
Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
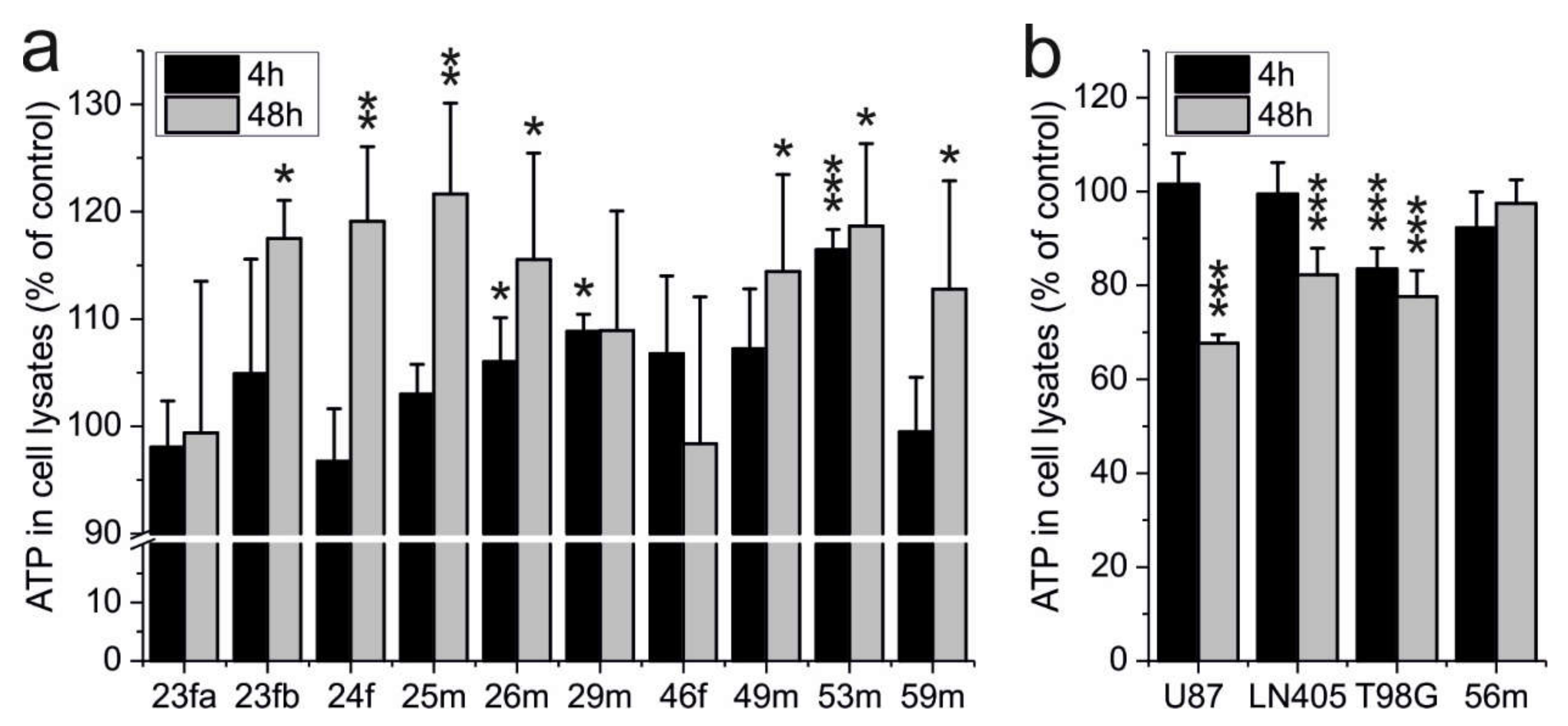
2.1. ATP Production of Erythrocytes Is Not Impaired by Carnosine
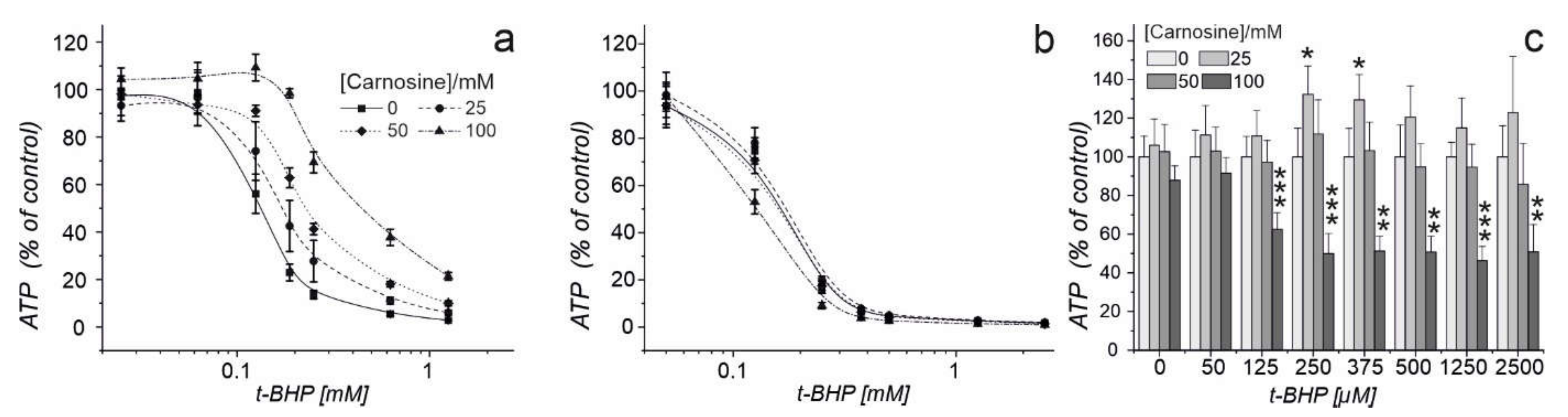
2.2. Carnosine Present in Medium Protects Erythrocytes against Oxidative Stress and Does Not Impair Their Defense against Radical Oxygen Species
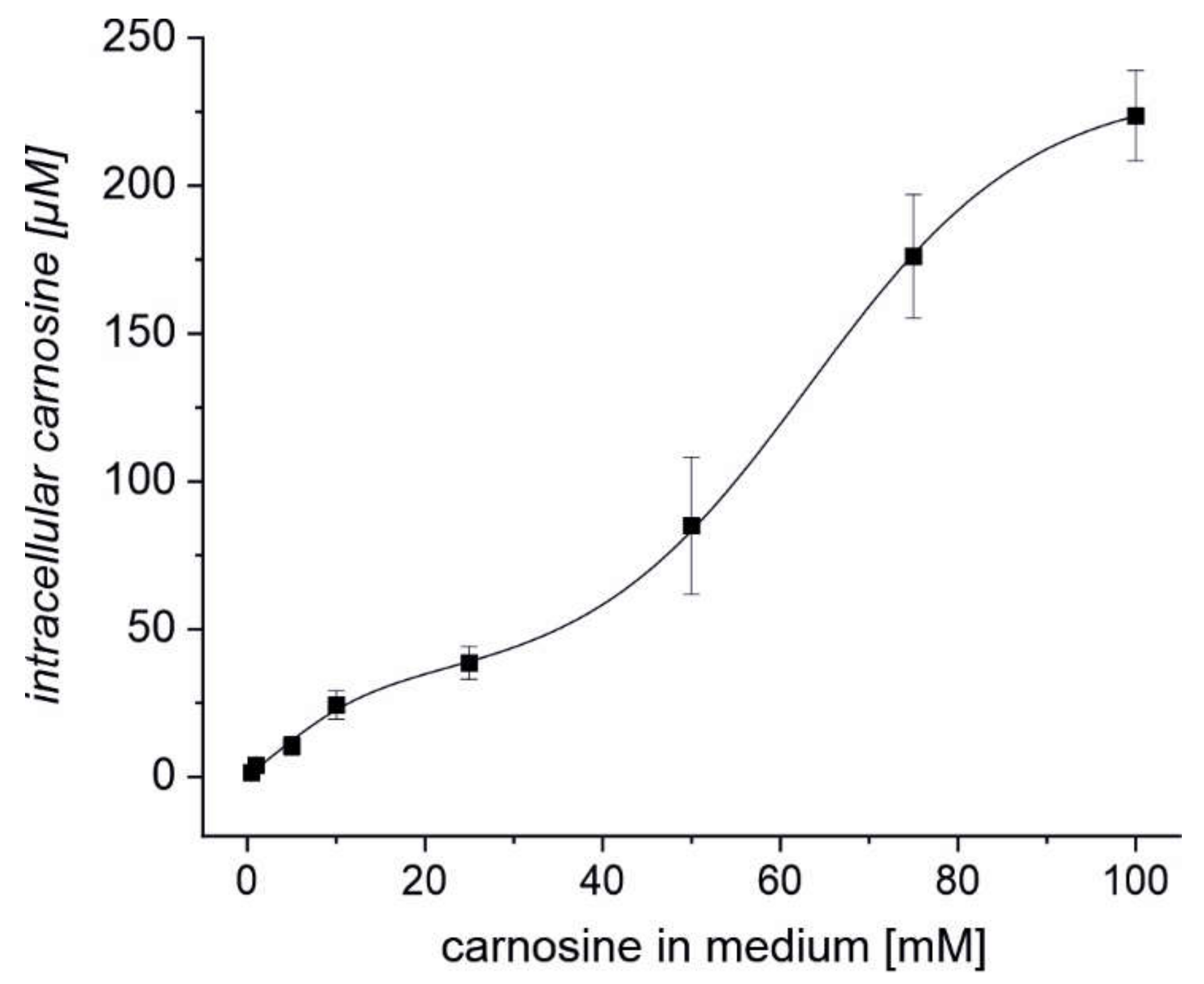
2.3. Erythrocytes Can Take up Extracellular Carnosine without Exhibiting Saturation up to a Concentration 100 mM
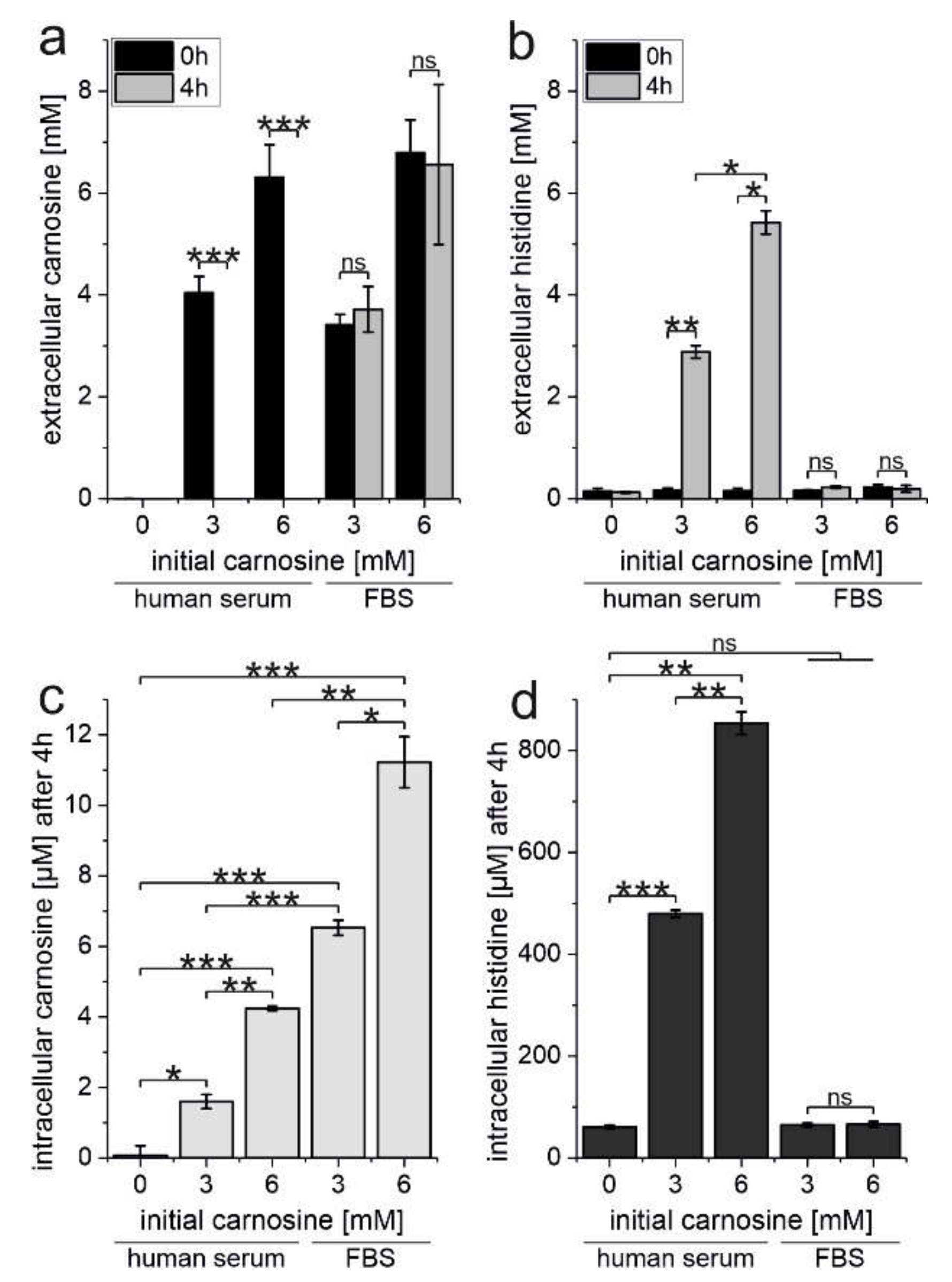
2.4. Erythrocytes Protect Carnosine from Degradation by Human Serum Carnosinase
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Isolation and Preparation of Erythrocytes
4.4. Determination of ATP in Cell Lysates
4.5. Pre-Treatment of Erythrocytes
4.6. Determination of Intracellular l-histidine and Carnosine
4.7. LC-MS Conditions for Carnosine and l-histidine Analysis
4.8. Quantification of the Intracellular Concentration of Carnosine in Red Blood Cells
4.9. Determination of Carnosinase Activity
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Informant Consent
References
- Gulewitsch, W.; Amiradzibi, S. Ueber das Carnosin, eine neue organische Base des Fleischextraktes. Ber. Dtsch. Chem. Ges. 1900, 33, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- Mannion, A.F.; Jakeman, P.M.; Dunnett, M.; Harris, R.C.; Willan, P.L. Carnosine and anserine concentrations in the quadriceps femoris muscle of healthy humans. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 64, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.C. The buffering of muscle in rigor; protein, phosphate and carnosine. J. Physiol. 1938, 92, 336–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, D.; Pisano, J.J.; Udenfriend, S. The distribution of homocarnosine in mammals. Arch. Biochem. Biophys. 1962, 99, 210–213. [Google Scholar] [CrossRef]
