HPGDS, and NPSR1 in plasma reveal association with childhood asthma. Allergy 2016, 71, 1357–1361. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Perkins, C.; Mingler, M.K.; Finkelman, F.D.; Rothenberg, M.E. The role of neuropeptide S and neuropeptide S receptor 1 in regulation of respiratory function in mice. Peptides 2011, 32, 818–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilmarinen, P.; James, A.; Moilanen, E.; Pulkkinen, V.; Daham, K.; Saarelainen, S.; Laitinen, T.; Dahlén, S.E.; Kere, J.; Dahlén, B.; et al. Enhanced expression of neuropeptide S (NPS) receptor in eosinophils from severe asthmatics and subjects with total IgE above 100IU/ml. Peptides 2014, 51, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Wtorek, K.; Janecka, A. Potential of Nociceptin/Orphanin FQ Peptide Analogs for Drug Development. Chem. Biodivers. 2021, 18, e2000871. [Google Scholar] [CrossRef]
- D’Agostino, B.; Sgambato, M.; Esposito, R.; Spaziano, G. N/OFQ-NOP System and Airways. Handb. Exp. Pharmacol. 2019, 254, 313–322. [Google Scholar] [CrossRef]
- Shah, S.; Page, C.P.; Spina, D. Nociceptin inhibits non-adrenergic non-cholinergic contraction in guinea-pig airway. Br. J. Pharmacol. 1998, 125, 510–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Agostino, B.; Orlotti, D.; Calò, G.; Sullo, N.; Russo, M.; Guerrini, R.; De Nardo, M.; Mazzeo, F.; Candeletti, S.; Rossi, F. Nociceptin modulates bronchoconstriction induced by sensory nerve activation in mouse lung. Am. J. Respir. Cell. Mol. Biol. 2010, 42, 250–254. [Google Scholar] [CrossRef]
- McLeod, R.L.; Bolser, D.C.; Jia, Y.; Parra, L.E.; Mutter, J.C.; Wang, X.; Tulshian, D.B.; Egan, R.W.; Hey, J.A. Antitussive effect of nociceptin/orphanin FQ in experimental cough models. Pulm. Pharmacol. Ther. 2002, 15, 213–216. [Google Scholar] [CrossRef]
- Singh, S.R.; Sullo, N.; D’Agostino, B.; Brightling, C.E.; Lambert, D.G. The effects of nociceptin peptide (N/OFQ)-receptor (NOP) system activation in the airways. Peptides 2013, 39, 36–46. [Google Scholar] [CrossRef]
- Singh, S.R.; Sullo, N.; Matteis, M.; Spaziano, G.; McDonald, J.; Saunders, R.; Woodman, L.; Urbanek, K.; De Angelis, A.; De Palma, R.; et al. Nociceptin/orphanin FQ (N/OFQ) modulates immunopathology and airway hyperresponsiveness representing a novel target for the treatment of asthma. Br. J. Pharmacol. 2016, 173, 1286–1301. [Google Scholar] [CrossRef] [PubMed]
- Sullo, N.; Roviezzo, F.; Matteis, M.; Ianaro, A.; Calò, G.; Guerrini, R.; De Gruttola, L.; Spaziano, G.; Cirino, G.; Rossi, F.; et al. Nociceptin/orphanin FQ receptor activation decreases the airway hyperresponsiveness induced by allergen in sensitized mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 304, L657–L664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaziano, G.; Sorrentino, R.; Matteis, M.; Malgieri, G.; Sgambato, M.; Russo, T.P.; Terlizzi, M.; Roviezzo, F.; Rossi, F.; Pinto, A.; et al. Nociceptin reduces the inflammatory immune microenvironment in a conventional murine model of airway hyperresponsiveness. Clin. Exp. Allergy 2017, 47, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartaglione, G.; Spaziano, G.; Sgambato, M.; Russo, T.P.; Liparulo, A.; Esposito, R.; Mirra, S.; Filosa, R.; Roviezzo, F.; Polverino, F.; et al. Nociceptin/Orphanin Fq in inflammation and remodeling of the small airways in experimental model of airway hyperresponsiveness. Physiol. Rep. 2018, 6, e13906. [Google Scholar] [CrossRef]
- Jensen, R.T.; Battey, J.F.; Spindel, E.R.; Benya, R.V. International Union of Pharmacology. LXVIII. Mammalian bombesin receptors: Nomenclature, distribution, pharmacology, signaling, and functions in normal and disease states. Pharmacol. Rev. 2008, 60, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, I.D.; Weber, H.C. Biology of mammalian bombesin-like peptides and their receptors. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, R.L.; Torday, J.S.; Mu, Q.; Asokananthan, N.; Sikorski, K.A.; Sunday, M.E. Bombesin-like peptides and receptors in normal fetal baboon lung: Roles in lung growth and maturation. Am. J. Physiol. 1999, 277, L1003–L1017. [Google Scholar] [CrossRef]
- Kane, M.A.; Toi-Scott, M.; Johnson, G.L.; Kelley, K.K.; Boose, D.; Escobedo-Morse, A. Bombesin-like peptide receptors in human bronchial epithelial cells. Peptides 1996, 17, 111–118. [Google Scholar] [CrossRef]
- Shan, L.; Emanuel, R.L.; Dewald, D.; Torday, J.S.; Asokanathan, N.; Wada, K.; Wada, E.; Sunday, M.E. Bombesin-like peptide receptor gene expression, regulation, and function in fetal murine lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L165–L173. [Google Scholar] [CrossRef] [Green Version]
- Lach, E.; Haddad, E.B.; Gies, J.P. Contractile effect of bombesin on guinea pig lung in vitro: Involvement of gastrin-releasing peptide-preferring receptors. Am. J. Physiol. 1993, 264, L80–L86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Potts, E.N.; Cuttitta, F.; Foster, W.M.; Sunday, M.E. Gastrin-releasing peptide blockade as a broad-spectrum anti-inflammatory therapy for asthma. Proc. Natl. Acad. Sci. USA 2011, 108, 2100–2105. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, M.; Sugiyama, K.; Coy, D.H.; Kong, Y.; Miller, Y.E.; Weller, P.F.; Wada, K.; Wada, E.; Sunday, M.E. Bombesin-like peptides and mast cell responses: Relevance to bronchopulmonary dysplasia? Am. J. Respir. Crit. Care Med. 2003, 168, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.G.; Brenol, C.V.; Edelweiss, M.I.; Brenol, J.C.; Petronilho, F.; Roesler, R.; Dal-Pizzol, F.; Schwartsmann, G.; Xavier, R.M. Effects of an antagonist of the bombesin/gastrin-releasing peptide receptor on complete Freund’s adjuvant-induced arthritis in rats. Peptides 2008, 29, 1726–1731. [Google Scholar] [CrossRef] [PubMed]
- Dal-Pizzol, F.; Di Leone, L.P.; Ritter, C.; Martins, M.R.; Reinke, A.; Pens Gelain, D.; Zanotto-Filho, A.; de Souza, L.F.; Andrades, M.; Barbeiro, D.F.; et al. Gastrin-releasing peptide receptor antagonist effects on an animal model of sepsis. Am. J. Respir. Crit. Care Med. 2006, 173, 84–90. [Google Scholar] [CrossRef]
- Petronilho, F.; Roesler, R.; Schwartsmann, G.; Dal Pizzol, F. Gastrin-releasing peptide receptor as a molecular target for inflammatory diseases. Inflamm. Allergy Drug Targets 2007, 6, 197–200. [Google Scholar] [PubMed]
- Sunday, M.E.; Yoder, B.A.; Cuttitta, F.; Haley, K.J.; Emanuel, R.L. Bombesin-like peptide mediates lung injury in a baboon model of bronchopulmonary dysplasia. J. Clin. Investig. 1998, 102, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.Q.; Qu, X. Extraintestinal roles of bombesin-like peptides and their receptors: Lung. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 22–26. [Google Scholar] [CrossRef]
- Li, M.; Liang, P.; Liu, D.; Yuan, F.; Chen, G.C.; Zhang, L.; Liu, Y.; Liu, H. Bombesin Receptor Subtype-3 in Human Diseases. Arch. Med. Res. 2019, 50, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, M.; Tan, Y.; Xiang, Y.; Liu, H.; Qu, F.; Qin, L.; Qin, X. BRS-3 activation transforms the effect of human bronchial epithelial cells from PGE2 mediated inhibition to TGF-beta1 dependent promotion on proliferation and collagen synthesis of lung fibroblasts. Cell. Biol. Int. 2007, 31, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Peng, L.; Liu, C.; Tan, Y.; Xiang, Y.; Qu, X.; Weber, H.C.; Dai, M.; Qin, X. Activation of Bombesin Receptor Subtype-3 Promotes Antigen-Presenting Action in Human Bronchial Epithelial Cells. Int. Arch. Allergy Immunol. 2018, 175, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Gamse, R.; Leeman, S.E.; Holzer, P.; Lembeck, F. Differential effects of capsaicin on the content of somatostatin, substance P, and neurotensin in the nervous system of the rat. Naunyn Schmiedebergs Arch. Pharmacol. 1981, 317, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.C.; Greenwood, M.T.; Panetta, R.; Demchyshyn, L.; Niznik, H.; Srikant, C.B. The somatostatin receptor family. Life Sci. 1995, 57, 1249–1265. [Google Scholar] [CrossRef] [PubMed]
