
Emerging Role of TCA Cycle-Related Enzymes in Human Diseases


Abstract
:1. Introduction
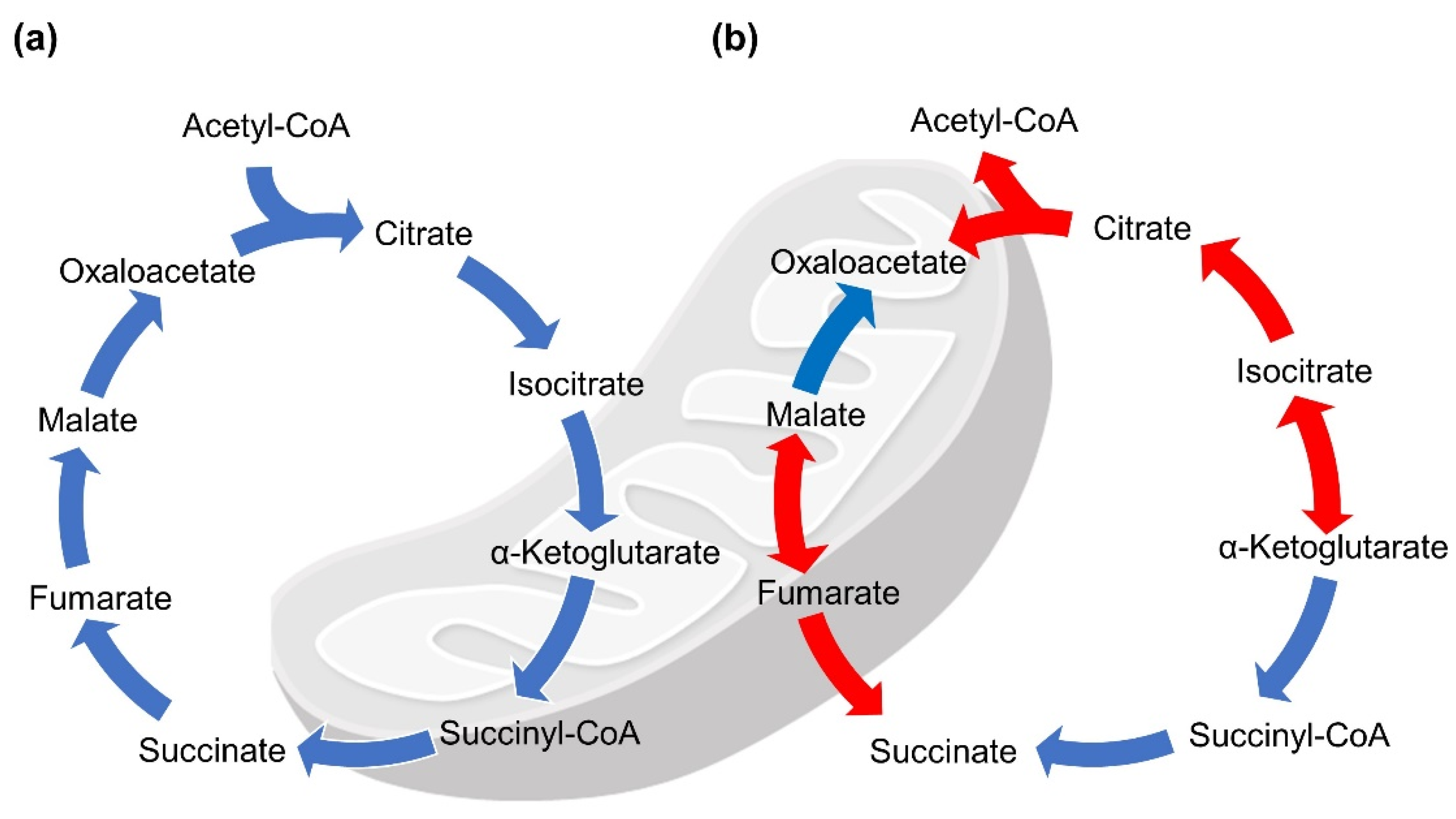
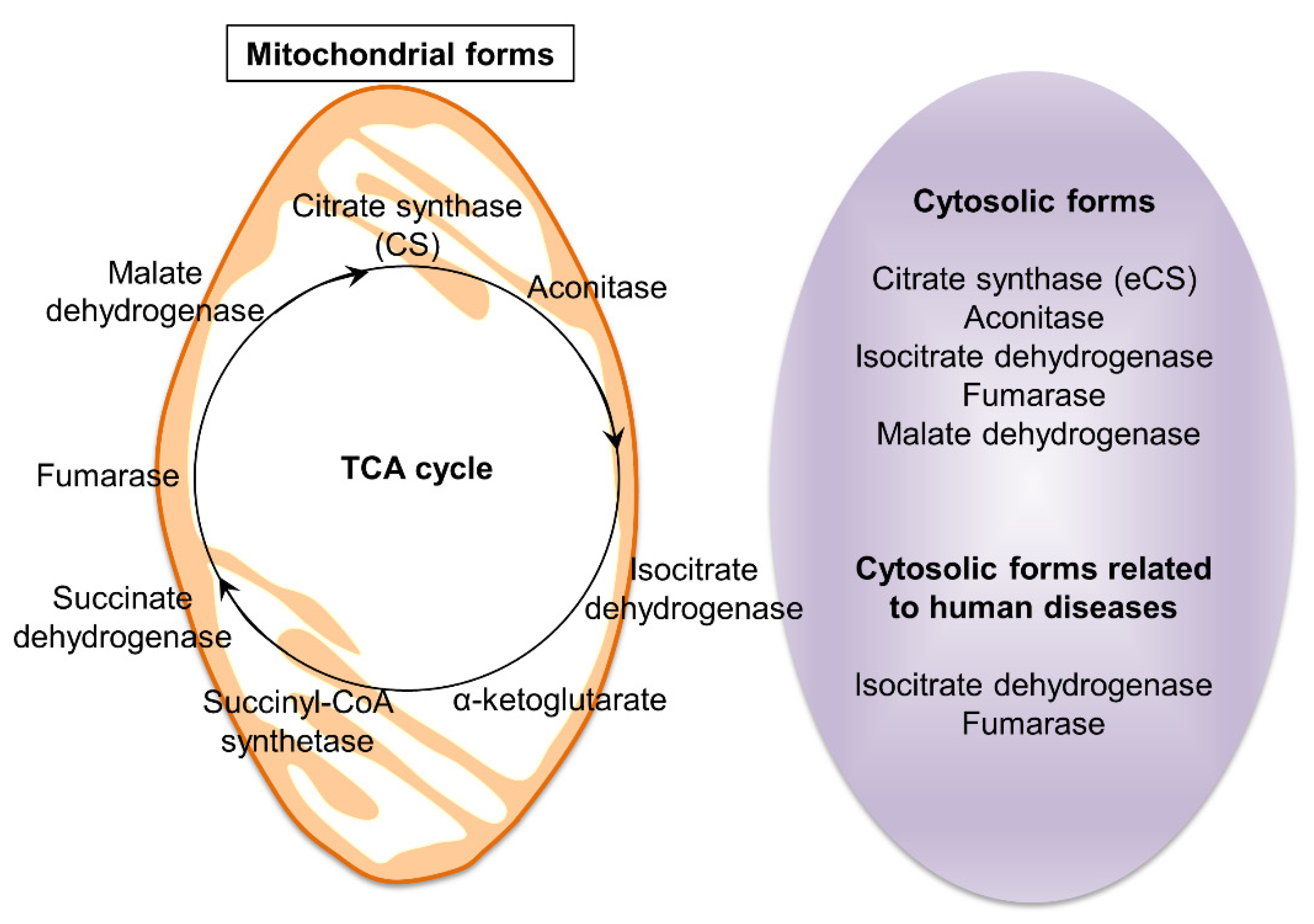
2. TCA-Related Enzymes and Diseases Arising from Their Dysfunction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Abbreviation | Localization | Diseases | References |
|---|---|---|---|---|
| Citrate synthase | CS | Mitochondria | Cell death (in a human cell line, in vitro) | [17] |
| Citrate lyase | ACLY | Cytosol | Atherosclerotic plaques (in mice, in vivo) | [19] |
| Extra-mitochondrial citrate synthase | eCS | Cytosol | Decrease in age-dependent male fertility (in mice, in vivo) | [33] |
| Aconitase | ACO1 | Cytosol | Encephalopathy (in humans, in vivo) | [18,21,22] |
| ACO2 | Mitochondria | Optic atrophy (in humans, in vivo) | ||
| Isocitrate dehydrogenase | IDH1 | Cytosol | Gliomas, acute myeloid leukemia (in humans, in vivo) | [6,18,23] |
| IDH2 | Mitochondria | |||
| Succinate dehydrogenase | SDHA | Mitochondria | Paragangliomas (in humans, in vivo) | [18,26] |
| SDHB | Gastrointestinal stromal tumors, paragangliomas, renal cell carcinoma, T-cell acute leukemia (in humans, in vivo) | [18,25,27,28,29] | ||
| SDHC | Gastrointestinal stromal tumors, paragangliomas (in humans, in vivo) | [18,25,29] | ||
| SDHD | Gastrointestinal stromal tumors, paragangliomas (in humans, in vivo) | [18,25,29] | ||
| Fumarase (fumarase hydratase) | FH | Mitochondria | Encephalopathy, leiomyomas, leiomyomatosis, renal cell cancer, ovary cystadenomas, breast cancer (in humans, in vivo) | [18,31] |
| Cytosol | ||||
| α-ketoglutarate dehydrogenase | OGDH | Mitochondria | Neurological disorder (in humans, in vivo) | [34] |
| Malate dehydrogenase | MDH1 | Cytosol | Encephalopathy (in a human cell line, in vitro) | [35,36] |
| MDH2 | Mitochondria | |||
| Malic enzyme | ME1 | Cytosol | Unknown | [37,38] |
| ME2 | Mitochondria | Idiopathic generalized epilepsy (in humans, in vivo) | [37,39] | |
| ME3 | Mitochondria | Unknown | [37] | |
| Glutamate-oxaloacetate transaminase | GOT1 | Cytosol | Unknown | [40] |
| GOT2 | Mitochondria | Neurometabolic disorder (in humans, in vivo) | [41] |
3. Ca2+ Signaling and Mitochondria
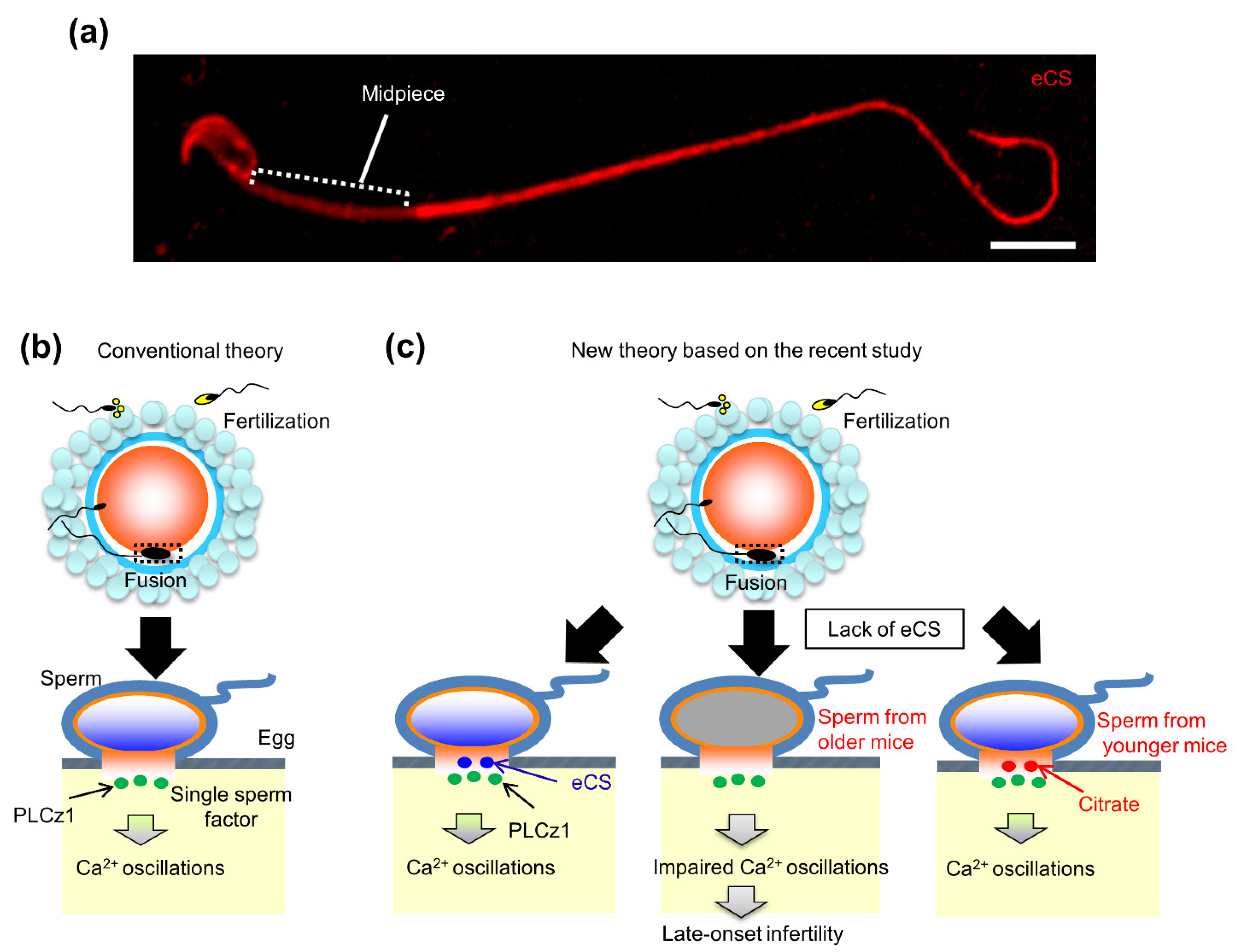
4. Age-Dependent eCS Function
5. Predicted Existence of Extra-Mitochondrial TCA (eTCA) Cycle
6. Clinical Trials as Metabolic Therapies
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martínez-Reyes, I.; Diebold, L.P.; Kong, H.; Schieber, M.; Huang, H.; Hensley, C.T.; Mehta, M.M.; Wang, T.; Santos, J.H.; Woychik, R. TCA cycle and mitochondrial membrane potential are necessary for diverse biological functions. Mol. Cell 2016, 61, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Chan, D.C. Fusion and fission: Interlinked processes critical for mitochondrial health. Annu. Rev. Genet. 2012, 46, 265–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqui, T.; Farooqui, A.A. Aging: An important factor for the pathogenesis of neurodegenerative diseases. Mech. Ageing Dev. 2009, 130, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Bruni, F. Mitochondria: From physiology to pathology. Life 2021, 11, 991. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachtershauser, G. Evolution of the first metabolic cycles. Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Muchowska, K.B.; Varma, S.J.; Chevallot-Beroux, E.; Lethuillier-Karl, L.; Li, G.; Moran, J. Metals promote sequences of the reverse Krebs cycle. Nat. Ecol. Evol. 2017, 1, 1716–1721. [Google Scholar] [CrossRef]
