
Human Recombinant DNase I (Pulmozyme®) Inhibits Lung Metastases in Murine Metastatic B16 Melanoma Model That Correlates with Restoration of the DNase Activity and the Decrease SINE/LINE and c-Myc Fragments in Blood Cell-Free DNA


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Comparison of DNase Activity of Pulmozyme® and DNase I
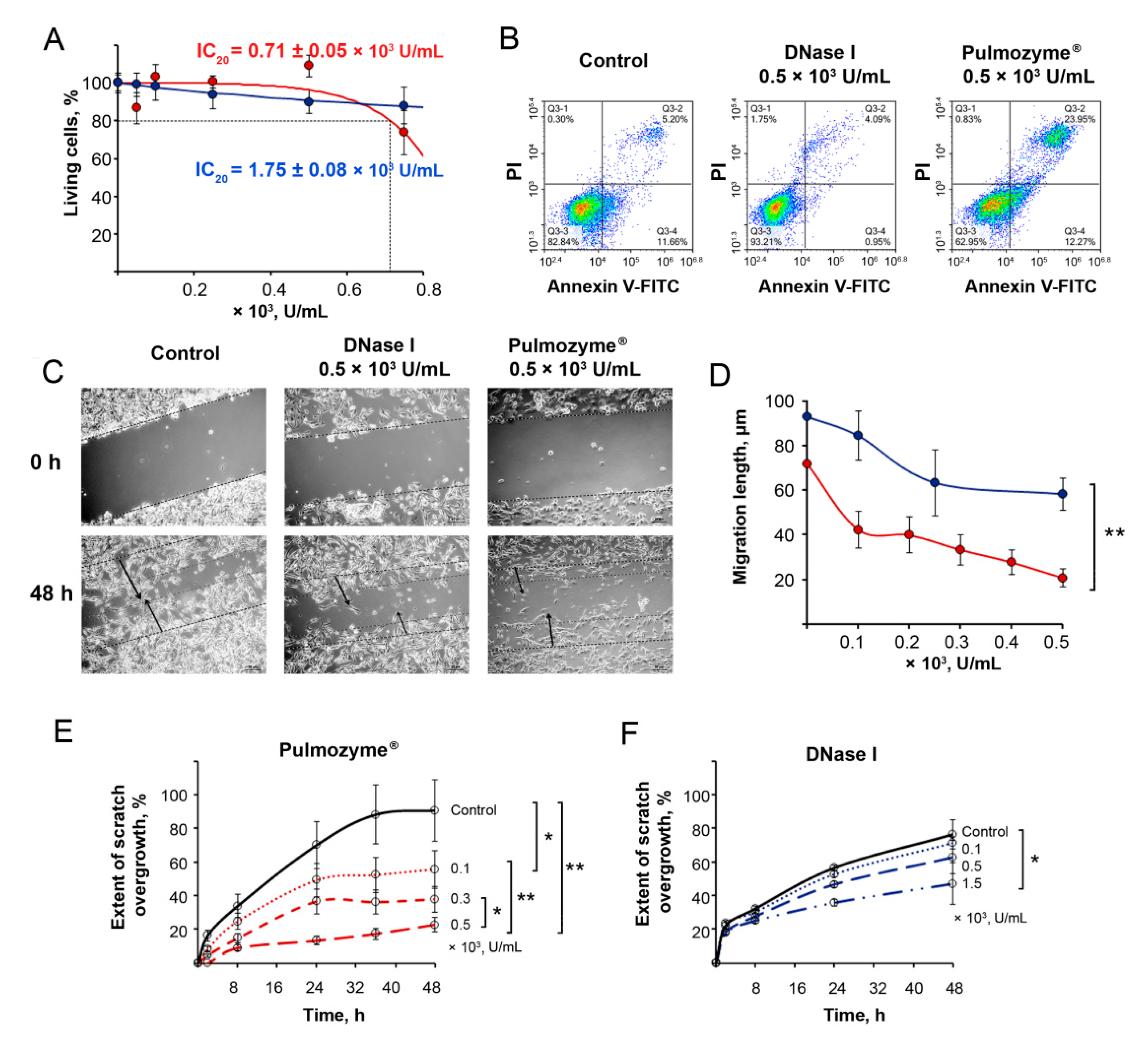
2.2. The Effect of Pulmozyme® on B16 Cell Proliferation, Apoptosis and Migration In Vitro
2.3. The Effect of Intramuscular (i.m.) Administration of Pulmozyme® on the Development of Metastases In Vivo
2.3.1. Scheme of the Intramuscular Experiment
2.3.2. Surface and Internal Metastases
2.3.3. cfDNA and DNase Activity
2.4. The Effect of the Intranasal (i.n.) Administration of Pulmozyme® on Metastasis Development In Vivo
2.4.1. Scheme of the Intranasal Experiment
2.4.2. Surface and Internal Metastases
2.4.3. cfDNA and DNase Activity
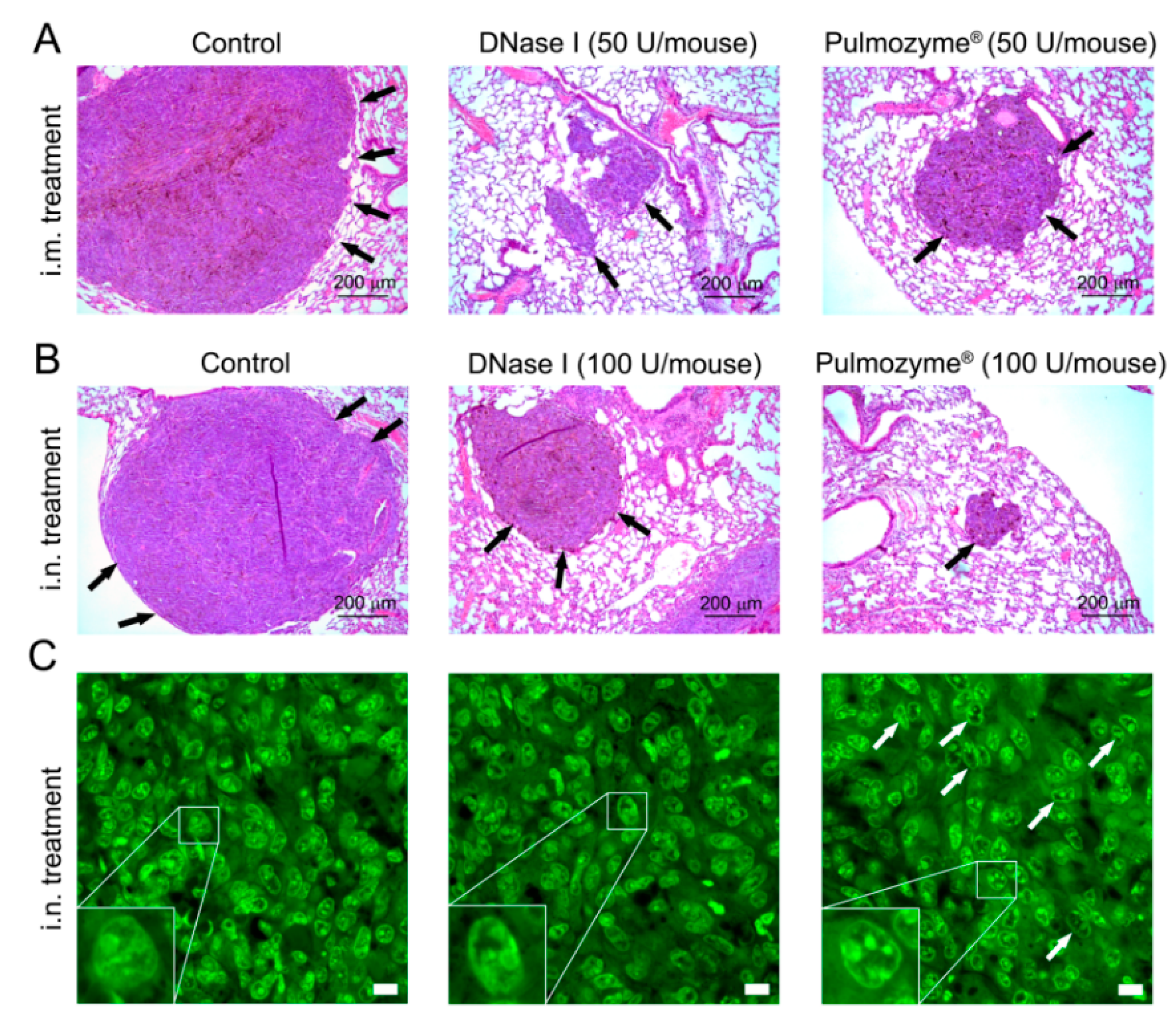
2.5. Histological Structure of Metastatic Foci in the Lungs of Mice with B16 after i.m. and i.n. Administration of Pulmozyme® and DNase I
2.6. One-Day Dynamic of cfDNA Concentration and DNase Activity in Blood Serum of Mice with B16 after i.n. Administration of Pulmozyme® and DNase I
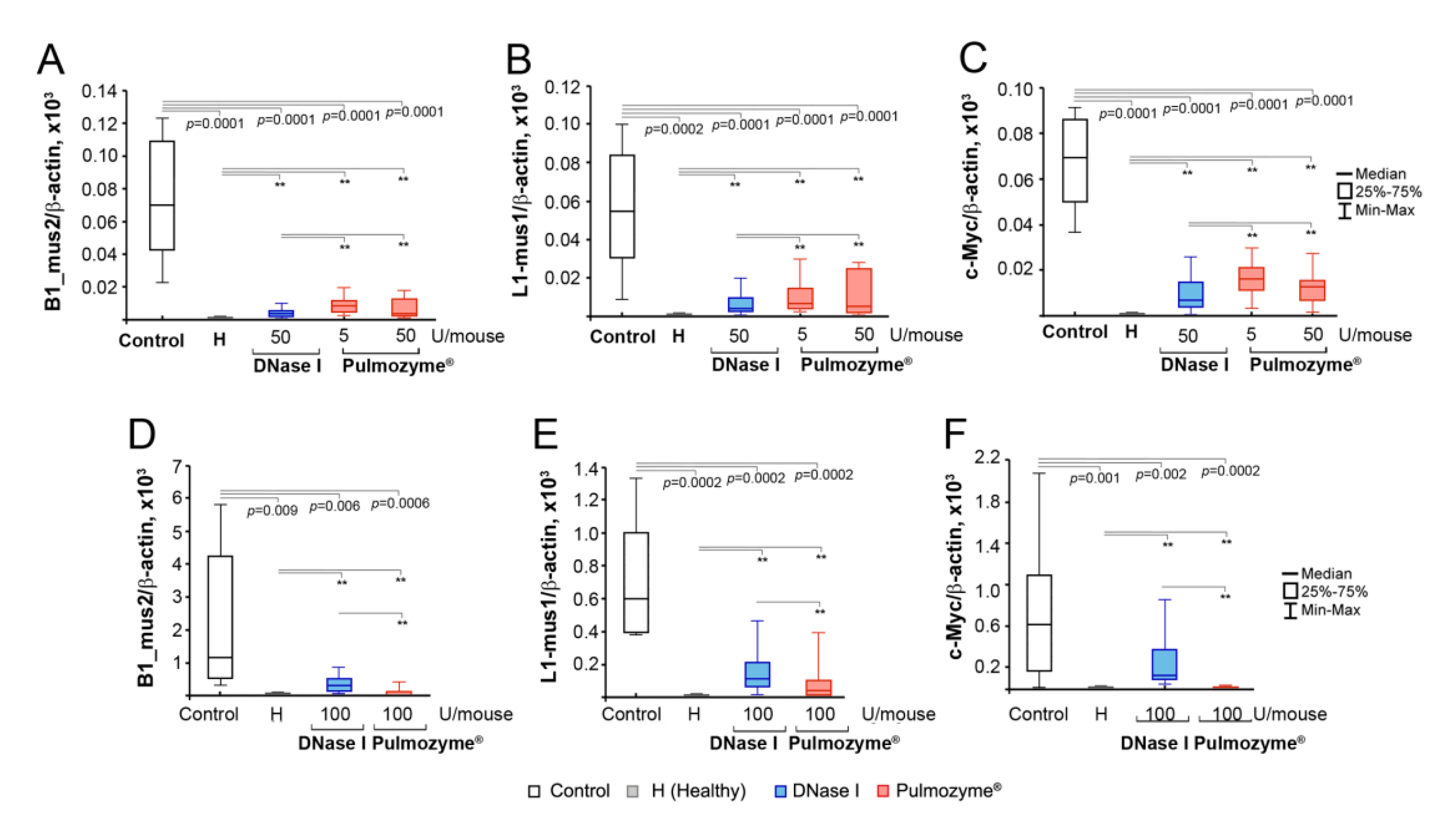
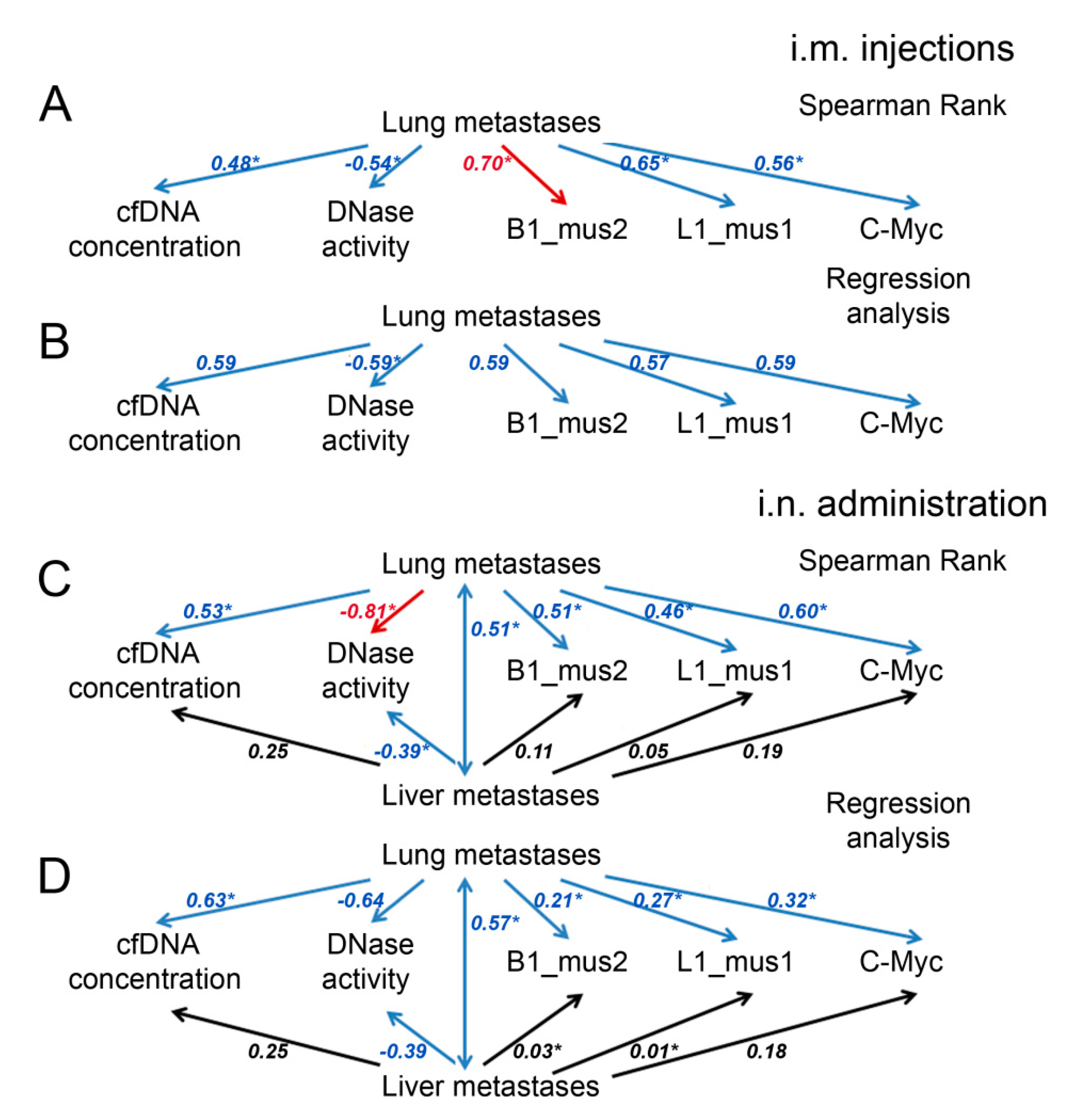
2.7. Analysis of the Abundance of SINE and LINE Elements and the c-Myc Gene in the Blood of Mice with B16
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures and Tumor Strains
4.3. Mice
4.4. The Study of Deoxyribonuclease Activity of Pulmozyme® and DNase I In Vitro
4.5. Influence of Pulmozyme® and DNase I on cfDNA In Vitro
4.6. Cell Viability Assay
4.7. Apoptosis Assay
4.8. Scratch Assay (Wound-Healing)