- ten Bokum, A.M.; Hofland, L.J.; van Hagen, P.M. Somatostatin and somatostatin receptors in the immune system: A review. Eur. Cytokine Netw. 2000, 11, 161–176. [Google Scholar] [PubMed]
- Hoyer, D.; Bell, G.I.; Berelowitz, M.; Epelbaum, J.; Feniuk, W.; Humphrey, P.P.; O’Carroll, A.M.; Patel, Y.C.; Schonbrunn, A.; Taylor, J.E.; et al. Classification and nomenclature of somatostatin receptors. Trends Pharmacol. Sci. 1995, 16, 86–88. [Google Scholar] [CrossRef]
- Reisine, T.; Bell, G.I. Molecular properties of somatostatin receptors. Neuroscience. 1995, 67, 777–790. [Google Scholar] [CrossRef]
- Hofland, L.J.; Visser-Wisselaar, H.A.; Lamberts, S.W. Somatostatin analogs: Clinical application in relation to human somatostatin receptor subtypes. Biochem. Pharmacol. 1995, 50, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Borie, R.; Fabre, A.; Prost, F.; Marchal-Somme, J.; Lebtahi, R.; Marchand-Adam, S.; Aubier, M.; Soler, P.; Crestani, B. Activation of somatostatin receptors attenuates pulmonary fibrosis. Thorax 2008, 63, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Varecza, Z.; Elekes, K.; László, T.; Perkecz, A.; Pintér, E.; Sándor, Z.; Szolcsányi, J.; Keszthelyi, D.; Szabó, A.; Sándor, K.; et al. Expression of the somatostatin receptor subtype 4 in intact and inflamed pulmonary tissues. J. Histochem. Cytochem. 2009, 57, 1127–1137. [Google Scholar] [CrossRef] [Green Version]
- Helyes, Z.; Elekes, K.; Németh, J.; Pozsgai, G.; Sándor, K.; Kereskai, L.; Börzsei, R.; Pintér, E.; Szabó, A.; Szolcsányi, J. Role of transient receptor potential vanilloid 1 receptors in endotoxin-induced airway inflammation in the mouse. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L1173–L1181. [Google Scholar] [CrossRef] [Green Version]
- Elekes, K.; Helyes, Z.; Kereskai, L.; Sándor, K.; Pintér, E.; Pozsgai, G.; Tékus, V.; Bánvölgyi, A.; Németh, J.; Szuts, T.; et al. Inhibitory effects of synthetic somatostatin receptor subtype 4 agonists on acute and chronic airway inflammation and hyperreactivity in the mouse. Eur. J. Pharmacol. 2008, 578, 313–322. [Google Scholar] [CrossRef]
- Helyes, Z.; Pintér, E.; Németh, J.; Kéri, G.; Thán, M.; Oroszi, G.; Horváth, A.; Szolcsányi, J. Anti-inflammatory effect of synthetic somatostatin analogues in the rat. Br. J. Pharmacol. 2001, 134, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Helyes, Z.; Pinter, E.; Nemeth, J.; Sandor, K.; Elekes, K.; Szabo, A.; Pozsgai, G.; Keszthelyi, D.; Kereskai, L.; Engstrom, M.; et al. Effects of the somatostatin receptor subtype 4 selective agonist J-2156 on sensory neuropeptide release and inflammatory reactions in rodents. Br. J. Pharmacol. 2006, 149, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Abad, C.; Gomariz, R.P.; Waschek, J.A. Neuropeptide mimetics and antagonists in the treatment of inflammatory disease: Focus on VIP and PACAP. Curr. Top. Med. Chem. 2006, 6, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Onoue, S.; Yamada, S.; Yajima, T. Bioactive analogues and drug delivery systems of vasoactive intestinal peptide (VIP) for the treatment of asthma/COPD. Peptides 2007, 28, 1640–1650. [Google Scholar] [CrossRef]
- Huang, M.C.; Miller, A.L.; Wang, W.; Kong, Y.; Paul, S.; Goetzl, E.J. Differential signaling of T cell generation of IL-4 by wild-type and short-deletion variant of type 2 G protein-coupled receptor for vasoactive intestinal peptide (VPAC2). J. Immunol. 2006, 176, 6640–6646. [Google Scholar] [CrossRef] [Green Version]
- Ghatei, M.A.; Springall, D.R.; Richards, I.M.; Oostveen, J.A.; Griffin, R.L.; Cadieux, A.; Polak, J.M.; Bloom, S.R. Regulatory peptides in the respiratory tract of Macaca fascicularis. Thorax 1987, 42, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Baraniuk, J.N.; Lundgren, J.D.; Okayama, M.; Mullol, J.; Merida, M.; Shelhamer, J.H.; Kaliner, M.A. Vasoactive intestinal peptide in human nasal mucosa. J. Clin. Investig. 1990, 86, 825–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howarth, P.H.; Springall, D.R.; Redington, A.E.; Djukanovic, R.; Holgate, S.T.; Polak, J.M. Neuropeptide-containing nerves in endobronchial biopsies from asthmatic and nonasthmatic subjects. Am. J. Respir. Cell. Mol. Biol. 1995, 13, 288–296. [Google Scholar] [CrossRef]
- Chanez, P.; Springall, D.; Vignola, A.M.; Moradoghi-Hattvani, A.; Polak, J.M.; Godard, P.; Bousquet, J. Bronchial mucosal immunoreactivity of sensory neuropeptides in severe airway diseases. Am. J. Respir. Crit. Care Med. 1998, 158, 985–990. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Springer, J.; Fischer, A. Vasoactive intestinal polypeptide as mediator of asthma. Pulm. Pharmacol. Ther. 2001, 14, 391–401. [Google Scholar] [CrossRef]
- Verma, A.K.; Manohar, M.; Upparahalli Venkateshaiah, S.; Mishra, A. Neuroendocrine cells derived chemokine vasoactive intestinal polypeptide (VIP) in allergic diseases. Cytokine Growth Factor. Rev. 2017, 38, 37–48. [Google Scholar] [CrossRef]
- Metwali, A.; Blum, A.M.; Ferraris, L.; Klein, J.S.; Fiocchi, C.; Weinstock, J.V. Eosinophils within the healthy or inflamed human intestine produce substance P and vasoactive intestinal peptide. J. Neuroimmunol. 1994, 52, 69–78. [Google Scholar] [CrossRef]
- Wershil, B.K.; Turck, C.W.; Sreedharan, S.P.; Yang, J.; An, S.; Galli, S.J.; Goetzl, E.J. Variants of vasoactive intestinal peptide in mouse mast cells and rat basophilic leukemia cells. Cell Immunol. 1993, 151, 369–378. [Google Scholar] [CrossRef]
- Leceta, J.; Martínez, C.; Delgado, M.; Garrido, E.; Gomariz, R.P. Expression of vasoactive intestinal peptide in lymphocytes: A possible endogenous role in the regulation of the immune system. Adv. Neuroimmunol. 1996, 6, 29–36. [Google Scholar] [CrossRef]
- Gonzalez-Rey, E.; Delgado, M. Role of vasoactive intestinal peptide in inflammation and autoimmunity. Curr. Opin. Investig. Drugs 2005, 6, 1116–1123. [Google Scholar] [PubMed]
- Sergejeva, S.; Hoshino, H.; Yoshihara, S.; Kashimoto, K.; Lötvall, J.; Lindén, A. A synthetic VIP peptide analogue inhibits neutrophil recruitment in rat airways in vivo. Regul. Pept. 2004, 117, 149–154. [Google Scholar] [CrossRef]
- Szema, A.M.; Hamidi, S.A.; Koller, A.; Martin, D.W. Vasoactive Intestinal Peptide Knockout (VIP KO) mouse model of sulfite-sensitive asthma: Up-regulation of novel lung carbonyl reductase. BMC Immunol. 2011, 12, 66. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Yen, J.H.; Ganea, D. A novel VIP signaling pathway in T cells cAMP-->protein tyrosine phosphatase (SHP-2?)-->JAK2/STAT4-->Th1 differentiation. Peptides 2007, 28, 1814–1824. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, E.J.; Voice, J.K.; Shen, S.; Dorsam, G.; Kong, Y.; West, K.M.; Morrison, C.F.; Harmar, A.J. Enhanced delayed-type hypersensitivity and diminished immediate-type hypersensitivity in mice lacking the inducible VPAC(2) receptor for vasoactive intestinal peptide. Proc. Natl. Acad. Sci. USA 2001, 98, 13854–13859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Wu, J.; Dong, J. Modified BuShenYiQi formula alleviates experimental allergic asthma in mice by negative regulation of type 2 innate lymphoid cells and CD4+ type 9 helper T cells and the VIP-VPAC2 signalling pathway. Pharm. Biol. 2021, 59, 1216–1232. [Google Scholar] [CrossRef]