- Keller, M.A.; Kampjut, D.; Harrison, S.A.; Ralser, M. Sulfate radicals enable a non-enzymatic Krebs cycle precursor. Nat. Ecol. Evol. 2017, 1, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffens, L.; Pettinato, E.; Steiner, T.M.; Mall, A.; Konig, S.; Eisenreich, W.; Berg, I.A. High CO2 levels drive the TCA cycle backwards towards autotrophy. Nature 2021, 592, 784–788. [Google Scholar] [CrossRef]
- Tang, K.H.; Blankenship, R.E. Both forward and reverse TCA cycles operate in green sulfur bacteria. J. Biol. Chem. 2010, 285, 35848–35854. [Google Scholar] [CrossRef] [Green Version]
- Nunoura, T.; Chikaraishi, Y.; Izaki, R.; Suwa, T.; Sato, T.; Harada, T.; Mori, K.; Kato, Y.; Miyazaki, M.; Shimamura, S.; et al. A primordial and reversible TCA cycle in a facultatively chemolithoautotrophic thermophile. Science 2018, 359, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Bodovitz, S. Single cell analysis: The new frontier in ‘omics’. Trends Biotechnol. 2010, 28, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstraete, K.; Verschueren, K.H.; Dansercoer, A.; Savvides, S.N. Acetyl-CoA is produced by the citrate synthase homology module of ATP-citrate lyase. Nat. Struct. Mol. Biol. 2021, 28, 636–638. [Google Scholar] [CrossRef]
- Cai, Q.; Zhao, M.; Liu, X.; Wang, X.; Nie, Y.; Li, P.; Liu, T.; Ge, R.; Han, F. Reduced expression of citrate synthase leads to excessive superoxide formation and cell apoptosis. Biochem. Biophys. Res. Commun. 2017, 485, 388–394. [Google Scholar] [CrossRef]
- Raimundo, N.; Baysal, B.E.; Shadel, G.S. Revisiting the TCA cycle: Signaling to tumor formation. Trends Mol. Med. 2011, 17, 641–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baardman, J.; Verberk, S.G.; Van der Velden, S.; Gijbels, M.J.; van Roomen, C.P.; Sluimer, J.C.; Broos, J.Y.; Griffith, G.R.; Prange, K.H.; van Weeghel, M. Macrophage ATP citrate lyase deficiency stabilizes atherosclerotic plaques. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Khodagholi, F.; Shaerzadeh, F.; Montazeri, F. Mitochondrial aconitase in neurodegenerative disorders: Role of a metabolism-related molecule in neurodegeneration. Curr. Drug Targets 2018, 19, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Abela, L.; Spiegel, R.; Crowther, L.M.; Klein, A.; Steindl, K.; Papuc, S.M.; Joset, P.; Zehavi, Y.; Rauch, A.; Plecko, B. Plasma metabolomics reveals a diagnostic metabolic fingerprint for mitochondrial aconitase (ACO2) deficiency. PLoS ONE 2017, 12, e0176363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Cai, G.H.; Xia, B.; Wang, X.; Zhang, C.C.; Xie, B.C.; Shi, X.C.; Liu, H.; Lu, J.F.; Zhang, R.X. Mitochondrial aconitase controls adipogenesis through mediation of cellular ATP production. FASEB J. 2020, 34, 6688–6702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uckermann, O.; Juratli, T.A.; Galli, R.; Conde, M.; Wiedemuth, R.; Krex, D.; Geiger, K.; Temme, A.; Schackert, G.; Koch, E.; et al. Optical analysis of glioma: Fourier-transform infrared spectroscopy reveals the IDH1 mutation status. Clin. Cancer Res. 2018, 24, 2530–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekstra, A.S.; Bayley, J.