4.9. Tumor Implantation and Design of Animal Experiments
4.9.1. Intramuscular Experiment
4.9.2. Intranasal Experiment
4.10. Histology and Immunohistochemistry
4.11. Blood Serum Preparation
4.12. Measurement of DNase Activity in Blood Serum
4.13. Isolation of cfDNA from Blood Serum
4.14. Real-Time RT-PCR
4.15. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2018, 18, 41–58. [Google Scholar] [CrossRef]
- Cirri, D.; Bartoli, F.; Pratesi, A.; Baglini, E.; Barresi, E.; Marzo, T. Strategies for the improvement of metal-based chemotherapeutic treatments. Biomedicines 2021, 9, 504. [Google Scholar] [CrossRef]
- Costa, B.; Vale, N. A review of repurposed cancer drugs in clinical trials for potential treatment of COVID-19. Pharmaceutics 2021, 13, 815. [Google Scholar] [CrossRef]
- Lorkowski, J.; Kolaszynska, O.; Pokorski, M. Artificial intelligence and precision medicine: A perspective. Adv. Exp. Med. Biol. 2021, 18, 18. [Google Scholar] [CrossRef]
- Yang, C.; Chilvers, M.; Montgomery, M.; Nolan, S.J. Dornase alfa for cystic fibrosis. Cochrane Database Syst. Rev. 2016, 2016, CD001127. [Google Scholar] [CrossRef] [Green Version]
- Robinson, C. Dornase alfa. Drugs Future 1994, 19, 542–545. [Google Scholar] [CrossRef]
- Den Hollander, B.; Linssen, R.S.N.; Cortjens, B.; Van Etten-Jamaludin, F.S.; Van Woensel, J.B.M.; Bem, R.A. Use of dornase alfa in the paediatric intensive care unit: Current literature and a national cross-sectional survey. Eur. J. Hosp. Pharm. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.H.; Machado, C.C.; Veras, F.P.; de Maganin, A.G.M.; de Souza, F.F.L.; Barroso, L.C.; de Oliveira, R.D.R.; Alves-Filho, J.C.; Cunha, T.M.; Fukada, S.Y.; et al. Neutrophil extracellular traps mediate joint hyperalgesia induced by immune inflammation. Rheumatology 2020, 60, 3461–3473. [Google Scholar] [CrossRef] [PubMed]