- Voice, J.K.; Dorsam, G.; Lee, H.; Kong, Y.; Goetzl, E.J. Allergic diathesis in transgenic mice with constitutive T cell expression of inducible vasoactive intestinal peptide receptor. FASEB J. 2001, 15, 2489–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindén, A.; Hansson, L.; Andersson, A.; Palmqvist, M.; Arvidsson, P.; Löfdahl, C.G.; Larsson, P.; Lötvall, J. Bronchodilation by an inhaled VPAC(2) receptor agonist in patients with stable asthma. Thorax 2003, 58, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Szema, A.M.; Hamidi, S.A.; Lyubsky, S.; Dickman, K.G.; Mathew, S.; Abdel-Razek, T.; Chen, J.J.; Waschek, J.A.; Said, S.I. Mice lacking the VIP gene show airway hyperresponsiveness and airway inflammation, partially reversible by VIP. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L880–L886. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.S.; Krause, J.E. Structure, Expression, and Some Regulatory Mechanisms of the Rat Preprotachykinin Gene Encoding Substance P, Neurokinin A, Neuropeptide K, and Neuropeptide Gamma. J. Neurosci. 1990, 10, 2203–2214. [Google Scholar] [CrossRef]
- Page, N.M. Characterization of the Gene Structures, Precursor Processing and Pharmacology of the Endokinin Peptides. Vascul. Pharmacol. 2006, 45, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Klassert, T.E.; Sánchez, J.J.; Almeida, T.A.; Candenas, L.; Pinto, F.; Acosta, O.; Hernández, M. Common Variants of the Neuropeptide Expressing Tachykinin Genes and Susceptibility to Asthma: A Case-Control Study. J. Neuroimmunol. 2010, 227, 202–207. [Google Scholar] [CrossRef]
- Page, N.M.; Bell, N.J.; Gardiner, S.M.; Manyonda, I.T.; Brayley, K.J.; Strange, P.G.; Lowry, P.J. Characterization of the Endokinins: Human Tachykinins with Cardiovascular Activity. Proc. Natl. Acad. Sci. USA 2003, 100, 6245–6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taracanova, A.; Alevizos, M.; Karagkouni, A.; Weng, Z.; Norwitz, E.; Conti, P.; Leeman, S.E.; Theoharides, T.C. SP and IL-33 Together Markedly Enhance TNF Synthesis and Secretion from Human Mast Cells Mediated by the Interaction of Their Receptors. Proc. Natl. Acad. Sci. USA 2017, 114, E4002–E4009. [Google Scholar] [CrossRef] [Green Version]
- Lindén, A. Role of Interleukin-17 and the Neutrophil in Asthma. Int. Arch. Allergy Immunol. 2001, 126, 179–184. [Google Scholar] [CrossRef]
- Mizuta, K.; Gallos, G.; Zhu, D.; Mizuta, F.; Goubaeva, F.; Xu, D.; Panettieri, R.A., Jr.; Yang, J.; Emala, C.W., Sr. Expression and coupling of neurokinin receptor subtypes to inositol phosphate and calcium signaling pathways in human airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L523–L534. [Google Scholar] [CrossRef] [Green Version]
- Schelfhout, V.; Louis, R.; Lenz, W.; Heyrman, R.; Pauwels, R.; Joos, G. The Triple Neurokinin-Receptor Antagonist CS-003 Inhibits Neurokinin A-Induced Bronchoconstriction in Patients with Asthma. Pulm. Pharmacol. Ther. 2006, 19, 413–418. [Google Scholar] [CrossRef]
- Lundberg, J.M.; Brodin, E.; Saria, A. Effects and Distribution of Vagal Capsaicin-Sensitive Substance P Neurons with Special Reference to the Trachea and Lungs. Acta Physiol. Scand. 1983, 119, 243–252. [Google Scholar] [CrossRef]
- Lundberg, J.M.; Hökfelt, T.; Martling, C.R.; Saria, A.; Cuello, C. Substance P-Immunoreactive Sensory Nerves in the Lower Respiratory Tract of Various Mammals Including Man. Cell Tissue Res. 1984, 235, 251–261. [Google Scholar] [CrossRef]
- Carr, M.J.; Hunter, D.D.; Jacoby, D.B.; Undem, B.J. Expression of Tachykinins in Nonnociceptive Vagal Afferent Neurons during Respiratory Viral Infection in Guinea Pigs. Am. J. Respir. Crit. Care Med. 2002, 165, 1071–1075. [Google Scholar] [CrossRef]
- Hunter, D.D.; Myers, A.C.; Undem, B.J. Nerve Growth Factor-Induced Phenotypic Switch in Guinea Pig Airway Sensory Neurons. Am. J. Respir. Crit. Care Med. 2000, 161, 1985–1990. [Google Scholar] [CrossRef]
- Luts, A.; Uddman, R.; Alm, P.; Basterra, J.; Sundler, F. Peptide-Containing Nerve Fibers in Human Airways: Distribution and Coexistence Pattern. Int. Arch. Allergy Immunol. 1993, 101, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.W.; Kraft, M.; Krause, J.E.; Rex, M.D.; Martin, R.J. Substance P and Its Receptor Neurokinin 1 Expression in Asthmatic Airways. J. Allergy Clin. Immunol. 2000, 106, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Maghni, K.; Michoud, M.-C.; Alles, M.; Rubin, A.; Govindaraju, V.; Meloche, C.; Martin, J.G. Airway Smooth Muscle Cells Express Functional Neurokinin-1 Receptors and the Nerve-Derived Preprotachykinin-a Gene: Regulation by Passive Sensitization. Am. J. Respir. Cell Mol. Biol. 2003, 28, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Aliakbari, J.; Sreedharan, S.P.; Turck, C.W.; Goetzl, E.J. Selective Localization of Vasoactive Intestinal Peptide and Substance P in Human Eosinophils. Biochem. Biophys. Res. Commun. 1987, 148, 1440–1445. [Google Scholar] [CrossRef]
- Ho, W.Z.; Lai, J.P.; Zhu, X.H.; Uvaydova, M.; Douglas, S.D. Human Monocytes and Macrophages Express Substance P and Neurokinin-1 Receptor. J. Immunol. Baltim. Md 1950 1997, 159, 5654–5660. [Google Scholar]
- Lai, J.P.; Douglas, S.D.; Ho, W.Z. Human Lymphocytes Express Substance P and Its Receptor. J. Neuroimmunol. 1998, 86, 80–86. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Germonpré, P.R.; Everaert, E.G.; Carro-Muino, I.; De Veerman, M.; de Felipe, C.; Hunt, S.P.; Thielemans, K.; Joos, G.F.; Pauwels, R.A. Endogenously Produced Substance P Contributes to Lymphocyte Proliferation Induced by Dendritic Cells and Direct TCR Ligation. Eur. J. Immunol. 1999, 29, 3815–3825. [Google Scholar] [CrossRef]
- De Swert, K.O.; Joos, G.F. 9 - Role of Tachykinins in Asthma and Allergic Disease. In New Insights to Neuroimmune Biology; Berczi, I., Ed.; Elsevier: London, UK, 2010; pp. 177–192. ISBN 978-0-12-384691-4. [Google Scholar]
- Severini, C.; Improta, G.; Falconieri-Erspamer, G.; Salvadori, S.; Erspamer, V. The Tachykinin Peptide Family. Pharmacol. Rev. 2002, 54, 285–322. [Google Scholar] [CrossRef]
- Borbély, É.; Helyes, Z. Role of Hemokinin-1 in Health and Disease. Neuropeptides 2017, 64, 9–17. [Google Scholar] [CrossRef]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide Substance P and the Immune Response. Cell. Mol. Life Sci. CMLS 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.W.; Hey, J.A.; McLeod, R.; Minnicozzi, M.; Rizzo, C. Tachykinins in the Lungs. Drug News Perspect. 1998, 11, 480–489. [Google Scholar]
- Choi, J.Y.; Khansaheb, M.; Joo, N.S.; Krouse, M.E.; Robbins, R.C.; Weill, D.; Wine, J.J. Substance P Stimulates Human Airway Submucosal Gland Secretion Mainly via a CFTR-Dependent Process. J. Clin. Investig. 2009, 119, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Khansaheb, M.; Choi, J.Y.; Joo, N.S.; Yang, Y.-M.; Krouse, M.; Wine, J.J. Properties of Substance P-Stimulated Mucus Secretion from Porcine Tracheal Submucosal Glands. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L370–L379. [Google Scholar] [CrossRef] [Green Version]
- Thapaliya, M.; Chompunud Na Ayudhya, C.; Amponnawarat, A.; Roy, S.; Ali, H. Mast Cell-Specific MRGPRX2: A Key Modulator of Neuro-Immune Interaction in Allergic Diseases. Curr. Allergy Asthma Rep. 2021, 21, 3. [Google Scholar] [CrossRef]