-P. The role of complex II in disease. Biochim. Biophys. Acta (BBA)-Bioenerg. 2013, 1827, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Astuti, D.; Hart-Holden, N.; Latif, F.; Lalloo, F.; Black, G.C.; Lim, C.; Moran, A.; Grossman, A.B.; Hodgson, S.V.; Freemont, A. Genetic analysis of mitochondrial complex II subunits SDHD, SDHB and SDHC in paraganglioma and phaeochromocytoma susceptibility. Clin. Endocrinol. 2003, 59, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Tufton, N.; Ghelani, R.; Srirangalingam, U.; Kumar, A.V.; Drake, W.M.; Iacovazzo, D.; Skordilis, K.; Berney, D.; Khoo, B.; Akker, S.A. SDHA mutated paragangliomas may be at high risk of metastasis. Endocr. Relat. Cancer 2017, 24, L43–L49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanharanta, S.; Buchta, M.; McWhinney, S.R.; Virta, S.K.; Peçzkowska, M.; Morrison, C.D.; Lehtonen, R.; Januszewicz, A.; Järvinen, H.; Juhola, M. Early-onset renal cell carcinoma as a novel extraparaganglial component of SDHB-associated heritable paraganglioma. Am. J. Hum. Genet. 2004, 74, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baysal, B.E. A recurrent stop-codon mutation in succinate dehydrogenase subunit B gene in normal peripheral blood and childhood T-cell acute leukemia. PLoS ONE 2007, 2, e436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratakis, C.; Carney, J. The triad of paragangliomas, gastric stromal tumours and pulmonary chondromas (Carney triad), and the dyad of paragangliomas and gastric stromal sarcomas (Carney–Stratakis syndrome): Molecular genetics and clinical implications. J. Intern. Med. 2009, 266, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffler, I.E. Mitochondria; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Crooks, D.R.; Maio, N.; Lang, M.; Ricketts, C.J.; Vocke, C.D.; Gurram, S.; Turan, S.; Kim, Y.Y.; Cawthon, G.M.; Sohelian, F.; et al. Mitochondrial DNA alterations underlie an irreversible shift to aerobic glycolysis in fumarate hydratase-deficient renal cancer. Sci Signal. 2021, 14, eabc4436. [Google Scholar] [CrossRef] [PubMed]
- Raimundo, N.; Ahtinen, J.; Fumić, K.; Barić, I.; Remes, A.M.; Renkonen, R.; Lapatto, R.; Suomalainen, A. Differential metabolic consequences of fumarate hydratase and respiratory chain defects. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2008, 1782, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Harada, Y.; Yamatoya, K.; Kawano, N.; Kanai, S.; Miyamoto, Y.; Nakamura, A.; Miyado, M.; Hayashi, Y.; Kuroki, Y.; et al. Extra-mitochondrial citrate synthase initiates calcium oscillation and suppresses age-dependent sperm dysfunction. Lab. Investig. 2020, 100, 583–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.E.; Park, L.C.; Sheu, K.-F.R.; Blass, J.P.; Calingasan, N.Y. The α-ketoglutarate dehydrogenase complex in neurodegeneration. Neurochem. Int. 2000, 36, 97–112. [Google Scholar] [CrossRef]