- Kohen, R.; Yamamoto, Y.; Cundy, K.C.; Ames, B.N. Antioxidant activity of carnosine, homocarnosine, and anserine present in muscle and brain. Proc. Natl. Acad. Sci. USA 1988, 85, 3175–3179. [Google Scholar] [CrossRef] [Green Version]
- Saunders, B.; Elliott-Sale, K.; Artioli, G.G.; Swinton, P.A.; Dolan, E.; Roschel, H.; Sale, C.; Gualano, B. β-alanine supplementation to improve exercise capacity and performance: A systematic review and meta-analysis. Br. J. Sports Med. 2017, 51, 658–669. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Baye, E.; de Courten, B. Carnosine and the processes of ageing. Maturitas 2016, 93, 28–33. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, K.; Zhang, W.; Zhang, G.; Zhou, M. Protective and Anti-Aging Effects of 5 Cosmeceutical Peptide Mixtures on Hydrogen Peroxide-Induced Premature Senescence in Human Skin Fibroblasts. Skin Pharmacol. Physiol. 2021, 34, 194–202. [Google Scholar] [CrossRef]
- Wang, S.; Niu, H.; Liu, Y.; Tan, Y.; Gao, H.; Ren, S.; Wang, L. Clinical Efficacy and Safety of Non-Cross-Linked Hyaluronic Acid Combined with L-carnosine for Horizontal Neck Wrinkles Treatment. Aesthetic Plast. Surg. 2021. Available online: https://doi.org/10.1007/s00266-021-02307-2 (accessed on 26 November 2011).
- Hipkiss, A.R.; Michaelis, J.; Syrris, P.; Kumar, S.; Lam, Y. Carnosine protects proteins against in vitro glycation and cross-linking. Biochem. Soc. Trans. 1994, 22, 399S. [Google Scholar] [CrossRef] [Green Version]
- Hipkiss, A.R.; Chana, H. Carnosine protects proteins against methylglyoxal-mediated modifications. Biochem. Biophys. Res. Commun. 1998, 248, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R.; Preston, J.E.; Himsworth, D.T.; Worthington, V.C.; Keown, M.; Michaelis, J.; Lawrence, J.; Mateen, A.; Allende, L.; Eagles, P.A.; et al. Pluripotent protective effects of carnosine, a naturally occurring dipeptide. Ann. N. Y. Acad. Sci. 1998, 854, 37–53. [Google Scholar]