- Evans, T.W.; Dixon, C.M.; Clarke, B.; Conradson, T.B.; Barnes, P.J. Comparison of Neurokinin A and Substance P on Cardiovascular and Airway Function in Man. Br. J. Clin. Pharmacol. 1988, 25, 273–275. [Google Scholar] [CrossRef]
- Hajna, Z.; Borbély, É.; Kemény, Á.; Botz, B.; Kereskai, L.; Szolcsányi, J.; Pintér, E.; Paige, C.J.; Berger, A.; Helyes, Z. Hemokinin-1 Is an Important Mediator of Endotoxin-Induced Acute Airway Inflammation in the Mouse. Peptides 2015, 64, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Grobman, M.; Graham, A.; Outi, H.; Dodam, J.R.; Reinero, C.R. Chronic Neurokinin-1 Receptor Antagonism Fails to Ameliorate Clinical Signs, Airway Hyper-Responsiveness or Airway Eosinophilia in an Experimental Model of Feline Asthma. J. Feline Med. Surg. 2016, 18, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Keith, I.M.; Pelto-Huikko, M.; Schalling, M.; Hökfelt, T. Calcitonin gene-related peptide and its mRNA in pulmonary neuroendocrine cells and ganglia. Histochemistry 1991, 96, 311–315. [Google Scholar] [CrossRef]
- Kajekar, R.; Myers, A.C. Calcitonin gene-related peptide affects synaptic and membrane properties of bronchial parasympathetic neurons. Respir. Physiol. Neurobiol. 2008, 160, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.P.; Rohde, J.; Tokuyama, K.; Barnes, P.J.; Rogers, D.F. Capsaicin and sensory neuropeptide stimulation of goblet cell secretion in Guinea-pig trachea. J. Physiol. 1990, 431, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.B.; Ali, F.R.; Heaney, L.G.; Benyahia, F.; Soh, C.P.C.; Renz, H.; Lee, T.H.; Larché, M. Airway expression of calcitonin gene-related peptide in T-cell peptide induced late asthmatic reactions in atopics. Allergy 2007, 62, 495–503. [Google Scholar] [CrossRef]
- Atanasova, K.R.; Reznikov, L.R. Neuropeptides in asthma, chronic obstructive pulmonary disease and cystic fibrosis. Respir. Res. 2018, 19, 149. [Google Scholar] [CrossRef] [Green Version]
- Tschirhart, E.; Bertrand, C.; Theodorsson, E.; Landry, Y. Evidence for the involvement of calcitonin gene-related peptide in the epithelium dependent contraction of Guinea-pig trachea in response to capsaicin. Naunyn Schmiedeberg’s Arch. Pharmacol. 1990, 342, 177–181. [Google Scholar] [CrossRef]
- Kanemura, T.; Tamaoki, J.; Horii, S.; Saka, N.; Kobayashi, K.; Isono, K.; Takeuchi, S.; Takizawa, T. Calcitonin gene-related peptide augments parasympathetic contraction of rabbit tracheal smooth muscle in vitro. Agents Actions 1990, 31, 219–224. [Google Scholar] [CrossRef]
- Dakhama, A.; Kanehiro, A.; Mäkelä, M.J.; Loader, J.E.; Larsen, G.L.; Gelfand, E.W. Regulation of airway hyperresponsiveness by calcitonin gene related peptide in allergen sensitized and challenged mice. Am. J. Respir. Crit. Care Med. 2002, 165, 1137–1144. [Google Scholar] [CrossRef]
- Rochlitzer, S.; Veres, T.Z.; Kühne, K.; Prenzler, F.; Pilzner, C.; Knothe, S.; Winkler, C.; Lauenstein, H.-D.; Willart, M.; Hammad, H.; et al. The neuropeptide calcitonin gene-related peptide affects allergic airway inflammation by modulating dendritic cell function. Clin. Exp. Allergy 2011, 41, 1609–1621. [Google Scholar] [CrossRef]
- Sood, A.; Shore, S.A. Adiponectin, Leptin, and Resistin in Asthma: Basic Mechanisms through Population Studies. J. Allergy 2013, 2013, 785835. [Google Scholar] [CrossRef] [Green Version]
- Bianco, A.; Nigro, E.; Monaco, M.L.; Matera, M.G.; Scudiero, O.; Mazzarella, G.; Daniele, A. The burden of obesity in asthma and COPD: Role of adiponectin. Pulm. Pharmacol. Therap. 2017, 43, 20–25. [Google Scholar] [CrossRef]
- Miller, M.; Cho, J.Y.; Pham, A.; Ramsdell, J.; Broide, D.H. Adiponectin and functional adiponectin receptor 1 are expressed by airway epithelial cells in chronic obstructive pulmonary disease. J. Immunol. 2009, 182, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, K.; Takeda, Y.; Tetsumoto, S.; Iwasaki, T.; Tsujino, K.; Kuhara, H.; Jin, Y.; Nagatomo, I.; Kida, H.; Goya, S.; et al. Involvement of endothelial apoptosis underlying chronic obstructive pulmonary disease-like phenotype in adiponectin-null mice: Implications for therapy. Am. J. Respir. Crit. Care Med. 2011, 183, 1164–1175. [Google Scholar] [CrossRef]
- Takeuchi, T.; Misaki, A.; Fujita, J.; Sonobe, H.; Ohtsuki, Y. T-cadherin (CDH13, Hcadherin) expression downregulated surfactant protein D in bronchioloalveolar cells. Virchows Arch. 2001, 438, 370–375. [Google Scholar] [CrossRef]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.S.; Lodish, H.F. T- cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.S.; Kasahara, D.I.; Verbout, N.G.; Fedulov, A.V.; Zhu, M.; Si, H.; Wurmbrand, A.P.; Hug, C.; Ranscht, B.; Shore, S.A. Role of the adiponectin binding protein, T-cadherin (Cdh13), in allergic airways responses in mice. PLoS ONE 2012, 7, e41088. [Google Scholar] [CrossRef]
- Ajuwon, K.M.; Spurlock, M.E. Adiponectin inhibits LPS-induced NF-𝜅B activation and IL-6 production and increases PPAR𝛾2 expression in adipocytes. Am. J. Physiol. Regulat. Integr. Comp. Physiol. 2005, 288, R1220–R1225. [Google Scholar] [CrossRef]
- Wulster-Radcliffe, M.C.; Ajuwon, K.M.; Wang, J.; Christian, J.A.; Spurlock, M.E. Adiponectin differentially regulates cytokines in porcine macrophages. Biochem. Biophys. Res. Commun. 2004, 316, 924–929. [Google Scholar] [CrossRef]
- Aydin, M.; Koca, C.; Ozol, D.; Uysal, S.; Yildirim, Z.; Sahin Kavakli, H.; Yigitoglu, M.R. Interaction of Metabolic Syndrome with Asthma in Postmenopausal Women: Role of Adipokines. Inflammation 2013, 36, 1232–1238. [Google Scholar] [CrossRef]
- Coffey, M.J.; Torretti, B.; Mancuso, P. Adipokines and Cysteinyl Leukotrienes in the Pathogenesis of Asthma. J. Allergy 2015, 157919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giouleka, P.; Papatheodorou, G.; Lyberopoulos, P.; Karakatsani, A.; Alchanatis, M.; Roussos, C.; Papiris, S.; Loukides, S. Body mass index is associated with leukotriene inflammation in asthmatics. Eur. J. Clin. Investig. 2011, 41, 30–38. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Toyomasu, K.; Uchimura, N.; Ishitake, T. Low-molecular-weight adiponectin is more closely associated with episodes of asthma than high-molecular-weight adiponectin. Endocr. J. 2013, 60, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Nigro, E.; Daniele, A.; Scudiero, O.; Monaco, M.L.; Roviezzo, F.; D’Agostino, B.; Mazzarella, G.; Bianco, A. Adiponectin in asthma: Implications for phenotyping. Curr. Protein Pept. Sci. 2015, 16, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, A.; Seagrave, J.; Herbert, G.; Harkins, M.; Qualls, C.; Schuyler, M. Asthma is associated with lower adiponectin concentrations in sputum than controls. Am. J. Respir. Crit. Care Med. 2012, 185. abstract (11)A6502. [Google Scholar]
- Sood, A.; Qualls, C.; Schuyler, M.; Thyagarajan, B.; Steffes, M.W.; Smith, L.J.; Jacobs, D.R., Jr. Low Serum Adiponectin Predicts Future Risk for Asthma in Women. Am. J. Respir. Crit. Care Med. 2012, 186, 41–47. [Google Scholar] [CrossRef]
- Baek, H.-S.; Kim, Y.-D.; Shin, J.-H.; Kim, J.-H.; Oh, J.-W.; Lee, H.-B. Serum leptin and adiponectin levels correlate with exercise-induced bronchoconstriction in children with asthma. Ann. Allergy Asthma Immunol. 2011, 107, 14–21. [Google Scholar] [CrossRef]
- Dogru, M.; Ozde, S.; Aktas, A.; Karatoprak, E.Y. The adiponectin levels and asthma control in non-obese children with asthma. J. Asthma 2015, 52, 772–776. [Google Scholar] [CrossRef]
- Kattan, M.; Kumar, R.; Bloomberg, G.R.; Mitchell, H.E.; Calatroni, A.; Gergen, P.J.; Kercsmar, C.M.; Visness, C.M.; Matsui, E.C.; Steinbach, S.F.; et al. Asthma control, adiposity, and adipokines among inner-city adolescents. J. Allergy Clin. Immunol. 2010, 125, 584–592. [Google Scholar] [CrossRef] [Green Version]