- Mulay, S.R.; Anders, H.J. Neutrophils and neutrophil extracellular traps regulate immune responses in health and disease. Cells 2020, 9, 2130. [Google Scholar] [CrossRef] [PubMed]
- Assallum, H.; Song, T.Y.; DeLorenzo, L.; Harris, K. Bronchoscopic instillation of DNase to manage refractory lobar atelectasis in a lung cancer patient. Ann. Transl. Med. 2019, 7, 363. [Google Scholar] [CrossRef]
- Salganik, R.I.; Martynova, R.P.; Matienko, N.A.; Ronichevskaya, G.M. Effect of deoxyribonuclease on the course of lymphatic leukaemia in AKR mice. Nature 1967, 214, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, S.; Yamamoto, T.; Tanaka, H.; Kambara, T.; Hiraoka, T.; Miyauchi, Y. Deoxyribonuclease treatment prevents blood-borne liver metastasis of cutaneously transplanted tumour cells in mice. Br. J. Cancer 1993, 67, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patutina, O.; Mironova, N.; Ryabchikova, E.; Popova, N.; Nikolin, V.; Kaledin, V.; Vlassov, V.; Zenkova, M. Inhibition of metastasis development by daily administration of ultralow doses of RNase A and DNase I. Biochimie 2011, 93, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Alexeeva, L.A.; Patutina, O.A.; Sen’kova, A.V.; Zenkova, M.A.; Mironova, N.L. Inhibition of invasive properties of murine melanoma by bovine pancreatic DNase I in vitro and in vivo. Mol. Biol. 2017, 51, 637–646. [Google Scholar] [CrossRef]
- Trejo-Becerril, C.; Pérez-Cardenas, E.; Gutiérrez-Díaz, B.; De La Cruz-Sigüenza, D.; Taja-Chayeb, L.; González-Ballesteros, M.; García-López, P.; Chanona, J.; Dueñas-González, A. Antitumor effects of systemic DNAse I and proteases in an in vivo model. Integr. Cancer Ther. 2016, 15, NP35–NP43. [Google Scholar] [CrossRef] [Green Version]