- Banerjee, S.; Mukherjee, B.; Poddar, M.K.; Dunbar, G.L. Carnosine improves aging-induced cognitive impairment and brain regional neurodegeneration in relation to the neuropathological alterations in the secondary structure of amyloid beta (Aβ). J. Neurochem. 2021, 158, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The Therapeutic Potential of Carnosine/Anserine Supplementation against Cognitive Decline: A Systematic Review with Meta-Analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef]
- Boldyrev, A.; Fedorova, T.; Stepanova, M.; Dobrotvorskaya, I.; Kozlova, E.; Boldanova, N.; Bagyeva, G.; Ivanova-Smolenskaya, I.; Illarioshkin, S. Carnosine [corrected] Increases Efficiency of DOPA Therapy of Parkinson’s Disease: A Pilot Study. Rejuvenation Res. 2008, 11, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Ding, S.; Cheng, J.; Liu, Y.; Wang, B.; Xu, H.; Shen, Y.; Lyu, J. Carnosine Inhibits the Proliferation of Human Cervical Gland Carcinoma Cells Through Inhibiting Both Mitochondrial Bioenergetics and Glycolysis Pathways and Retarding Cell Cycle Progression. Integr. Cancer Ther. 2018, 17, 80–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, C.-H.; Hu, M.-L. L-Carnosine Inhibits Metastasis of SK-Hep-1 Cells by Inhibition of Matrix Metaoproteinase-9 Expression and Induction of an Antimetastatic Gene, nm23-H1. Nutr. Cancer 2008, 60, 526–533. [Google Scholar] [CrossRef]
- Lee, J.; Park, J.-R.; Lee, H.; Jang, S.; Ryu, S.-M.; Kim, H.; Kim, D.; Jang, A.; Yang, S.-R. L-carnosine induces apoptosis/cell cycle arrest via suppression of NF-κB/STAT1 pathway in HCT116 colorectal cancer cells. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 505–512. [Google Scholar] [CrossRef]
- Renner, C.; Zemitzsch, N.; Fuchs, B.; Geiger, K.D.; Hermes, M.; Hengstler, J.; Gebhardt, R.; Meixensberger, J.; Gaunitz, F. Carnosine retards tumor growth in vivo in an NIH3T3-HER2/neu mouse model. Mol. Cancer 2010, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yang, J.; Li, J.; Shi, X.; Ouyang, L.; Tian, Y.; Lu, J. Carnosine inhibits the proliferation of human gastric cancer SGC-7901 cells through both of the mitochondrial respiration and glycolysis pathways. PLoS ONE 2014, 9, e104632. [Google Scholar] [CrossRef] [Green Version]