- Nagel, G.; Koenig, W.; Rapp, K.; Wabitsch, M.; Zoellner, I.; Weiland, S.K. Associations of adipokines with asthma, rhinoconjunctivitis, and eczema in German schoolchildren. Pediatr. Allergy Immunol. 2009, 20, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Rothenbacher, D.; Weyermann, M.; Fantuzziw, G.; Brenner, H. Adipokines in cord blood and risk of wheezing disorders within the first two years of life. Clin. Exp. Allergy 2007, 37, 1143–1149. [Google Scholar] [CrossRef]
- Sood, A.; Cui, X.; Qualls, C.; Beckett, W.S.; Gross, M.D.; Steffes, M.W.; Smith, L.J.; Jacobs, D.R., Jr. Association between asthma and serum adiponectin concentration in women. Thorax 2008, 63, 877–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Yang, H.; He, H.; Shi, H.; He, P.; Yan, S.; Jin, T. Plasma Adiponectin Concentrations and Adiponectin Gene Polymorphisms Are Associated with Bronchial Asthma in the Chinese Li Population. Iran J. Allergy Asthma Immunol. 2015, 14, 292–297. [Google Scholar]
- Baltieri, L.; Cazzo, E.; Modena, D.A.O.; Gobato-Rentel, R.C.; Martins, L.C.; Chaim, E.A. Correlation between levels of adipokines and inflammatory mediators with spirometric parameters in individuals with obesity and symptoms of asthma: Cross-sectional study. Pulmonology 2020, 16. [Google Scholar] [CrossRef]
- Kalmarzi, R.N.; Ataee, P.; Mansori, M.; Moradi, G.; Ahmadi, S.; Kaviani, Z.; Khalafi, B.; Kooti, W. Serum levels of adiponectin and leptin in asthmatic patients and its relation with asthma severity, lung function and BMI. Allergol. Immunopathol. (Madr) 2017, 45, 258–264. [Google Scholar] [CrossRef]
- Mikalsen, I.B.; Byberg, K.; Forman, M.R.; Øymar, K. Adipokines in adolescence; the associations with lung function and atopy - A cross-sectional study. Respir. Med. 2020, 170, 106063. [Google Scholar] [CrossRef]
- Nigro, E.; Matteis, M.; Roviezzo, F.; Mattera Iacono, V.; Scudiero, O.; Spaziano, G.; Tartaglione, G.; Urbanek, K.; Filosa, R.; Daniele, A.; et al. Role of adiponectin in sphingosine-1-phosphate induced airway hyperresponsiveness and inflammation. Pharmacol. Res. 2016, 103, 114–122. [Google Scholar] [CrossRef]
- Shore, S.A.; Terry, R.D.; Flynt, L.; Xu, A.; Hug, C. Adiponectin attenuates allergen-induced airway inflammation and hyperresponsiveness in mice. J. Allergy Clin. Immunol. 2006, 118, 389–395. [Google Scholar] [CrossRef]
- Otelea, M.R.; Arghir, O.C.; Zugravu, C.; Rascu, A. Adiponectin and Asthma: Knowns, Unknowns and Controversies. Int. J. Mol. Sci. 2021, 22, 8971. [Google Scholar] [CrossRef]
- Hsu, A.; Aronoff, D.M.; Phipps, J.; Goel, D.; Mancuso, P. Leptin improves pulmonary bacterial clearance and survival in ob/ob mice during pneumococcal pneumonia. Clin. Exp. Immunol. 2007, 150, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, P.; Gottschalk, A.; Phare, S.M.; Peters-Golden, M.; Lukacs, N.W.; Huffnagle, G.B. Leptin-deficient mice exhibit impaired host defense in Gram-negative pneumonia. J. Immunol. 2002, 168, 4018–4024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forny-Germano, L.; De Felice, F.G.; Vieira, M.N. The Role of Leptin and Adiponectin in Obesity-Associated Cognitive Decline and Alzheimer’s Disease. Front. Neurosci. 2019, 12, 1027. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Amling, M.; Takeda, S.; Priemel, M.; Schilling, A.F.; Beil, F.T.; Shen, J.; Vinson, C.; Rueger, J.M.; Karsenty, G. Leptin inhibits bone formation through a hypothalamic relay: A central control of bone mass. Cell 2000, 100, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Matarese, G.; Moschos, S.; Mantzoros, C.S. Leptin in immunology. J. Immunol. 2005, 174, 3137–3142. [Google Scholar] [CrossRef] [Green Version]
- Vernooy, J.H.J.; Ubags, N.D.J.; Brusselle, G.G.; Tavernier, J.; Suratt, B.T.; Joos, G.F.; Wouters, E.F.M.; Bracke, K.R. Leptin as regulator of pulmonary immune responses: Involvement in respiratory diseases. Pulm. Pharmacol. Ther. 2013, 26, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Bruno, A.; Pace, E.; Chanez, P.; Gras, D.; Vachier, I.; Chiappara, G.; La Guardia, M.; Gerbino, S.; Profita, M.; Gjomarkaj, M. Leptin and leptin receptor expression in asthma. J. Allergy Clin. Immunol. 2009, 124, 230–237. [Google Scholar] [CrossRef]
- Holguin, F.; Rojas, M.; Brown, L.; Fitzpatrick, A.M. Airway and Plasma Leptin and Adiponectin in Lean and Obese Asthmatics and Controls. J. Asthma 2011, 48, 217–223. [Google Scholar] [CrossRef]
- Nair, P.; Radford, K.; Fanat, A.; Janssen, L.J.; Peters-Golden, M.; Cox, P.G. The effects of leptin on airway smooth muscle responses. Am. J. Respir. Cell. Mol. Biol. 2008, 39, 475–481. [Google Scholar] [CrossRef]
- Cvejoska Cholakovska, V.; Vlashki, E.; Kochova, M.; Velikj Stefanovska, V.; Petlichkovski, A. Leptin, Obesity Parameters, and Atopy among Children with Asthma. Pril (Makedon Akad Nauk Umet Odd Med Nauki) 2021, 42, 79–88. [Google Scholar] [CrossRef]
- Doaa, M.Y.; Rabab, M.E.; Dina, M.S.; Eman, M.E. The influence of leptin on Th1/Th2 balance in obese children with asthma. J. Bras. Pneumol. 2013, 39, 562–568. [Google Scholar]
- Lessard, A.; St-Laurent, J.; Turcotte, H.; Boulet, L.-P. Leptin and adiponectin in obese and non-obese subjects with asthma. Biomarkers 2011, 16, 271–273. [Google Scholar] [CrossRef]
- Ma, C.; Wang, Y.; Xue, M. Correlations of severity of asthma in children with body mass index, adiponectin and leptin. J. Clin. Lab. Anal. 2019, 33, e22915. [Google Scholar] [CrossRef]
- Morishita, R.; do Carmo Franco, M.; Suano-Souza, F.I.; Solé, D.; Fiorini Puccini, R.; Louzada Strufaldi, M.W. Body mass index, adipokines and insulin resistance in asthmatic children and adolescents. J. Asthma 2016, 53, 478–484. [Google Scholar] [CrossRef]
- Newson, R.B.; Jones, M.; Forsberg, B.; Janson, C.; Bossios, A.; Dahlen, S.-E.; Toskala, E.M.; Al-Kalemji, A.; Kowalski, M.L.; Rymarczyk, B.; et al. The association of asthma, nasal allergies, and positive skin prick tests with obesity, leptin, and adiponectin. Clin. Exp. Allergy 2014, 44, 250–260. [Google Scholar] [CrossRef]
- Tanju, A.; Cekmez, F.; Aydinoz, S.; Karademir, F.; Suleymanoglu, S.; Gocmen, I. Association Between Clinical Severity of Childhood Asthma and Serum Leptin Levels. Indian J. Pediatr. 2011, 78, 291–295. [Google Scholar] [CrossRef]
- Leivo-Korpela, S.; Lehtimäki, L.; Vuolteenaho, K.; Nieminen, R.; Kankaanranta, H.; Saarelainen, S.; Moilanen, E. Adipokine resistin predicts anti-inflammatory effect of glucocorticoids in asthma. J. Inflamm. (Lond.) 2011, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Leão da Silva, P.; de Mello, M.T.; Cheik, N.C.; Sanches, P.L.; Munhoz da Silveira Campos, R.; Carnier, J.; Inoue, D.; do Nascimento, C.M.; Oyama, L.M.; Tock, L.; et al. Reduction in the leptin concentration as a predictor of improvement in lung function in obese adolescents. Obes. Facts 2012, 5, 806–820. [Google Scholar] [CrossRef] [Green Version]
- Al-Ayed, M.; Alshaybari, K.; Alshehri, J.A.; Nasser, I.; Alaamri, H.; Alaseeri, W.; Mahfouz, A.A.; Alsareli, S.A.; Asaad, A.M.; Magzoub, A.A.; et al. Obesity and childhood asthma in male schoolchildren in Saudi Arabia: Is there a role for leptin, interleukin-4, interleukin-5, and interleukin-21? Ann. Saudi. Med. 2019, 39, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Vassiliou, A.G.; Vitsas, V.; Kardara, M.; Keskinidou, C.; Michalopoulou, P.; Rovina, N.; Dimopoulou, I.; Orfanos, S.E.; Tsoukalas, G.; Koutsoukou, A.; et al. Study of inflammatory biomarkers in COPD and asthma exacerbations. Adv. Respir. Med. 2020, 88, 558–566. [Google Scholar] [CrossRef]
