
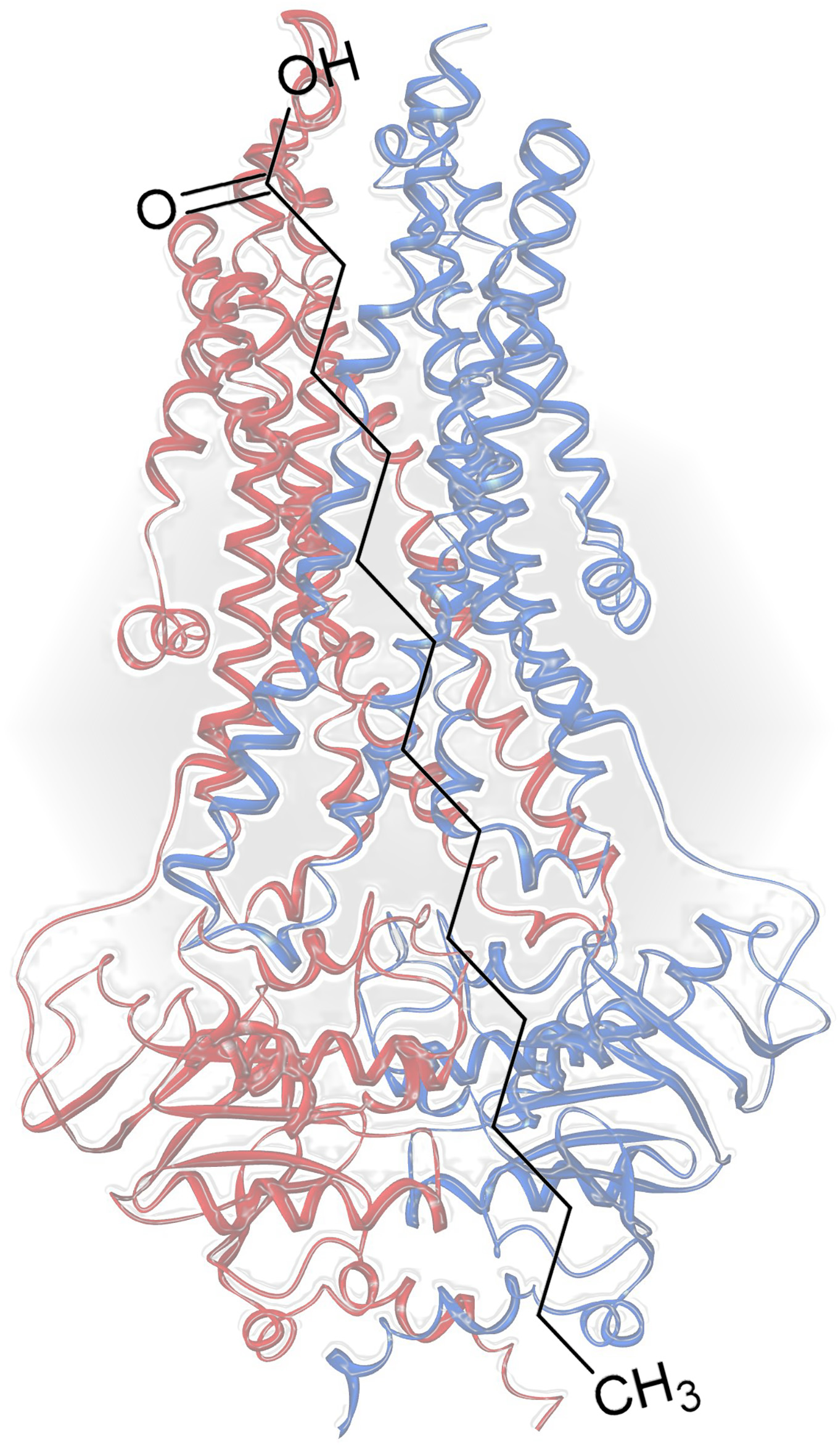
Myristic Acid Inhibits the Activity of the Bacterial ABC Transporter BmrA

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
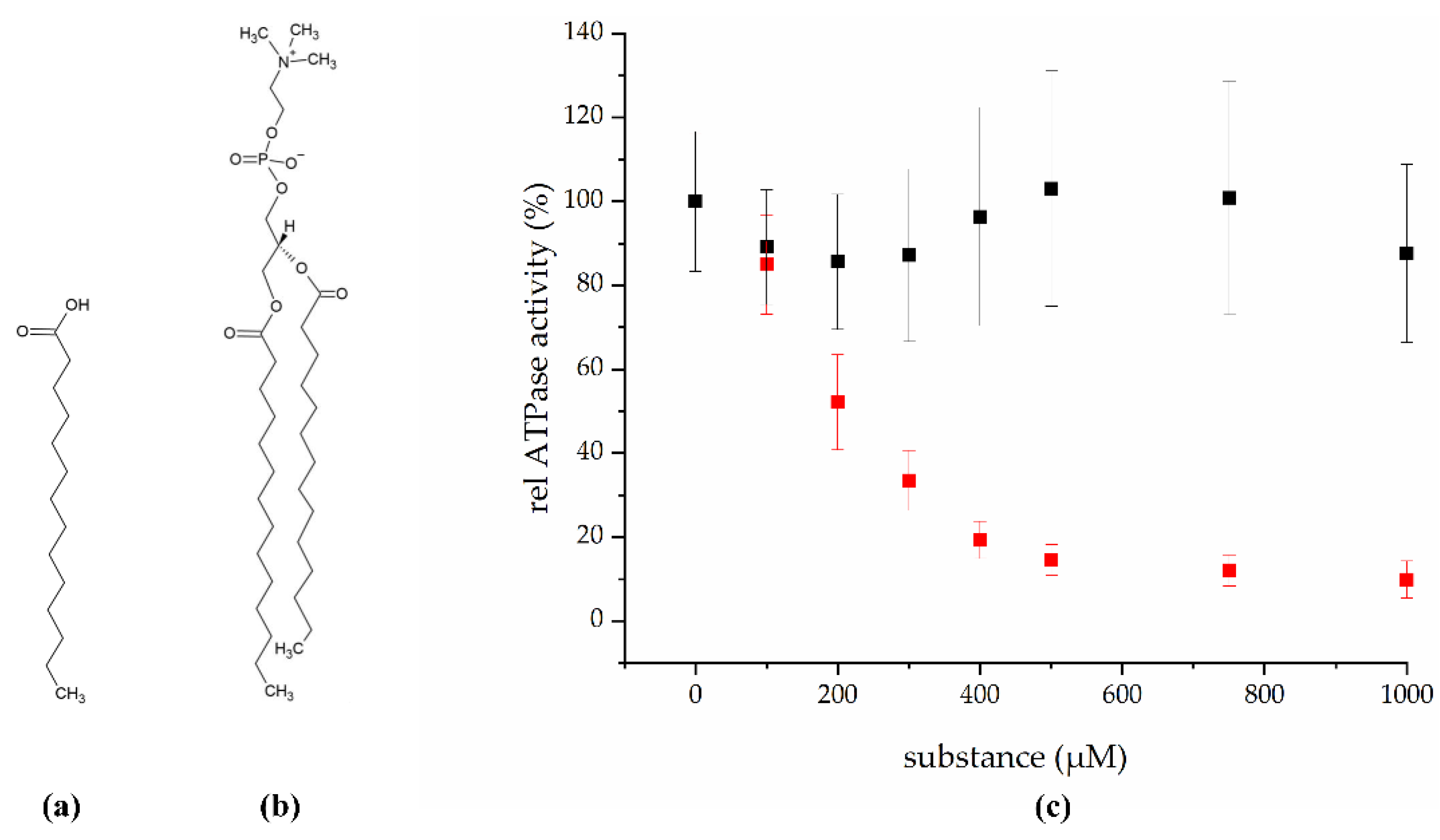
2.1. Myristic Acid Inhibits the ATPase Activity of the ABC Transporter BmrA
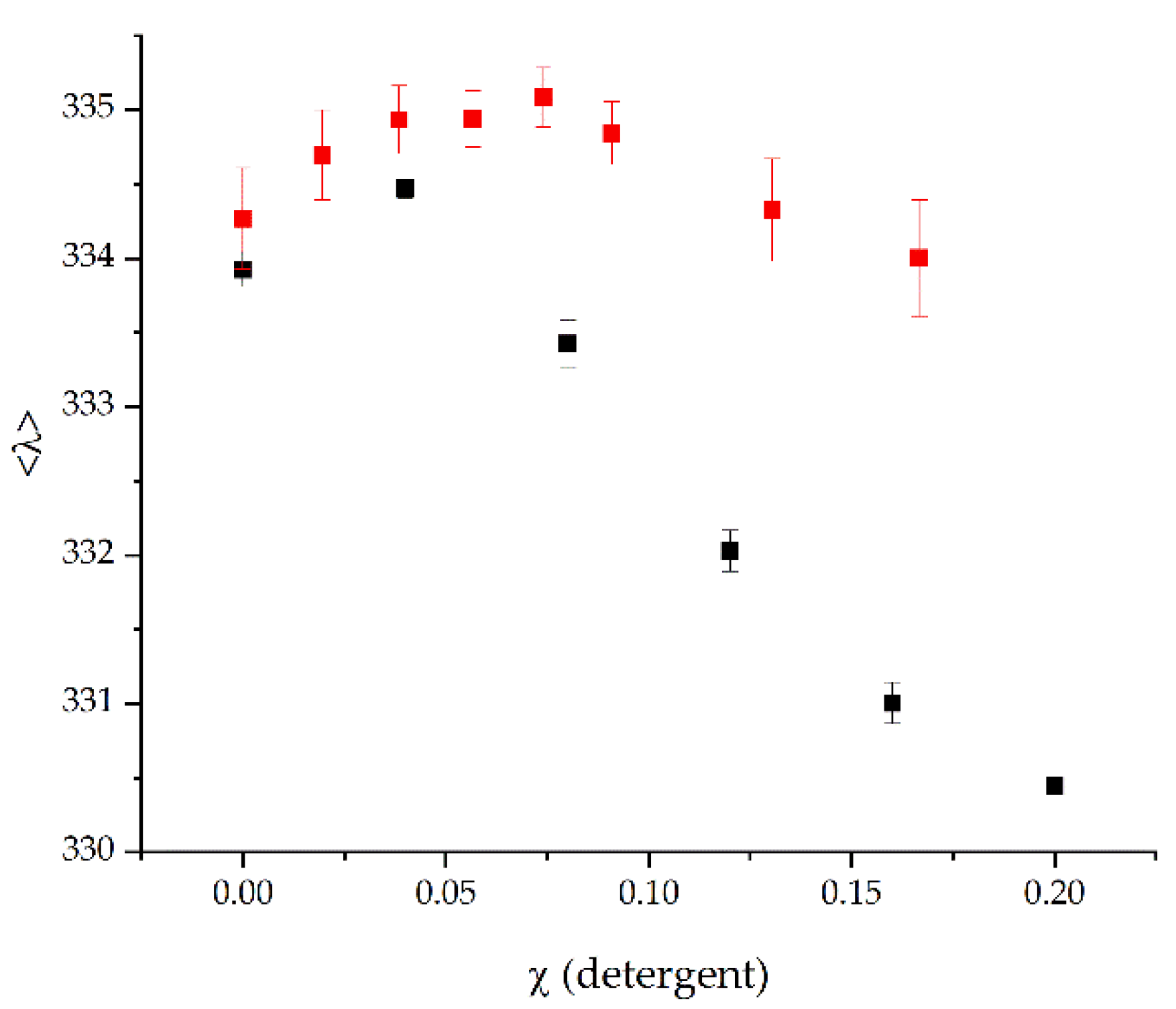
2.2. The Stability of BmrA in Micelles Is Not Affected by Myristic Acid
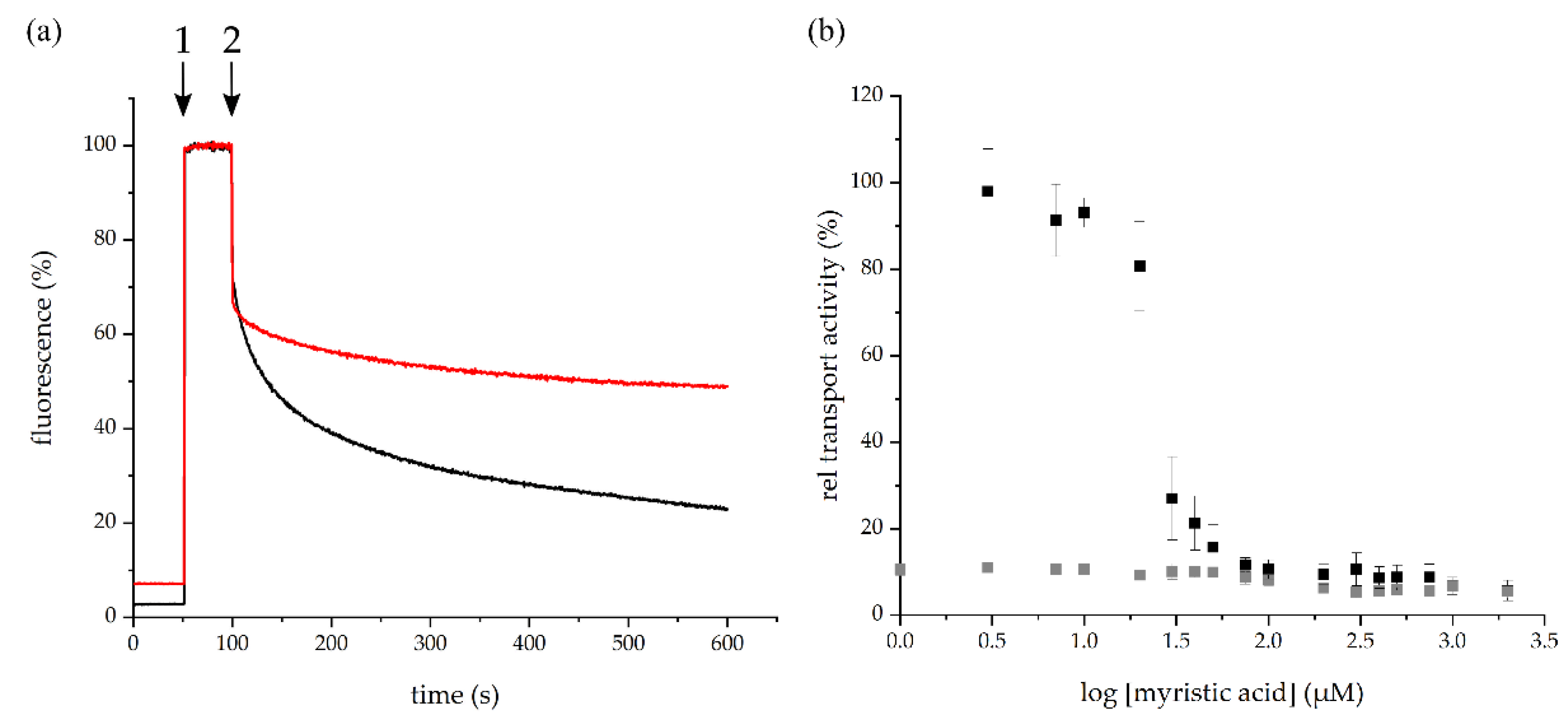
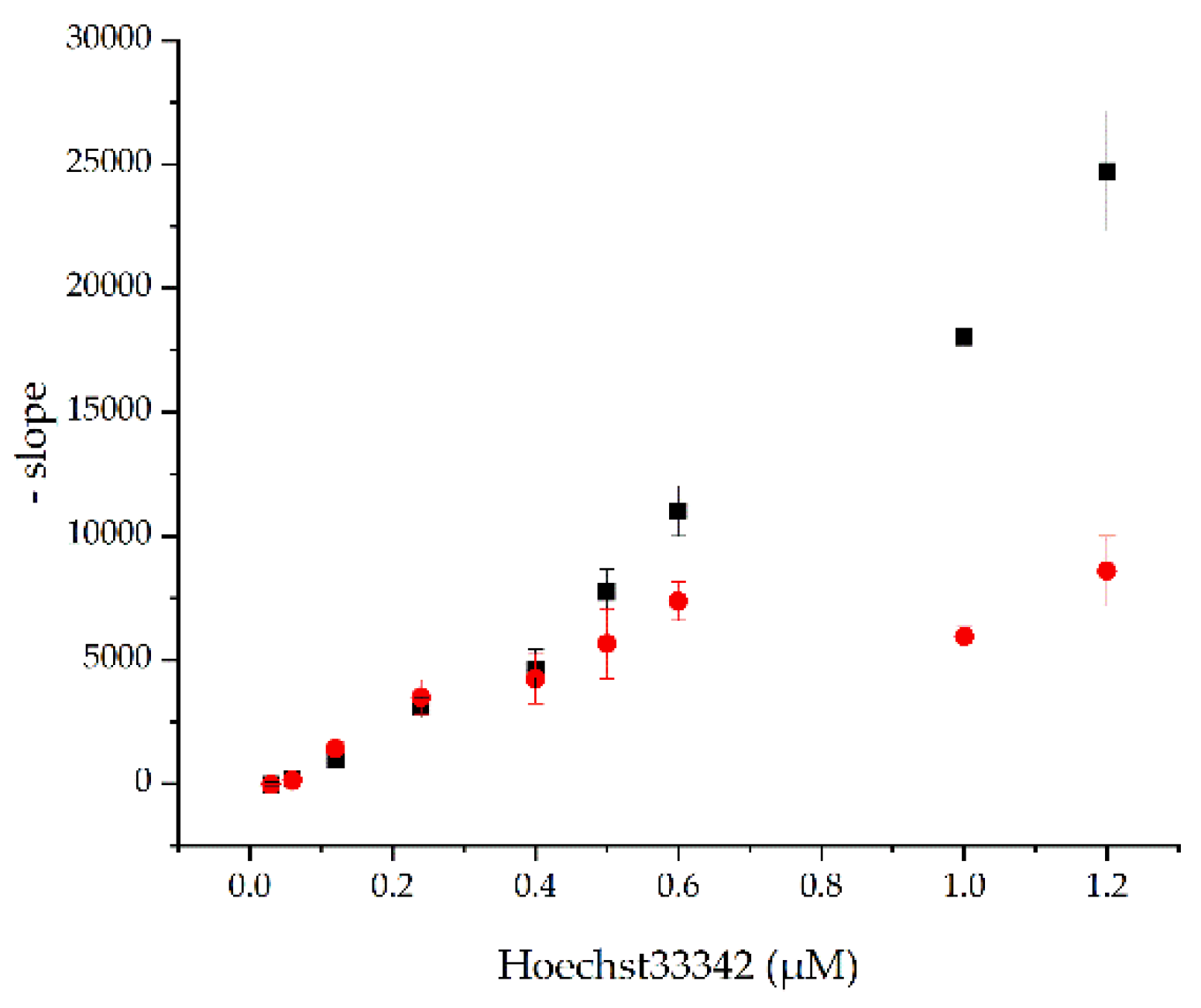
2.3. Myristic Acid Inhibits the BmrA-Mediated Transport of Hoechst 33342 in Inverted Membrane Vesicles
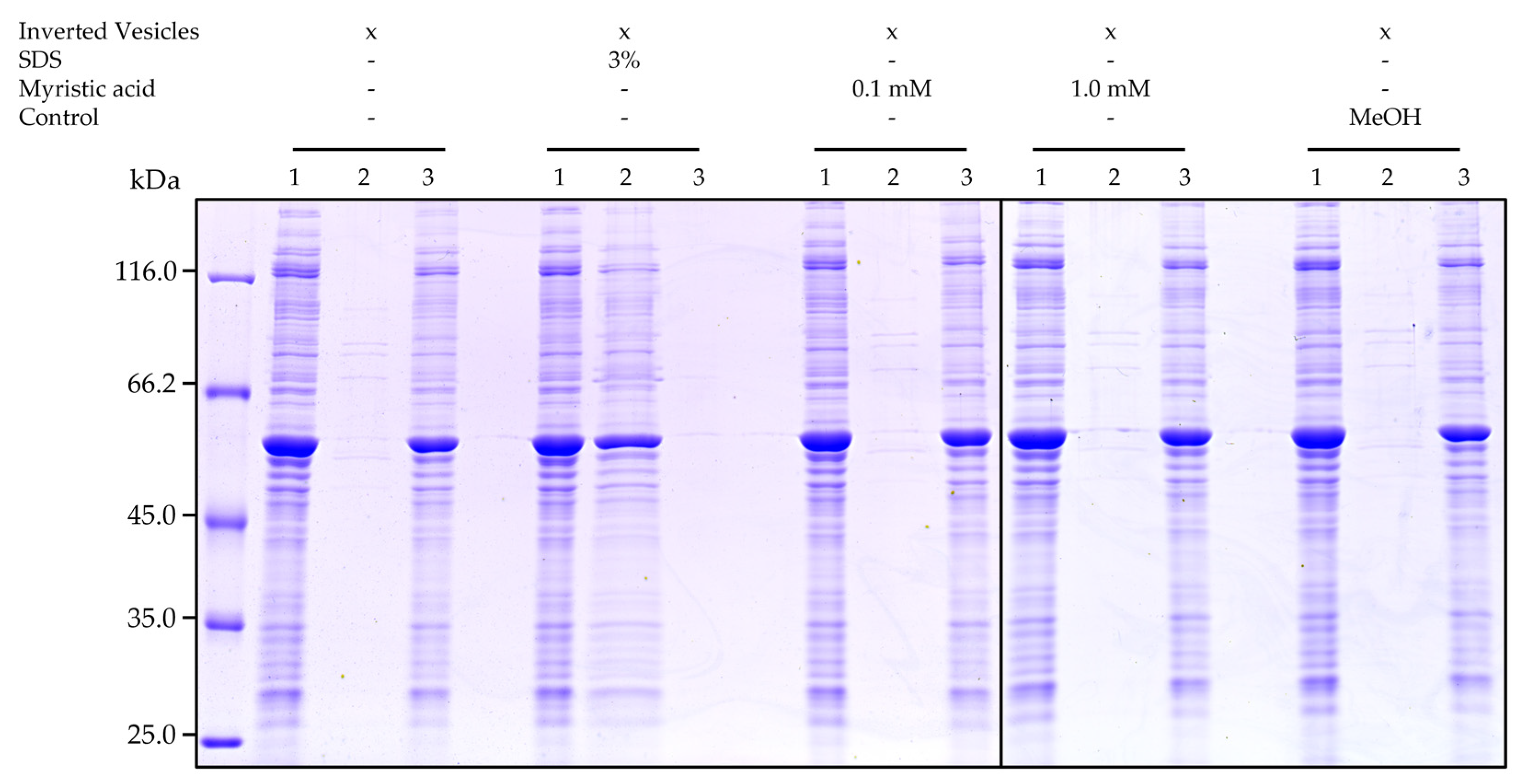
2.4. Myristic Acid Does Not Solubilize Overexpressed BmrA in Inverted Vesicles
3. Discussion
4. Materials and Methods
4.1. Screening for BmrA Inhibitors using Fungal Extracts and Isolation of Myristic Acid from IBWF 030-11
4.2. NMR Analysis
4.3. Cloning
4.4. Expression
4.5. Purification
4.6. ATPase Activity of Purified BmrA
4.7. Hoechst 33342 Transport Measured Using BmrA-Containing Inverted E. coli Membrane Vesicles
4.8. Destabilization of BmrA in DDM Micelles
4.9. Stability of BmrA in Inverted Vesicles
5. Conclusions and Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilkens, S. Structure and Mechanism of ABC Transporters. F1000Prime Rep. 2015, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hediger, M.A.; Romero, M.F.; Peng, J.