- Wen, F.; Shen, A.; Choi, A.; Gerner, E.W.; Shi, J. Extracellular DNA in pancreatic cancer promotes cell invasion and metastasis. Cancer Res. 2013, 73, 4256–4266. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Yao, H.; Liu, Z.; Xu, M.; Tsung, A.; Wang, Y. Endogenous PAD4 in breast cancer cells mediates cancer extracellular chromatin network formation and promotes lung metastasis. Mol. Cancer Res. 2020, 18, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Takesue, S.; Ohuchida, K.; Shinkawa, T.; Otsubo, Y.; Matsumoto, S.; Sagara, A.; Yonenaga, A.; Ando, Y.; Kibe, S.; Nakayama, H.; et al. Neutrophil extracellular traps promote liver micrometastasis in pancreatic ductal adenocarcinoma via the activation of cancer-associated fibroblasts. Int. J. Oncol. 2020, 56, 596–605. [Google Scholar] [CrossRef]
- Alekseeva, L.A.; Mironova, N.L.; Brenner, E.V.; Kurilshikov, A.M.; Patutina, O.A.; Zenkova, M.A. Alteration of the exDNA profile in blood serum of LLC-bearing mice under the decrease of tumour invasion potential by bovine pancreatic DNase I treatment. PLoS ONE 2017, 12, e0171988. [Google Scholar] [CrossRef] [Green Version]
- Alekseeva, L.A.; Sen’kova, A.V.; Zenkova, M.A.; Mironova, N.L. Targeting circulating SINEs and LINEs with DNase I provides metastases inhibition in experimental tumor models. Mol. Ther. Nucleic Acids 2020, 20, 50–61. [Google Scholar] [CrossRef]
- Combaret, V.; Audoynaud, C.; Iacono, I.; Favrot, M.C.; Schell, M.; Bergeron, C.; Puisieux, A. Circulating MYCN DNA as a tumor-specific marker in neuroblastoma patients. Cancer Res. 2002, 62, 3646–3648. [Google Scholar]