- Castro-Rodriguez, J.A.; Forno, E.; Casanello, P.; Padilla, O.; Krause, B.J.; Uauy, R. Leptin in Cord Blood Associates with Asthma Risk at Age 3 in the Offspring of Gestational Obesity. Ann. Am. Thorac. Soc. 2020, 17, 1583–1589. [Google Scholar] [CrossRef]
- Rastogi, D.; Lang, J. Leptin as a Predictor of Incident Asthma in Offspring of Obese Mothers. Ann. Am. Thorac. Soc. 2020, 17, 1530–1532. [Google Scholar] [CrossRef]
- Jang, A.-S.; Kim, T.-H.; Park, J.-S.; Kim, K.-U.; Uh, S.-T.; Seo, K.-H.; Kim, Y.H.; Lim, H.I.; Park, C.-S. Association of Serum Leptin and Adiponectin with Obesity in Asthmatics. J. Asthma 2009, 46, 59–63. [Google Scholar] [CrossRef]
- Jartti, T.; Saarikoski, L.; Jartti, L.; Lisinen, I.; Jula, A.; Huupponen, R.; Viikari, J.; Raitakari, O.T. Obesity, adipokines and asthma. Allergy 2009, 64, 770–777. [Google Scholar] [CrossRef]
- Arshi, M.; Cardinal, J.; Hill, R.J.; Davies, P.S.; Wainwright, C. Asthma and insulin resistance in children. Respirology 2010, 15, 779–784. [Google Scholar] [CrossRef]
- Kim, K.W.; Shin, Y.H.; Lee, K.E.; Kim, E.S.; Sohn, M.H.; Kim, K.E. Relationship between adipokines and manifestations of childhood asthma. Pediatr. Allergy Immunol. 2008, 19, 535–540. [Google Scholar] [CrossRef]
- Muc, M.; Todo-Bom, A.; Mota-Pinto, A.; Vale-Pereira, S.; Loureiro, C. Leptin and resistin in overweight patients with and without asthma. Allergol. Immunopathol. (Madr) 2014, 42, 415–421. [Google Scholar] [CrossRef]
- Sutherland, T.J.T.; Sears, M.R.; McLachlan, C.R.; Poulton, R.; Hancox, R.J. Leptin, adiponectin, and asthma: Findings from a population-based cohort study. Ann. Allergy Asthma Immunol. 2009, 103, 101–107. [Google Scholar] [CrossRef]
- Arteaga-Solis, E.; Zee, T.; Emala, C.W.; Vinson, C.; Wess, J.; Karsenty, G. Inhibition of Leptin Regulation of Parasympathetic Signaling as a Cause of Extreme Body Weight-Associated Asthma. Cell Metab. 2013, 17, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Polotsky, M.; Elsayed-Ahmed, A.S.; Pichard, L.; Harris, C.C.; Smith, P.L.; Schneider, H.; Kirkness, J.P.; Polotsky, V.; Schwartz, A.R. Effects of leptin and obesity on the upper airway function. J. Appl. Physiol. 2012, 112, 1637–1643. [Google Scholar] [CrossRef] [Green Version]
- Shore, S.A.; Fredberg, J.J. Obesity, smooth muscle, and airway hyperresponsiveness. J. Allergy Clin. Immunol. 2005, 115, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Shore, S.A.; Rivera-Sanchez, Y.M.; Schwartzman, I.N.; Johnston, R.A. Responses to ozone are increased in obese mice. J. Appl. Physiol. 2003, 95, 938–945. [Google Scholar] [CrossRef]
- Shore, S.A.; Schwartzman, I.N.; Mellema, M.S.; Flynt, L.; Imrich, A.; Johnston, R.A. Effect of leptin on allergic airway responses in mice. J. Allergy Clin. Immunol. 2005, 115, 103–109. [Google Scholar] [CrossRef]
- Lu, F.L.; Johnston, R.A.; Flynt, L.; Theman, T.A.; Terry, R.D.; Schwartzman, I.N.; Lee, A.; Shore, S.A. Increased pulmonary responses to acute ozone exposure in obese db/db mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L856–L865. [Google Scholar] [CrossRef] [Green Version]
- Johnston, R.A.; Theman, T.A.; Lu, F.L.; Terry, R.D.; Williams, E.S.; Shore, S.A. Diet-induced obesity causes innate airway hyperresponsiveness to methacholine and enhances ozone-induced pulmonary inflammation. J. Appl. Physiol. 2008, 104, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Cheung, P.F.; Lam, C.W. Leptin-mediated cytokine release and migration of eosinophils: Implications for immunopathophysiology of allergic inflammation. Eur. J. Immunol. 2007, 37, 2337–2348. [Google Scholar] [CrossRef]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Lehrke, M.; Reilly, M.P.; Millington, S.C.; Iqbal, N.; Rader, D.J.; Lazar, M.A. An inflammatory cascade leading to hyperresistinemia in humans. PLoS Med. 2004, 1, e45. [Google Scholar] [CrossRef]
- Larochelle, J.; Freiler, J.; Dice, J.; Hagan, L. Plasma resistin levels in asthmatics as a marker of disease state. J. Asthma 2007, 44, 509–513. [Google Scholar] [CrossRef]
- Mishra, A.; Wang, M.; Schlotman, J.; Nikolaidis, N.M.; DeBrosse, C.W.; Karow, M.L.; Rothenberg, M.E. Resistin-like molecule-beta is an allergen-induced cytokine with inflammatory and remodeling activity in the murine lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L305–L313. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Johns, R.A. Resistin family proteins in pulmonary diseases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L422–L434. [Google Scholar] [CrossRef]
- Kwak, S.; Kim, Y.D.; Na, H.G.; Bae, C.H.; Song, S.Y.; Choi, Y.S. Resistin upregulates MUC5AC/B mucin gene expression in human airway epithelial cells. Biochem. Biophys. Res. Commun. 2018, 499, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Silswal, N.; Singh, A.K.; Aruna, B.; Mukhopadhyay, S.; Ghosh, S.; Ehtesham, N.Z. Human resistin stimulates the pro-inflammatory cytokines TNF-alpha and IL-12 in macrophages by NF-kappaB-dependent pathway. Biochem. Biophys. Res. Commun. 2005, 334, 1092–1101. [Google Scholar] [CrossRef]
- Lu, S.C.; Shieh, W.Y.; Chen, C.Y.; Hsu, S.C.; Chen, H.L. Lipopolysaccharide increases resistin gene expression in vivo and in vitro. FEBS Lett. 2002, 530, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Hirai, H.; Satoh, H.; Kudoh, A.; Watanabe, T. Interaction between resistin and adiponectin in the proliferation of rat vascular smooth muscle cells. Mol. Cell. Endocrinol. 2013, 366, 108–116. [Google Scholar] [CrossRef]
- Filková, M.; Haluzík, M.; Gay, S.; Senolt, L. The role of resistin as a regulator of inflammation: Implications for various human pathologies. Clin. Immunol. 2009, 133, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Pang, S.S.; Le, Y.Y. Role of resistin in inflammation and inflammation-related diseases. Cell. Mol. Immunol. 2006, 3, 29–34. [Google Scholar] [PubMed]
- Zhang, L.; Wang, M.; Kang, X.; Boontheung, P.; Li, N.; Nel, A.E.; Loo, J.A. Oxidative stress and asthma: Proteome analysis of chitinase-like proteins and FIZZ1 in lung tissue and bronchoalveolar lavage fluid. J. Proteome Res. 2009, 8, 1631–1638. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Wang, S.J.; Camoretti-Mercado, B.; Li, H.J.; Chen, M.; Bi, W.X. FIZZ1 plays a crucial role in early stage airway remodeling of OVA-induced asthma. J Asthma 2008, 45, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, S.; Hirose, K.; Takahashi, K.; Tamachi, T.; Ikeda, K.; Tokoyoda, K.; Nakayama, T.; Nakajima, H. Interleukin-25 induces pulmonary arterial remodeling via natural killer T cell-dependent mechanisms. Int. Arch. Allergy Immunol. 2013, 161, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Meng, Q.; Wu, H.; Eid, G.; Zhang, G.; Zhang, X.; Yang, S.; Huang, K.; Lee, T.H.; Corrigan, C.J.; et al. Resistin-like molecule-β is a human airway remodelling mediator. Eur. Respir. J. 2012, 39, 458–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, C.L.; Yin, L.J.; Sharma, S.; Kierstein, S.; Wu, H.F.; Eid, G.; Haczku, A.; Corrigan, C.J.; Ying, S. Resistin-like molecule-β (RELM-β) targets airways fibroblasts to effect remodelling in asthma: From mouse to man. Clin. Exp. Allergy. 2015, 45, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.R.; Shim, D.; Yoon, J.; Jang, H.S.; Oh, S.W.; Suh, S.H.; Choi, J.H.; Oh, G.T. Retnla overexpression attenuates allergic inflammation of the airway. PLoS ONE 2014, 9, e112666. [Google Scholar] [CrossRef]