- Gaunitz, F.; Hipkiss, A.R. Carnosine and cancer: A perspective. Amino Acids 2012, 43, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkiss, A.R.; Gaunitz, F. Inhibition of tumour cell growth by carnosine: Some possible mechanisms. Amino Acids 2014, 46, 327–337. [Google Scholar] [CrossRef]
- Sadikali, F.; Darwish, R.; Watson, W.C. Carnosinase activity of human gastrointestinal mucosa. Gut 1975, 16, 585–589. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.C.; Kucera, C.M.; Lenney, J.F. Purification and Properties of Human Serum Carnosinase. Clin. Chim. Acta 1991, 196, 193–205. [Google Scholar] [CrossRef]
- Gilardoni, E.; Gervasoni, S.; Maspero, M.; Dallanoce, C.; Vistoli, G.; Carini, M.; Aldini, G.; Regazzoni, L. Development of a direct LC-ESI-MS method for the measurement of human serum carnosinase activity. J. Pharm. Biomed. Anal. 2020, 189, 113440. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Jansen, E.E.W.; Jakobs, C.; Riedl, E.; Janssen, B.; Yard, B.A.; Wedel, J.; Hoffmann, G.F.; Zschocke, J.; Gotthardt, D.; et al. Anserine inhibits carnosine degradation but in human serum carnosinase (CN1) is not correlated with histidine dipeptide concentration. Clin. Chim. Acta 2011, 412, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Mehrazad-Saber, Z.; Kheirouri, S.; Noorazar, S.-G. Effects of l-Carnosine Supplementation on Sleep Disorders and Disease Severity in Autistic Children: A Randomized, Controlled Clinical Trial. Basic Clin. Pharmacol. Toxicol. 2018, 123, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Chez, M.G.; Buchanan, C.P.; Aimonovitch, M.C.; Becker, M.; Schaefer, K.; Black, C.; Komen, J. Double-blind, placebo-controlled study of L-carnosine supplementation in children with autistic spectrum disorders. J. Child. Neurol. 