- Sorensen, B.S.; Mortensen, L.S.; Andersen, J.; Nexo, E. Circulating HER2 DNA after trastuzumab treatment predicts survival and response in breast cancer. Anticancer Res. 2010, 30, 2463–2468. [Google Scholar]
- Forschner, A.; Hilke, F.J.; Bonzheim, I.; Gschwind, A.; Demidov, G.; Amaral, T.; Ossowski, S.; Riess, O.; Schroeder, C.; Martus, P.; et al. MDM2, MDM4 and EGFR amplifications and hyperprogression in metastatic acral and mucosal melanoma. Cancers 2020, 12, 540. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Galindo, R.; Kaplan, M.H.; Leissner, P.; Verjat, T.; Ferlenghi, I.; Bagnoli, F.; Giusti, F.; Dosik, M.H.; Hayes, D.F.; Gitlin, S.D.; et al. Human endogenous retrovirus K (HML-2) elements in the plasma of people with lymphoma and breast cancer. J. Virol. 2008, 82, 9329–9336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Seo, A.N.; Jung, H.Y.; Gwak, J.M.; Jung, N.; Cho, N.Y.; Kang, G.H. Alu and LINE-1 hypomethylation is associated with HER2 enriched subtype of breast cancer. PLoS ONE 2014, 9, e100429. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Zhu, Y.M.; Woll, P.J. Circulating DNA and lung cancer. Ann. N. Y. Acad. Sci. 2006, 1075, 154–164. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef]
- Belancio, V.P.; Roy-Engel, A.M.; Deininger, P.L. All y’all need to know ‘bout retroelements in cancer. Semin. Cancer Biol. 2010, 20, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Carreira, P.E.; Richardson, S.R.; Faulkner, G.J. L1 retrotransposons, cancer stem cells and oncogenesis. FEBS J. 2014, 281, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, J.R.; Longworth, M.S. Crossing the LINE toward genomic