-B.; Rolfs, A.; Takanaga, H.; Bruford, E.A. The ABCs of Solute Carriers: Physiological, Pathological and Therapeutic Implications of Human Membrane Transport Proteins. Pflugers Arch. Eur. J. Physiol. 2004, 447, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC Transporters: From Microorganisms to Man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Hanekop, N.; Zaitseva, J.; Jenewein, S.; Holland, I.B.; Schmitt, L. Molecular Insights into the Mechanism of ATP-Hydrolysis by the NBD of the ABC-Transporter HlyB. FEBS Lett. 2006, 580, 1036–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, I.B.; Blight, M.A. ABC-ATPases, Adaptable Energy Generators Fuelling Transmembrane Movement of a Variety of Molecules in Organisms from Bacteria to Humans. J. Mol. Biol. 1999, 293, 381–399. [Google Scholar] [CrossRef]
- Dalmas, O.; Do Cao, M.-A.; Lugo, M.R.; Sharom, F.J.; Di Pietro, A.; Jault, J.-M. Time-Resolved Fluorescence Resonance Energy Transfer Shows That the Bacterial Multidrug ABC Half-Transporter BmrA Functions as a Homodimer. Biochemistry 2005, 44, 4312–4321. [Google Scholar] [CrossRef]
- Falasca, M.; Linton, K.J. Investigational ABC Transporter Inhibitors. Expert Opin. Investig. Drugs 2012, 21, 657–666. [Google Scholar] [CrossRef]