- Liu, T.; Dhanasekaran, S.M.; Jin, H.; Hu, B.; Tomlins, S.A.; Chinnaiyan, A.M.; Phan, S.H. FIZZ1 stimulation of myofibroblast differentiation. Am. J. Pathol. 2004, 164, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- LeMessurier, K.S.; Palipane, M.; Tiwary, M.; Gavin, B.; Samarasinghe, A.E. Chronic features of allergic asthma are enhanced in the absence of resistin-like molecule-beta. Sci. Rep. 2018, 8, 7061. [Google Scholar] [CrossRef] [Green Version]
- Pine, G.M.; Batugedara, H.M.; Nair, M.G. Here, there and everywhere: Resistin-like molecules in infection, inflammation, and metabolic disorders. Cytokine 2018, 110, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Meuchel, L.W.; Su, Q.; Angelini, D.J.; Zhang, A.; Cheadle, C.; Kolosova, I.; Makarevich, O.D.; Yamaji-Kegan, K.; Rothenberg, M.E.; et al. Resistin-Like Molecule α in Allergen-Induced Pulmonary Vascular Remodeling. Am. J. Respir. Cell. Mol. Biol. 2015, 53, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Al Mutairi, S.S.; Mojiminiyi, O.A.; Shihab-Eldeen, A.; Al Rammah, T.; Abdella, N. Putative roles of circulating resistin in patients with asthma, COPD and cigarette smokers. Dis. Markers 2011, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, D.; Scott, H.; MacDonald-Wicks, L.; Gibson, P.G.; Wood, L.G. Resistin is a predictor of asthma risk and resistin:adiponectin ratio is a negative predictor of lung function in asthma. Clin. Exp. Allergy 2016, 46, 1056–1065. [Google Scholar] [CrossRef] [Green Version]
- Vezir, E.; Civelek, E.; Dibek Misirlioglu, E.; Toyran, M.; Capanoglu, M.; Karakus, E.; Kahraman, T.; Ozguner, M.; Demirel, F.; Gursel, I.; et al. Effects of Obesity on Airway and Systemic Inflammation in Asthmatic Children. Int. Arch. Allergy Immunol. 2021, 182, 679–689. [Google Scholar] [CrossRef]
- Grainge, C.; Dulay, V.; Ward, J.; Sammut, D.; Davies, E.; Green, B.; Lau, L.; Cottey, L.; Haitchi, H.M.; Davies, D.E.; et al. Resistin-like molecule-β is induced following bronchoconstriction of asthmatic airways. Respirology 2012, 17, 1094–1100. [Google Scholar] [CrossRef]
- Baatar, D.; Patel, K.; Taub, D.D. The effects of ghrelin on inflammation and the immune system. Mol. Cell. Endocrinol. 2011, 340, 44–58. [Google Scholar] [CrossRef]
- Wang, H.; Yang, T.; Shen, Y.; Wan, C.; Li, X.; Li, D.; Liu, Y.; Wang, T.; Xu, D.; Wen, F.; et al. Ghrelin Inhibits Interleukin-6 Production Induced by Cigarette Smoke Extract in the Bronchial Epithelial Cell Via NF-κB Pathway. Inflammation 2016, 39, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Wang, L.; Zeng, Q.; Zhang, Y.; Sheng, B.; Han, L. Ghrelin Ameliorates Asthma by Inhibiting Endoplasmic Reticulum Stress. Am. J. Med. Sci. 2017, 354, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Kangawa, K. Ghrelin: Structure and function. Physiol. Rev. 2005, 85, 495–522. [Google Scholar] [CrossRef] [PubMed]
- Ueberberg, B.; Unger, N.; Saeger, W.; Mann, K.; Petersenn, S. Expression of ghrelin and its receptor in human tissues. Horm. Metab. Res. 2009, 41, 814–821. [Google Scholar] [CrossRef]
- Volante, M.; Fulcheri, E.; Allìa, E.; Cerrato, M.; Pucci, A.; Papotti, M. Ghrelin expression in fetal, infant, and adult human lung. J. Histochem. Cytochem. 2002, 50, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Dong, W.; Zhou, M.; Zhang, F.; Marini, C.P.; Ravikumar, T.S.; Wang, P. Ghrelin attenuates sepsis-induced acute lung injury and mortality in rats. Am. J. Respir. Crit. Care Med. 2007, 176, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Dixit, V.D.; Schaffer, E.M.; Pyle, R.S.; Collins, G.D.; Sakthivel, S.K.; Palaniappan, R.; Lillard, J.W., Jr.; Taub, D.D. Ghrelin inhibits leptin- and activation-induced proinflammatory cytokine expression by human monocytes and T cells. J. Clin. Investig. 2004, 114, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Li, W.G.; Gavrila, D.; Liu, X.; Wang, L.; Gunnlaugsson, S.; Stoll, L.L.; McCormick, M.L.; Sigmund, C.D.; Tang, C.; Weintraub, N.L. Ghrelin inhibits proinflammatory responses and nuclear factor-kappaB activation in human endothelial cells. Circulation 2004, 109, 2221–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizaki, T.; Maegawa, T.; Sakurai, T.; Ogasawara, J.E.; Ookawara, T.; Ohishi, S.; Izawa, T.; Haga, S.; Ohno, H. Voluntary exercise attenuates obesity-associated inflammation through ghrelin expressed in macrophages. Biochem. Biophys. Res. Commun. 2011, 413, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Zhou, M.; Das, P.; Dong, W.; Ji, Y.; Yang, D.; Miksa, M.; Zhang, F.; Ravikumar, T.S.; Wang, P. Ghrelin inhibits sympathetic nervous activity in sepsis. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1697–E1702. [Google Scholar] [CrossRef]
- Xia, Q.; Pang, W.; Pan, H.; Zheng, Y.; Kang, J.S.; Zhu, S.G. Effects of ghrelin on the proliferation and secretion of splenic T lymphocytes in mice. Regul. Pept. 2004, 122, 173–178. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, S.; Yu, F.; Li, H.; Guo, C.; Fan, X. Ghrelin Attenuates Liver Fibrosis through Regulation of TGF-β1 Expression and Autophagy. Int. J. Mol. Sci. 2015, 16, 21911–21930. [Google Scholar] [CrossRef] [Green Version]
- Ota, Y.; Kawaguchi, Y.; Takagi, K.; Ichida, H.; Gono, T.; Hanaoka, M.; Higuchi, T.; Yamanaka, H. Ghrelin attenuates collagen production in lesional fibroblasts from patients with systemic sclerosis. Clin. Immunol. 2013, 147, 71–78. [Google Scholar] [CrossRef]
- Zhang, G.G.; Cai, H.Q.; Li, Y.H.; Sui, Y.B.; Zhang, J.S.; Chang, J.R.; Ning, M.; Wu, Y.; Tang, C.S.; Qi, Y.F.; et al. Ghrelin protects heart against ERS-induced injury and apoptosis by activating AMP-activated protein kinase. Peptides 2013, 48, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Imazu, Y.; Yanagi, S.; Miyoshi, K.; Tsubouchi, H.; Yamashita, S.; Matsumoto, N.; Ashitani, J.; Kangawa, K.; Nakazato, M. Ghrelin ameliorates bleomycin-induced acute lung injury by protecting alveolar epithelial cells and suppressing lung inflammation. Eur. J. Pharmacol. 2011, 672, 153–158. [Google Scholar] [CrossRef]
- Tsubouchi, H.; Yanagi, S.; Miura, A.; Iizuka, S.; Mogami, S.; Yamada, C.; Hattori, T.; Nakazato, M. Rikkunshito ameliorates bleomycin-induced acute lung injury in a ghrelin-independent manner. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L233–L245. [Google Scholar] [CrossRef]
- Sato, T.; Nakamura, Y.; Shiimura, Y.; Ohgusu, H.; Kangawa, K.; Kojima, M. Structure, regulation and function of ghrelin. J. Biochem. 2012, 151, 119–128. [Google Scholar] [CrossRef]
- Wang, H.; Dou, S.; Zhu, J.; Shao, Z.; Wang, C.; Cheng, B. Regulatory effects of ghrelin on endoplasmic reticulum stress, oxidative stress, and autophagy: Therapeutic potential. Neuropeptides 2021, 85, 102112. [Google Scholar] [CrossRef]
- Jafari, A.; Sadeghpour, S.; Ghasemnejad-Berenji, H.; Pashapour, S.; Ghasemnejad-Berenji, M. Potential Antioxidative, Anti-inflammatory and Immunomodulatory Effects of Ghrelin, an Endogenous Peptide from the Stomach in SARS-CoV2 Infection. Int. J. Pept. Res. Ther. 2021, 16, 1–9. [Google Scholar] [CrossRef]
- Tsaroucha, A.; Daniil, Z.; Malli, F.; Georgoulias, P.; Minas, M.; Kostikas, K.; Bargiota, A.; Zintzaras, E.; Gourgoulianis, K.I. Leptin, adiponectin, and ghrelin levels in female patients with asthma during stable and exacerbation periods. J. Asthma 2013, 50, 188–197. [Google Scholar] [CrossRef]