instability: LINE-1 retrotransposition in cancer. Front. Chem. 2015, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Helman, E.; Lawrence, M.S.; Stewart, C.; Sougnez, C.; Getz, G.; Meyerson, M. Somatic retrotransposition in human cancer revealed by whole-genome and exome sequencing. Genome Res. 2014, 24, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martín, C.; Cidre, F.; Fernández-Teijeiro, A.; Gómez-Mariano, G.; De La Vega, L.; Ramos, P.; Zaballos, A.; Monzón, S.; Alonso, J. Familial retinoblastoma due to intronic LINE-1 insertion causes aberrant and noncanonical mRNA splicing of the RB1 gene. J. Hum. Genet. 2016, 61, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Stacey, S.N.; Kehr, B.; Gudmundsson, J.; Zink, F.; Jonasdottir, A.; Gudjonsson, S.A.; Sigurdsson, A.; Halldorsson, B.V.; Agnarsson, B.A.; Benediktsdottir, K.R.; et al. Insertion of an SVA-E retrotransposon into the CASP8 gene is associated with protection against prostate cancer. Hum. Mol. Genet. 2016, 25, 1008. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Bian, Y.; Luo, X.; Wang, C.; Mu, D.; Pan, G.; Wu, J.; Shi, H. Synergistic effect of docosahexaenoic acid or conjugated linoleic acid with caffeic acid on ameliorating oxidative stress of HepG2 cells. J. Food Sci. 2021, 86, 3240–3251. [Google Scholar] [CrossRef] [PubMed]
- Inokuchi, S.; Mitoma, H.; Kawano, S.; Nakano, S.; Ayano, M.; Kimoto, Y.; Akahoshi, M.; Arinobu, Y.; Tsukamoto, H.; Akashi, K.; et al. Homeostatic milieu induces production of Deoxyribonuclease 1–like 3 from myeloid cells. J. Immunol. 2020, 204, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Pisko, J.; Špirková, A.; Čikoš, Š.; Olexiková, L.; Kovaříková, V.; Šefčíková, Z.; Fabian, D. Apoptotic cells in mouse blastocysts are eliminated by neighbouring blastomeres. Sci. Rep. 2021, 11, 9228. [Google Scholar] [CrossRef]