- Broeks, M.H.; Shamseldin, H.E.; Alhashem, A.; Hashem, M.; Abdulwahab, F.; Alshedi, T.; Alobaid, I.; Zwartkruis, F.; Westland, D.; Fuchs, S.; et al. MDH1 deficiency is a metabolic disorder of the malate-aspartate shuttle associated with early onset severe encephalopathy. Hum. Genet. 2019, 138, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Ait-El-Mkadem, S.; Dayem-Quere, M.; Gusic, M.; Chaussenot, A.; Bannwarth, S.; François, B.; Genin, E.C.; Fragaki, K.; Volker-Touw, C.L.; Vasnier, C. Mutations in MDH2, encoding a Krebs cycle enzyme, cause early-onset severe encephalopathy. Am. J. Hum. Genet. 2017, 100, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassinen, I.E. Signaling and regulation through the NAD+ and NADP+ networks. Antioxid. Redox Signal. 2019, 30, 857–874. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-X.; Ju, H.-Q.; Liu, Z.-X.; Chen, D.-L.; Wang, Y.; Zhao, Q.; Wu, Q.-N.; Zeng, Z.-l.; Qiu, H.-B.; Hu, P.-S. ME1 regulates NADPH homeostasis to promote gastric cancer growth and metastasis. Cancer Res. 2018, 78, 1972–1985. [Google Scholar] [CrossRef] [Green Version]
- Lenzen, K.P.; Heils, A.; Lorenz, S.; Hempelmann, A.; Sander, T. Association analysis of malic enzyme 2 gene polymorphisms with idiopathic generalized epilepsy. Epilepsia 2005, 46, 1637–1641. [Google Scholar] [CrossRef]
- Kremer, D.M.; Nelson, B.S.; Lin, L.; Yarosz, E.L.; Halbrook, C.J.; Kerk, S.A.; Sajjakulnukit, P.; Myers, A.; Thurston, G.; Hou, S.W. GOT1 inhibition promotes pancreatic cancer cell death by ferroptosis. Nat. Commun. 2021, 12, 1–13. [Google Scholar]
- Ramos, R.J.; van Karnebeek, C.D.; Ciapaite, J.; Pras-Raves, M.; Waterham, H.R.; Wanders, R.J.; Jans, J.J.; Verhoeven-Duif, N.M. Metabolic Consequences of GOT2 Deficiency. New Insight into Vitamin B6 Metabolism and Related Diseases. Ph.D. Thesis, Utrecht University, Utrecht, The Netherlands, 2019. [Google Scholar]
- Murai, S.; Ando, A.; Ebara, S.; Hirayama, M.; Satomi, Y.; Hara, T. Inhibition of malic enzyme 1 disrupts cellular metabolism and leads to vulnerability in cancer cells in glucose-restricted conditions. Oncogenesis 2017, 6, e329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellis, A.-T.; Misko, A.L.; Arjune, S.; Liang, Y.; Erdélyi, K.; Ditrói, T.; Kaczmarek, A.T.; Nagy, P.; Schwarz, G. The role of glutamate oxaloacetate transaminases in sulfite biosynthesis and H2S metabolism. Redox Biol. 2021, 38, 101800. [Google Scholar] [CrossRef]
- Santulli, G.; Nakashima, R.; Yuan, Q.; Marks, A.R. Intracellular calcium release channels: An update. J. Physiol. 2017, 595, 3041–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J.W. Calcium oscillations. Cold Spring Harb. Perspect. Biol. 2011, 3, a004226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feridooni, H.A.; Dibb, K.M.; Howlett, S.E. How cardiomyocyte excitation, calcium release and contraction become altered with age. J. Mol. Cell Cardiol. 2015, 83, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Bezprozvanny, I. Calcium signaling and neurodegenerative diseases. Trends Mol. Med. 2009, 15, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, J.; Josts, I.; Heidemann, J.; Mertens, H.D.; Maric, S.; Moulin, M.; Haertlein, M.; Busch, S.; Forsyth, V.T.; Svergun, D.I.; et al. Structural basis for activation of plasma-membrane Ca2+-ATPase by calmodulin. Commun. Biol. 2018, 1, 206. [Google Scholar] [CrossRef]