- Smith, P.C.; Karpowich, N.; Millen, L.; Moody, J.E.; Rosen, J.; Thomas, P.J.; Hunt, J.F. ATP Binding to the Motor Domain from an ABC Transporter Drives Formation of a Nucleotide Sandwich Dimer. Mol. Cell 2002, 10, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Neyfakh, A.A.; Bidnenko, V.E.; Chen, L.B. Efflux-Mediated Multidrug Resistance in Bacillus Subtilis: Similarities and Dissimilarities with the Mammalian System. Proc. Natl. Acad. Sci. USA 1991, 88, 4781–4785. [Google Scholar] [CrossRef] [Green Version]
- Krügel, H.; Licht, A.; Biedermann, G.; Petzold, A.; Lassak, J.; Hupfer, Y.; Schlott, B.; Hertweck, C.; Platzer, M.; Brantl, S.; et al. Cervimycin C Resistance in Bacillus Subtilis Is Due to a Promoter Up-Mutation and Increased MRNA Stability of the Constitutive ABC-Transporter Gene BmrA. FEMS Microbiol. Lett. 2010, 313, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Steinfels, E.; Orelle, C.; Fantino, J.-R.; Dalmas, O.; Rigaud, J.-L.; Denizot, F.; Di Pietro, A.; Jault, J.-M. Characterization of YvcC (BmrA), a Multidrug ABC Transporter Constitutively Expressed in Bacillus Subtilis. Biochemistry 2004, 43, 7491–7502. [Google Scholar] [CrossRef]
- Chami, M.; Steinfels, E.; Orelle, C.; Jault, J.-M.; Di Pietro, A.; Rigaud, J.