- Yasuda, T.; Kawai, Y.; Ueki, M.; Kishi, K. Clinical applications of DNase I, a genetic marker already used for forensic identification. Leg. Med. 2005, 7, 274–277. [Google Scholar] [CrossRef]
- Okur, H.K.; Yalcin, K.; Tastan, C.; Demir, S.; Yurtsever, B.; Karakus, G.S.; Kancagi, D.D.; Abanuz, S.; Seyis, U.; Zengin, R.; et al. Preliminary report of in vitro and in vivo effectiveness of dornase alfa on SARS-CoV-2 infection. New Microbes New Infect. 2020, 37, 100756. [Google Scholar] [CrossRef]
- Chhabra, D.; Bao, S.; Dos Remedios, C.G. The distribution of cofilin and DNase I in vivo. Cell Res. 2002, 12, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Boddy, J.L.; Gal, S.; Malone, P.R.; Harris, A.L.; Wainscoat, J.S. Prospective study of quantitation of plasma DNA levels in the diagnosis of malignant versus benign prostate disease. Clin. Cancer Res. 2005, 11, 1394–1399. [Google Scholar] [CrossRef] [Green Version]
- Sunami, E.; Vu, A.T.; Nguyen, S.L.; Giuliano, A.E.; Hoon, D.S.B. Quantification of LINE1 in circulating DNA as a molecular biomarker of breast cancer. Ann. N. Y. Acad. Sci. 2008, 1137, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Kamat, A.A.; Baldwin, M.; Urbauer, D.; Dang, D.; Han, L.Y.; Godwin, A.; Karlan, B.Y.; Simpson, J.L.; Gershenson, D.M.; Coleman, R.L.; et al. Plasma cell-free DNA in ovarian cancer: An independent prognostic biomarker. Cancer 2010, 116, 1918–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimberger, P.; Roth, C.; Pantel, K.; Kasimir-Bauer, S.; Kimmig, R.; Schwarzenbach, H. Impact of platinum-based chemotherapy on circulating nucleic acid levels, protease activities in blood and disseminated tumor cells in bone marrow of ovarian cancer patients. Int. J. Cancer 2011, 128, 2572–2580. [Google Scholar] [CrossRef] [PubMed]