- Juhaszova, M.; Church, P.; Blaustein, M.P.; Stanley, E.F. Location of calcium transporters at presynaptic terminals. Eur. J. Neurosci. 2000, 12, 839–846. [Google Scholar] [CrossRef]
- Gherardi, G.; Monticelli, H.; Rizzuto, R.; Mammucari, C. The mitochondrial Ca2+ uptake and the fine-tuning of aerobic metabolism. Front. Physiol. 2020, 11, 554904. [Google Scholar] [CrossRef] [PubMed]
- Boyman, L.; Mikhasenko, H.; Hiller, R.; Khananshvili, D. Kinetic and equilibrium properties of regulatory calcium sensors of NCX1 protein. J. Biol. Chem. 2009, 284, 6185–6193. [Google Scholar] [CrossRef] [Green Version]
- Chovancova, B.; Liskova, V.; Babula, P.; Krizanova, O. Role of sodium/calcium exchangers in tumors. Biomolecules 2020, 10, 1257. [Google Scholar] [CrossRef] [PubMed]
- Lock, J.T.; Parker, I. IP(3) mediated global Ca2+ signals arise through two temporally and spatially distinct modes of Ca2+ release. eLife 2020, 9, e55008. [Google Scholar] [CrossRef]
- Roos, J.; DiGregorio, P.J.; Yeromin, A.V.; Ohlsen, K.; Lioudyno, M.; Zhang, S.; Safrina, O.; Kozak, J.A.; Wagner, S.L.; Cahalan, M.D.; et al. STIM1, an essential and conserved component of store-operated Ca2+ channel function. J. Cell Biol. 2005, 169, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.S. The molecular choreography of a store-operated calcium channel. Nature 2007, 446, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Fahrner, M.; Schindl, R.; Muik, M.; Derler, I.; Romanin, C. The STIM-Orai Pathway: The Interactions Between STIM and Orai. Adv. Exp. Med. Biol. 2017, 993, 59–81. [Google Scholar]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ sensing: Its role in calcium homeostasis and signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, R.; Zhang, X. Ca2+ Regulation of TRP ion channels. Int. J. Mol. Sci. 2018, 19, 1256. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, E.J.; Rutter, G.A. Mitochondrial calcium as a key regulator of mitochondrial ATP production in mammalian cells. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 1324–1333. [Google Scholar] [CrossRef] [Green Version]
- Creamer, T.P. Calcineurin. Cell Commun. Signal. 2020, 18, 137. [Google Scholar] [CrossRef]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.L.; Stein, P.; Jefferson, W.N.; Padilla-Banks, E.; Williams, C.J. Calcium influx-mediated signaling is required for complete mouse egg activation. Proc. Natl. Acad. Sci. USA 2012, 109, 4169–4174. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.R.; Yang, W.X. Calcium influx and sperm-evoked calcium responses during oocyte maturation and egg activation. Oncotarget 2017, 8, 89375–89390. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Kwon, W.S.; Pang, M.G. Calcium influx and male fertility in the context of the sperm proteome: An update. Biomed. Res. Int. 2014, 2014, 841615. [Google Scholar] [CrossRef]
- Swann, K.; Lai, F.A. Egg activation at fertilization by a soluble sperm protein. Physiol. Rev. 2016, 96, 127–149. [Google Scholar] [CrossRef] [Green Version]