2002, 17, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Baraniuk, J.N.; El-Amin, S.; Corey, R.; Rayhan, R.; Timbol, C. Carnosine Treatment for Gulf War Illness: A Randomized Controlled Trial. Glob. J. Health Sci. 2013, 5, 69–81. [Google Scholar] [CrossRef]
- Gardner, M.L.G.; Illingworth, K.M.; Kelleher, J.; Wood, D. Intestinal-Absorption of the Intact Peptide Carnosine in Man, and Comparison with Intestinal Permeability to Lactulose. J. Physiol. 1991, 439, 411–422. [Google Scholar] [CrossRef]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppermann, H.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Non-enzymatic reaction of carnosine and glyceraldehyde-3-phosphate accompanies metabolic changes of the pentose phosphate pathway. Cell Prolif. 2019, 53, e12702. [Google Scholar] [CrossRef] [Green Version]
- Remigante, A.; Morabito, R.; Marino, A. Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants 2019, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remigante, A.; Morabito, R.; Marino, A. Band 3 protein function and oxidative stress in erythrocytes. J. Cell. Physiol. 2021, 236, 6225–6234. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; van der Hoeven, L.H.; Haddox, C.H. Glutathione reductase in the red blood cells. Ann. Clin. Lab. Sci. 1978, 8, 23–29. [Google Scholar]
- Oppermann, H.; Schnabel, L.; Meixensberger, J.; Gaunitz, F. Pyruvate attenuates the anti-neoplastic effect of carnosine independently from oxidative phosphorylation. Oncotarget 2016, 7, 85848–85860. [Google Scholar] [CrossRef] [Green Version]
- Renner, C.; Asperger, A.; Seyffarth, A.; Meixensberger, J.; Gebhardt, R.; Gaunitz, F. Carnosine inhibits ATP production in cells from malignant glioma. Neurol. Res. 2010, 32, 101–105. [Google Scholar] [CrossRef]