- Yuksel, H.; Sogut, A.; Yilmaz, O.; Onur, E.; Dinc, G. Role of adipokines and hormones of obesity in childhood asthma. Allergy Asthma Immunol. Res. 2012, 4, 98–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, K.; Nishi, Y.; Okamatsu, Y.; Kojima, M.; Matsuishi, T. Ghrelin and leptin: A link between obesity and allergy? J. Allergy Clin. Immunol. 2006, 117, 705–706. [Google Scholar] [CrossRef] [PubMed]
- Al-Ayed, M.S.; Al-Shaibari, K.S.; Alshehri, D.; Alzahrani, M.J.; Nasser, I.; Alaamri, H.S.; Alaseeri, W.A.; Mahfouz, A.A.; Alsareii, S.A.; Asaad, A.M.; et al. Serum Ghrelin Levels in Saudi Obese Asthmatic School-Children-Correlation with Interleukin-4, Interleukin-5, and Interleukin-21. Int. J. Environ. Res. Public Health 2020, 17, 1656. [Google Scholar] [CrossRef] [Green Version]
- Toru, Ü.; Ayada, C.; Genç, O.; Şahin, S.; Arık, Ö.; Acat, M.; Bulut, İ.; Çetinkaya, E. Visfatin and ghrelin: Can they be forthcoming biomarkers or new drug targets for asthma? Int. J. Clin. Exp. Med. 2015, 8, 6257–6261. [Google Scholar] [PubMed]
- Kodama, T.; Ashitani, J.; Matsumoto, N.; Kangawa, K.; Nakazato, M. Ghrelin treatment suppresses neutrophil-dominant inflammation in airways of patients with chronic respiratory infection. Pulm. Pharmacol. Ther. 2008, 21, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Miki, K.; Maekura, R.; Nagaya, N.; Nakazato, M.; Kimura, H.; Murakami, S.; Ohnishi, S.; Hiraga, T.; Miki, M.; Kitada, S.; et al. Ghrelin treatment of cachectic patients with chronic obstructive pulmonary disease: A multicenter, randomized, double-blind, placebo-controlled trial. PLoS ONE 2012, 7, e35708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pałgan, K.; Tykwińska, M.; Bartuzi, Z. Antimicrobial peptides in asthma pathogenesis. Postępy Higieny i Medycyny Doświadczalnej 2015, 69, 10–13. [Google Scholar] [CrossRef]
- Lecaille, F.; Lalmanach, G.; Andrault, P.M. Antimicrobial proteins and peptides in human lung diseases: A friend and foe partnership with host proteases. Biochimie 2016, 122, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, C.; Bals, R. Antimicrobial peptides in lung inflammation. Chem. Immunol. Allergy 2005, 86, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Wong, C.K.; Tsang, M.S.; Chu, I.M.; Liu, D.; Zhu, J.; Chu, M.; Lam, C.W. Activation of Eosinophils Interacting with Bronchial Epithelial Cells by Antimicrobial Peptide LL-37: Implications in Allergic Asthma. Sci. Rep. 2017, 7, 1848. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Dahlén, B.; Agerberth, B.; Haeggström, J.Z. The antimicrobial peptide LL-37 induces synthesis and release of cysteinyl leukotrienes from human eosinophils--implications for asthma. Allergy 2013, 68, 304–311. [Google Scholar] [CrossRef]
- Buss, W.W.; Lemanske, R.J.; Gern, J.E. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet 2010, 376, 826–834. [Google Scholar] [CrossRef]
- Levy, H.; Raby, B.A.; Lake, S.; Tantisira, K.G.; Kwiatkowski, D.; Lazarus, R.; Silverman, E.K.; Richter, B.; Klimecki, W.T.; Vercelli, D.; et al. Association of defensin beta-1 gene polymorphisms with asthma. J. Allergy Clin. Immunol. 2005, 115, 252–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchers, N.S.; Santos-Valente, E.; Toncheva, A.A.; Wehkamp, J.; Franke, A.; Gaertner, V.D.; Nordkild, P.; Genuneit, J.; Jensen, B.; Kabesch, M. Human β-Defensin 2 Mutations Are Associated With Asthma and Atopy in Children and Its Application Prevents Atopic Asthma in a Mouse Model. Front. Immunol. 2021, 12, 636061. [Google Scholar] [CrossRef]
- Pinkerton, J.W.; Kim, R.Y.; Koeninger, L.; Armbruster, N.S.; Hansbro, N.G.; Brown, A.C.; Jayaraman, R.; Shen, S.; Malek, N.; Cooper, M.A.; et al. Human β-defensin-2 suppresses key features of asthma in murine models of allergic airways disease. Clin. Exp. Allergy 2021, 51, 120–131. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peptide (Sequence) | Asthma Suppression | Asthma Promoting | Receptors | References |
|---|---|---|---|---|
| Neuropeptide Y (NPY) (YPSKPDNPGEDAPAEDLARYYSALRHYINLITRQRY) | Induction of AHR | [19,20] | ||
| ↑ AHR and airway inflammation | NPY-Y1 | [11,19] | ||
| Neurotensin (NT) (XLYENKPRRPYIL) | ↓ AHR and inflammation | NTSR1 | [10,26,27] | |
| Neuropeptide S (NPS) (SFRNGVGTGMKKTSFQRAKS) | ↓ AHR | [35] | ||
| Nociceptin (N)/Orphanin OFQ (FGGFTGARKSARKLANQ) | ↓ airway constriction ↓ inflammation | NOP1 | [40,43,44,46] | |
| Bombesin (BN) (XQRLGNQWAVGHLM) | ↑ neutrophilic inflammation ↑ cytokine production | GRPR | [53,55,56,57] | |
| ↑ contractile response | GRPR | [52] | ||
| ↑ mast cell proliferation and chemotaxis | [54] | |||
| ↑ airway injury and re-modeling | BRS-3 | [58,61] | ||
| Somatostatin (ST) (AGCKNFFWKTFTSC) | ↓ inflammation and AHR | sst4 | [71,72,73,74] |
| Peptide (Sequence) | Affinity [99] | Activation of NKA Receptors | |
|---|---|---|---|
| SP (RPKPQQFFGLM) | NK1 > NK2 >> NK3 | NK1 ↑ inflammatory cells recruitment [100,101] | NK1, NK2, NK3 ↑ airway smooth muscle constriction [102,103] |
| NKA (HKTDSFVGLM) | NK2 > NK 3> NK1 | ||
| NKB (DMHDFFVGLM) | NK3 > NK2 | ||
| HK1 (RSRTRQFYGLM) | NK1 >> NK2 >> NK3 | ||
| Endokinin (A/B GKASQFFGLM) | NK1 > NK2 >> NK3 | ||
| Endokinin C (KKAYQLEHTFQGLL) | NK3 | ||
| Endokinin D (VGAYQLEHTFQGLL) | NK3 | ||
| Peptide (Sequence) | Asthma Suppression | Asthma Promoting | Receptors | References | |
|---|---|---|---|---|---|
| Vasoactive intestinal polypeptide (VIP) (HSDAVFTDNYTRLRKQMAVKKYLNSILN) | Mice lacking the VIP gene exhibited spontaneous asthma features including AHR | [95] | |||
| Bronchodilatation | VAPC2 | [87,94] | |||
| TACHYKININS | Substance P (SP) (RPKPQQFFGLM) | ↑ mucus secretion ↑ development of AHR↑ recruitment of airwayneutrophils, ↑ degranulation of mast cells | NK1 | [100,101,119,122] | |
| Neurokinin A (NKA) (HKTDSFVGLM) | ↑ bronchoconstriction ↑ development of AHR | NK2 | [102,119,123] | ||
| HK-1 (RSRTRQFYGLM) | ↑degranulation of mast cells | NK-1, MRGPRX2 | [122] | ||
| Calcitonin gene–related peptide (CGRP) (ACDTATCVTHRLAGLLSRSGGVVKNNFVPTNVGSKAF) | ↑ vasodilatation, ↑ mucus secretion and edema ↑ trachea muscle contractions to capsaicin and electrical field | RAMP 1 | [5,128,129,130,131,132] | ||
| ↓ AHR and eosinophilic inflammation | [133] | ||||
| Peptide | Asthma Suppression | Asthma Promoting | Receptors | References |
|---|---|---|---|---|
| Adiponectin | Reduces airway inflammation | T-cadherin | [140,141,147] | |
| Leptin | Promotes airway inflammation and bronchoconstriction | Ob-R | [145,169,194] | |
| Resistin | Promotes airway inflammation and airway remodeling | Not known | [204,209,210] | |
| Ghrelin | Reduces airway inflammation and airway hyperreactivity | GHS-R | [225,226,227] |
| Peptide | Asthma Suppression | Asthma Promoting | References |
|---|---|---|---|
| LL-37 | Proinflammatory and exacerbates inflammation and AHR | [254,255,256] | |
| β-defensin 1 (hBD-1) | Promotes airway inflammation; chemoattractant for immune cells | [254,258] | |
| β-defensin 2 (hBD-2) | Prevents airway inflammation and AHR | [259,260] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczyńska, K.; Zając, D.; Wojciechowski, P.; Jampolska, M. Regulatory Peptides in Asthma. Int. J. Mol. Sci. 2021, 22, 13656. https://doi.org/10.3390/ijms222413656
Kaczyńska K, Zając D, Wojciechowski P, Jampolska M. Regulatory Peptides in Asthma. International Journal of Molecular Sciences. 2021; 22(24):13656. https://doi.org/10.3390/ijms222413656
Chicago/Turabian StyleKaczyńska, Katarzyna, Dominika Zając, Piotr Wojciechowski, and Monika Jampolska. 2021. "Regulatory Peptides in Asthma" International Journal of Molecular Sciences 22, no. 24: 13656. https://doi.org/10.3390/ijms222413656