- Hogben, M.; Parrington, J.; Shevchenko, V.; Swann, K.; Lai, F.A. Calcium oscillations, sperm factors and egg activation at fertilisation. J. Mol. Med. 1998, 76, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Parrington, J.; Swann, K.; Shevchenko, V.I.; Sesay, A.K.; Lai, F.A. Calcium oscillations in mammalian eggs triggered by a soluble sperm protein. Nature 1996, 379, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Churchill, G.C.; O’Neill, J.S.; Masgrau, R.; Patel, S.; Thomas, J.M.; Genazzani, A.A.; Galione, A. Sperm deliver a new second messenger: NAADP. Curr. Biol. 2003, 13, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Whalley, T.; McDougall, A.; Crossley, I.; Swann, K.; Whitaker, M. Internal calcium release and activation of sea urchin eggs by cGMP are independent of the phosphoinositide signaling pathway. Mol. Biol. Cell 1992, 3, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Kuo, R.C.; Baxter, G.T.; Thompson, S.H.; Stricker, S.A.; Patton, C.; Bonaventura, J.; Epel, D. NO is necessary and sufficient for egg activation at fertilization. Nature 2000, 406, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Galione, A.; McDougall, A.; Busa, W.B.; Willmott, N.; Gillot, I.; Whitaker, M. Redundant mechanisms of calcium-induced calcium release underlying calcium waves during fertilization of sea urchin eggs. Science 1993, 261, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.R.; Swann, K. Molecular triggers of egg activation at fertilization in mammals. Reproduction 2016, 152, R41–R50. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.T.; Sutovsky, P.; Manandhar, G.; Xu, W.; Katayama, M.; Day, B.N.; Park, K.W.; Yi, Y.J.; Xi, Y.W.; Prather, R.S.; et al. PAWP, a sperm-specific WW domain-binding protein, promotes meiotic resumption and pronuclear development during fertilization. J. Biol. Chem. 2007, 282, 12164–12175. [Google Scholar] [CrossRef] [Green Version]
- Zafar, M.I.; Lu, S.; Li, H. Sperm-oocyte interplay: An overview of spermatozoon’s role in oocyte activation and current perspectives in diagnosis and fertility treatment. Cell Biosci. 2021, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Hachem, A.; Godwin, J.; Ruas, M.; Lee, H.C.; Ferrer Buitrago, M.; Ardestani, G.; Bassett, A.; Fox, S.; Navarrete, F.; de Sutter, P.; et al. PLCζ is the physiological trigger of the Ca2+ oscillations that induce embryogenesis in mammals but conception can occur in its absence. Development 2017, 144, 2914–2924. [Google Scholar]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef] [PubMed]
- Aras-Tosun, D.; Cakar, Z.; Can, A.; Ozkavukcu, S.; Kaplanoglu, I.; Cinar, O. Phospholipase C-zeta levels are not correlated with fertilisation rates in infertile couples. Andrologia 2021, e14269. [Google Scholar] [CrossRef]
- Harada, Y.; Matsumoto, T.; Hirahara, S.; Nakashima, A.; Ueno, S.; Oda, S.; Miyazaki, S.; Iwao, Y. Characterization of a sperm factor for egg activation at fertilization of the newt Cynops pyrrhogaster. Dev. Biol. 2007, 306, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Yamatoya, K.; Miyado, K.; Miyado, M.; Miyamoto, Y. Neuronal expression of Ca2+ oscillation initiator is linked to rapid neonatal growth in mice. MicroPubl. Biol. 2020, 2020. [Google Scholar] [CrossRef]