- Letzien, U.; Oppermann, H.; Meixensberger, J.; Gaunitz, F. The antineoplastic effect of carnosine is accompanied by induction of PDK4 and can be mimicked by l-histidine. Amino Acids 2014, 46, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Glycotoxins: Dietary and Metabolic Origins; Possible Amelioration of Neurotoxicity by Carnosine, with Special Reference to Parkinson’s Disease. Neurotox. Res. 2018, 34, 164–172. [Google Scholar] [CrossRef]
- Arzumanyan, E.S.; Makhro, A.V.; Tyulina, O.V.; Boldyrev, A.A. Carnosine protects erythrocytes from the oxidative stress caused by homocysteic acid. Dokl. Biochem. Biophys. 2008, 418, 44–46. [Google Scholar] [CrossRef]
- Sariev, A.K.; Abaimov, D.A.; Tankevich, M.V.; Pantyukhova, E.Y.; Prokhorov, D.I.; Fedorova, T.N.; Lopachev, A.V.; Stvolinskii, S.L.; Konovalova, E.V.; Seifulla, R.D. Experimental study of the basic pharmacokinetic characteristics of dipeptide carnosine and its efficiency of penetration into brain tissues. Eksp. Klin. Farmakol. 2015, 78, 30–35. [Google Scholar]
- de Courten, B.; Jakubova, M.; de Courten, M.P.; Kukurova, I.J.; Vallova, S.; Krumpolec, P.; Valkovic, L.; Kurdiova, T.; Garzon, D.; Barbaresi, S.; et al. Effects of carnosine supplementation on glucose metabolism: Pilot clinical trial. Obesity 2016, 24, 1027–1034. [Google Scholar] [CrossRef]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A. l-Carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products, and tumor necrosis factor-α levels in patients with type 2 diabetes: A double-blind placebo-controlled randomized clinical trial. Nutr. Res. 2018, 49, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatr. Diabetes 2018, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.; Saleh, S.; El Abhar, H.; Saad, A.S.; Schaalan, M. L-Carnosine protects against Oxaliplatin-induced peripheral neuropathy in colorectal cancer patients: A perspective on targeting Nrf-2 and NF-κB pathways. Toxicol. Appl. Pharmacol. 2019, 365, 41–50. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Deyev, A.I.; Yermakova, V.N.; Semiletov, Y.A.; Davydova, N.G.; Kurysheva, N.I.; Zhukotskii, A.V.; Goldman, I.M. N-Acetylcarnosine, a natural histidine-containing dipeptide, as a potent ophthalmic drug in treatment of human cataracts. Peptides 2001, 22, 979–994. [Google Scholar] [CrossRef]