-L.; Marco, S. Three-Dimensional Structure by Cryo-Electron Microscopy of YvcC, an Homodimeric ATP-Binding Cassette Transporter from Bacillus Subtilis. J. Mol. Biol. 2002, 315, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Lacabanne, D.; Lends, A.; Danis, C.; Kunert, B.; Fogeron, M.L.; Jirasko, V.; Chuilon, C.; Lecoq, L.; Orelle, C.; Chaptal, V.; et al. Gradient Reconstitution of Membrane Proteins for Solid-State NMR Studies. J. Biomol. NMR 2017, 69, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, A.D. ACD/ChemSketch 1.0 (Freeware); ACD/ChemSketch 2.0 and Its Tautomers, Dictionary, and 3D Plug-Ins; ACD/HNMR 2.0; ACD/CNMR 2.0. J. Chem. Educ. 1997, 74, 905–906. [Google Scholar] [CrossRef] [Green Version]
- Ravaud, S.; Do Cao, M.A.; Jidenko, M.; Ebel, C.; Le Maire, M.; Jault, J.M.; Di Pietro, A.; Haser, R.; Aghajari, N. The ABC Transporter BmrA from Bacillus Subtilis Is a Functional Dimer When in a Detergent-Solubilized State. Biochem. J. 2006, 395, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orelle, C.; Gubellini, F.; Durand, A.; Marco, S.; Lévy, D.; Gros, P.; Di Pietro, A.; Jault, J.-M. Conformational Change Induced by ATP Binding in the Multidrug ATP-Binding Cassette Transporter BmrA. Biochemistry 2008, 47, 2404–2412. [Google Scholar] [CrossRef]
- Sharom, F.J. Characterization and Functional Reconstitution of the Multidrug Transporter. J. Bioenerg. Biomembr. 