- Sokolov, A.A.; Miall, R.C.; Ivry, R.B. The cerebellum: Adaptive prediction for movement and cognition. Trends Cogn. Sci. 2017, 21, 313–332. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.; Scheffler, P.; Lohr, C. Dopamine-induced calcium signaling in olfactory bulb astrocytes. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cutando, L.; Puighermanal, E.; Castell, L.; Tarot, P.; Belle, M.; Bertaso, F.; Arango-Lievano, M.; Ango, F.; Rubinstein, M.; Chédotal, A. Cerebellar dopamine D2 receptors regulate preference for social novelty. bioRxiv 2019. [Google Scholar] [CrossRef]
- Mehdizadeh, M.; Ashtari, N.; Jiao, X.; Rahimi Balaei, M.; Marzban, A.; Qiyami-Hour, F.; Kong, J.; Ghavami, S.; Marzban, H. Alteration of the dopamine receptors’ expression in the cerebellum of the lysosomal acid phosphatase 2 mutant (naked-ataxia (NAX)) mouse. Int. J. Mol. Sci. 2020, 21, 2914. [Google Scholar] [CrossRef] [PubMed]
- Kight, C.E.; Fleming, S.E. Oxidation of glucose carbon entering the TCA cycle is reduced by glutamine in small intestine epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 1995, 268, G879–G888. [Google Scholar] [CrossRef] [PubMed]
- Yarian, C.S.; Toroser, D.; Sohal, R.S. Aconitase is the main functional target of aging in the citric acid cycle of kidney mitochondria from mice. Mech. Ageing Dev. 2006, 127, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moller, A. Ejaculate quality, testes size and sperm production in mammals. Funct. Ecol. 1989, 3, 91–96. [Google Scholar] [CrossRef]
- Cardaci, S.; Ciriolo, M.R. TCA cycle defects and cancer: When metabolism tunes redox state. Int. J. Cell Biol. 2012, 2012, 161837. [Google Scholar] [CrossRef]
- Wachnowsky, C.; Hendricks, A.L.; Wesley, N.A.; Ferguson, C.; Fidai, I.; Cowan, J.A. Understanding the mechanism of [4Fe-4S] cluster assembly on eukaryotic mitochondrial and cytosolic aconitase. Inorg. Chem. 2019, 58, 13686–13695. [Google Scholar] [CrossRef]
- Johnson, N.B.; Deck, K.M.; Nizzi, C.P.; Eisenstein, R.S. A synergistic role of IRP1 and FBXL5 proteins in coordinating iron metabolism during cell proliferation. J. Biol. Chem. 2017, 292, 15976–15989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leshets, M.; Silas, Y.B.H.; Lehming, N.; Pines, O. Fumarase: From the TCA cycle to DNA damage response and tumor suppression. Front. Mol. Biosci. 2018, 5, 68. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Starinina, E.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of fumarase in germinating maize seeds. Physiol. Plant. 2014, 152, 231–240. [Google Scholar] [CrossRef]
- Dyson, B.C.; Miller, M.A.; Feil, R.; Rattray, N.; Bowsher, C.G.; Goodacre, R.; Lunn, J.E.; Johnson, G.N. FUM2, a cytosolic fumarase, is essential for acclimation to low temperature in Arabidopsis thaliana. Plant. Physiol. 2016, 172, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Himpsl, S.D.; Shea, A.E.; Zora, J.; Stocki, J.A.; Foreman, D.; Alteri, C.J.; Mobley, H.L.T. The oxidative fumarase FumC is a key contributor for E. coli fitness under iron-limitation and during UTI. PLoS Pathog. 2020, 16, e1008382. [Google Scholar] [CrossRef] [Green Version]