- Chengappa, K.R.; Turkin, S.R.; DeSanti, S.; Bowie, C.R.; Brar, J.S.; Schlicht, P.J.; Murphy, S.L.; Hetrick, M.L.; Bilder, R.; Fleet, D. A preliminary, randomized, double-blind, placebo-controlled trial of l-carnosine to improve cognition in schizophrenia. Schizophr. Res. 2012, 142, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, C.; Carubelli, V.; Lazzarini, V.; Vizzardi, E.; Bordonali, T.; Ciccarese, C.; Castrini, A.I.; Dei, C.A.; Nodari, S.; Metra, M. Effects of oral administration of orodispersible levo-carnosine on quality of life and exercise performance in patients with chronic heart failure. Nutrition 2015, 31, 71–78. [Google Scholar] [CrossRef]
- Qiu, J.; Hauske, S.J.; Zhang, S.; Rodriguez-Niño, A.; Albrecht, T.; Pastene, D.O.; van den Born, J.; van Goor, H.; Ruf, S.; Kohlmann, M.; et al. Identification and characterisation of carnostatine (SAN9812), a potent and selective carnosinase (CN1) inhibitor with in vivo activity. Amino Acids 2019, 51, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Christensen, H.N. Interorgan amino acid nutrition. Physiol. Rev. 1982, 62, 1193–1233. [Google Scholar] [CrossRef]
- Tunnicliff, G. Amino acid transport by human erythrocyte membranes. Comp. Biochem. Physiol. Part A Physiol. 1994, 108, 471–478. [Google Scholar] [CrossRef]
- Oppermann, H.; Heinrich, M.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. The proton-coupled oligopeptide transporters PEPT2, PHT1 and PHT2 mediate the uptake of carnosine in glioblastoma cells. Amino Acids 2019, 51, 999–1008. [Google Scholar] [CrossRef]