1995, 27, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, S.V.; Lelong, I.H.; Zhang, J.; Cardarelli, C.O.; Gottesman, M.M.; Pastan, I. Partial Purification and Reconstitution of the Human Multidrug-Resistance Pump: Characterization of the Drug-Stimulatable ATP Hydrolysis. Proc. Natl. Acad. Sci. USA 1992, 89, 8472–8476. [Google Scholar] [CrossRef] [Green Version]
- de Athayde Moncorvo Collado, A.; Corbalán, N.; Homolya, L.; Morero, R.; Minahk, C. Resveratrol Modulates ATPase Activity of Liposome-Reconstituted ABCG1. FEBS Lett. 2013, 587, 2359–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fameau, A.-L.; Ventureira, J.; Novales, B.; Douliez, J.-P. Foaming and Emulsifying Properties of Fatty Acids Neutralized by Tetrabutylammonium Hydroxide. Colloids Surfaces A Physicochem. Eng. Asp. 2012, 403, 87–95. [Google Scholar] [CrossRef]
- Veerappan, A.; Cymer, F.; Klein, N.; Schneider, D. The Tetrameric α-Helical Membrane Protein GlpF Unfolds via a Dimeric Folding Intermediate. Biochemistry 2011, 50, 10223–10230. [Google Scholar] [CrossRef]
- Anbazhagan, V.; Cymer, F.; Schneider, D. Unfolding a Transmembrane Helix Dimer: A FRET Study in Mixed Micelles. Arch. Biochem. Biophys. 2010, 495, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, P.; Mogensen, J.E.; Otzen, D.E. Using Micellar Mole Fractions to Assess Membrane Protein Stability in Mixed Micelles. Biochim. Biophys. Acta-Biomembr. 2005, 1716, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellmann, N.; Schneider, D. A Complex Unfolding Pathway of α-Helical Membrane Proteins in SDS-Containing Micelles. Biophys. J. 2021, 120, 3857–3859. [Google Scholar] [CrossRef] [PubMed]