- Yogev, O.; Yogev, O.; Singer, E.; Shaulian, E.; Goldberg, M.; Fox, T.D.; Pines, O. Fumarase: A mitochondrial metabolic enzyme and a cytosolic/nuclear component of the DNA damage response. PLoS Biol. 2010, 8, e1000328. [Google Scholar] [CrossRef] [Green Version]
- Van Lith, S.A.; Navis, A.C.; Lenting, K.; Verrijp, K.; Schepens, J.T.; Hendriks, W.J.; Schubert, N.A.; Venselaar, H.; Wevers, R.A.; van Rooij, A.; et al. Identification of a novel inactivating mutation in Isocitrate Dehydrogenase 1 (IDH1-R314C) in a high grade astrocytoma. Sci. Rep. 2016, 6, 30486. [Google Scholar] [CrossRef] [Green Version]
- Meemongkolkiat, T.; Allison, J.; Seebacher, F.; Lim, J.; Chanchao, C.; Oldroyd, B.P. Thermal adaptation in the honeybee (Apis mellifera) via changes to the structure of malate dehydrogenase. J. Exp. Biol. 2020, 223, jeb228239. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. The warburg effect: Why and how do cancer cells activate glycolysis in the presence of oxygen? Anti Cancer Agents Med. Chem. Former. Curr. Med. Chem. Anti Cancer Agents 2008, 8, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Panfoli, I.; Calzia, D.; Ravera, S.; Bruschi, M.; Tacchetti, C.; Candiani, S.; Morelli, A.; Candiano, G. Extramitochondrial tricarboxylic acid cycle in retinal rod outer segments. Biochimie 2011, 93, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Ari, C.; Pilla, R.; D’Agostino, D. Nutritional/metabolic therapies in animal models of amyotrophic lateral sclerosis, Alzheimer’s disease, and seizures. In Bioactive Nutraceuticals and Dietary Supplements in Neurological and Brain Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 449–459. [Google Scholar]
- Luengo, A.; Gui, D.Y.; Vander Heiden, M.G. Targeting metabolism for cancer therapy. Cell Chem. Biol. 2017, 24, 1161–1180. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, P.; Yarani, R.; Dokaneheifard, S.; Mansouri, K. The emerging role of targeting cancer metabolism for cancer therapy. Tumor Biol. 2020, 42, 1010428320965284. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fu, L.; Li, Y.; Wang, W.; Gong, M.; Zhang, J.; Dong, X.; Huang, J.; Wang, Q.; Mackay, C.R. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8+ T cell immunity. Cell Metab. 2021, 33, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Descamps, H.C.; Herrmann, B.; Wiredu, D.; Thaiss, C.A. The path toward using microbial metabolites as therapies. EBioMedicine 2019, 44, 747–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, W.; Suzuki, M.; Saito, T.; Miyado, K. Emerging Role of TCA Cycle-Related Enzymes in Human Diseases. Int. J. Mol. Sci. 2021, 22, 13057. https://doi.org/10.3390/ijms222313057
Kang W, Suzuki M, Saito T, Miyado K. Emerging Role of TCA Cycle-Related Enzymes in Human Diseases. International Journal of Molecular Sciences. 2021; 22(23):13057. https://doi.org/10.3390/ijms222313057
Chicago/Turabian StyleKang, Woojin, Miki Suzuki, Takako Saito, and Kenji Miyado. 2021. "Emerging Role of TCA Cycle-Related Enzymes in Human Diseases" International Journal of Molecular Sciences 22, no. 23: 13057. https://doi.org/10.3390/ijms222313057