- Wilson, M.C.; Trakarnsanga, K.; Heesom, K.J.; Cogan, N.; Green, C.; Toye, A.M.; Parsons, S.F.; Anstee, D.J.; Frayne, J. Comparison of the Proteome of Adult and Cord Erythroid Cells, and Changes in the Proteome Following Reticulocyte Maturation. Mol. Cell. Proteom. 2016, 15, 1938–1946. [Google Scholar] [CrossRef] [Green Version]
- Bryk, A.H.; Wiśniewski, J.R. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [Green Version]
- Rivera, A.; Kam, S.Y.; Ho, M.; Romero, J.R.; Lee, S. Ablation of the Kell/Xk complex alters erythrocyte divalent cation homeostasis. Blood Cells Mol. Dis. 2013, 50, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Gaunitz, F.; Heise, K. HTS compatible assay for antioxidative agents using primary cultured hepatocytes. Assay Drug Dev. Technol. 2003, 1, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, H.; Purcz, K.; Birkemeyer, C.; Baran-Schmidt, R.; Meixensberger, J.; Gaunitz, F. Carnosine’s inhibitory effect on glioblastoma cell growth is independent of its cleavage. Amino Acids 2019, 51, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Strazzullo, P.; Cappuccio, F.P.; Iacoviello, L.; Cipollaro, M.; Varriale, V.; Giorgione, N.; Farinaro, E.; Mancini, M. Erythrocyte volume and blood pressure in a cross-sectional population-based study. J. Hypertens. 1990, 8, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6521–6531. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oppermann, H.; Elsel, S.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase. Int. J. Mol. Sci. 2021, 22, 12802. https://doi.org/10.3390/ijms222312802
Oppermann H, Elsel S, Birkemeyer C, Meixensberger J, Gaunitz F. Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase. International Journal of Molecular Sciences. 2021; 22(23):12802. https://doi.org/10.3390/ijms222312802
Chicago/Turabian StyleOppermann, Henry, Stefanie Elsel, Claudia Birkemeyer, Jürgen Meixensberger, and Frank Gaunitz. 2021. "Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase" International Journal of Molecular Sciences 22, no. 23: 12802. https://doi.org/10.3390/ijms222312802