- Otzen, D.E.; Nedergaard Pedersen, J.; Kumar Somavarapu, A.; Clement, A.; Ji, M.; Hartvig Petersen, E.; Skov Pedersen, J.; Urban, S.; Schafer, N.P. Cys-Labeling Kinetics of Membrane Protein GlpG: A Role for Specific SDS Binding and Micelle Changes? Biophys. J. 2021, 120, 4115–4128. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.B.; Ling, V. Reconstitution of Drug Transport by Purified P-Glycoprotein. J. Biol. Chem. 1995, 270, 16167–16175. [Google Scholar] [CrossRef] [Green Version]
- Burdock, G.A.; Carabin, I.G. Safety Assessment of Myristic Acid as a Food Ingredient. Food Chem. Toxicol. 2007, 45, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.G.; Rutberg, L.; Samuelsson, B. The Chemical Composition of the Cytoplasmic Membrane of Bacillus Subtilis. Eur. J Biochem 1967, 2, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Doige, C.A.; Yu, X.; Sharom, F.J. The Effects of Lipids and Detergents on ATPase-Active P-Glycoprotein. Biochim. Biophys. Acta 1993, 1146, 65–72. [Google Scholar] [CrossRef]
- Sharom, F.J. The P-Glycoprotein Efflux Pump: How Does It Transport Drugs? J. Membr. Biol. 1997, 160, 161–175. [Google Scholar] [CrossRef]
- Neumann, J.; Rose-Sperling, D.; Hellmich, U.A. Diverse Relations between ABC Transporters and Lipids: An Overview. Biochim. Biophys. Acta-Biomembr. 2017, 1859, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Janvilisri, T.; Venter, H.; Shahi, S.; Balakrishnan, L.; van Veen, H.W. The ATP Binding Cassette Multidrug Transporter LmrA and Lipid Transporter MsbA Have Overlapping Substrate Specificities. J. Biol. Chem. 2003, 278, 35193–35198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woebking, B.; Reuter, G.; Shilling, R.A.; Velamakanni, S.; Shahi, S.; Venter, H.; Balakrishnan, L.; van Veen, H.W. Drug-Lipid A Interactions on the. J. Bacteriol. 2005, 187, 6363–6369. [Google Scholar] [CrossRef] [Green Version]
- Lee, A. Lipid–Protein Interactions in Biological Membranes: A Structural Perspective. Biochim. Biophys. Acta-Biomembr. 2003, 1612, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Lacabanne, D.; Orelle, C.; Lecoq, L.; Kunert, B.; Chuilon, C.; Wiegand, T.; Ravaud, S.; Jault, J.M.; Meier, B.H.; Böckmann, A. Flexible-to-Rigid Transition Is Central for Substrate Transport in the ABC Transporter BmrA from Bacillus Subtilis. Commun. Biol. 2019, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, H.E.; Kotlyar, V.; Nudelman, A. NRM Chemicals Shifts of Common Laboratory Solvents as Traces Imputities. J. Org. Chem. 1997, 62, 7512–7515. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.T.; Matton, P.; Fairhurst, N.W.G.; John, M.P.; Carbery, D.R. Biomimetic Flavin-Catalyzed Aldehyde Oxidation. Org. Lett. 2012, 14, 3656–3659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinfels, E.; Orelle, C.; Dalmas, O.; Penin, F.; Miroux, B.; Di Pietro, A.; Jault, J.-M. Highly Efficient Over-Production in E. Coli of YvcC, a Multidrug-like ATP-Binding Cassette Transporter from Bacillus Subtilis. Biochim. Biophys. Acta-Biomembr. 2002, 1565, 1–5. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oepen, K.; Özbek, H.; Schüffler, A.; Liermann, J.C.; Thines, E.; Schneider, D. Myristic Acid Inhibits the Activity of the Bacterial ABC Transporter BmrA. Int. J. Mol. Sci. 2021, 22, 13565. https://doi.org/10.3390/ijms222413565
Oepen K, Özbek H, Schüffler A, Liermann JC, Thines E, Schneider D. Myristic Acid Inhibits the Activity of the Bacterial ABC Transporter BmrA. International Journal of Molecular Sciences. 2021; 22(24):13565. https://doi.org/10.3390/ijms222413565
Chicago/Turabian StyleOepen, Kristin, Hüseyin Özbek, Anja Schüffler, Johannes C. Liermann, Eckhard Thines, and Dirk Schneider. 2021. "Myristic Acid Inhibits the Activity of the Bacterial ABC Transporter BmrA" International Journal of Molecular Sciences 22, no. 24: 13565. https://doi.org/10.3390/ijms222413565