- Tamkovich, S.N.; Cherepanova, A.V.; Kolesnikova, E.V.; Rykova, E.Y.; Pyshnyi, D.V.; Vlassov, V.V.; Laktionov, P.P. Circulating DNA and DNase activity in human blood. Ann. N. Y. Acad. Sci. 2006, 1075, 191–196. [Google Scholar] [CrossRef]
- Cherepanova, A.V.; Tamkovich, S.N.; Vlassov, V.V.; Laktionov, P.P. Blood deoxyribonuclease activity in health and diseases. Biomed. Khim. 2007, 53, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Golonka, R.M.; Yeoh, B.S.; Petrick, J.L.; Weinstein, S.J.; Albanes, D.; Gewirtz, A.T.; McGlynn, K.A.; Vijay-Kumar, M. Deoxyribonuclease I activity, cell-free DNA, and risk of liver cancer in a prospective cohort. JNCI Cancer Spectr. 2018, 2, pky083. [Google Scholar] [CrossRef] [Green Version]
- García-Olmo, D.C.; Domínguez, C.; García-Arranz, M.; Anker, P.; Stroun, M.; García-Verdugo, J.M.; García-Olmo, D. Cell-free nucleic acids circulating in the plasma of colorectal cancer patients induce the oncogenic transformation of susceptible cultured cells. Cancer Res. 2010, 70, 560–567. [Google Scholar] [CrossRef] [Green Version]
- Ehnfors, J.; Kost-Alimova, M.; Persson, N.L.; Bergsmedh, A.; Castro, J.; Levchenko-Tegnebratt, T.; Yang, L.; Panaretakis, T.; Holmgren, L. Horizontal transfer of tumor DNA to endothelial cells in vivo. Cell Death Differ. 2009, 16, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Becerril, C.; Pérez-Cárdenas, E.; Taja-Chayeb, L.; Anker, P.; Herrera-Goepfert, R.; Medina-Velázquez, L.A.; Hidalgo-Miranda, A.; Pérez-Montiel, D.; Chávez-Blanco, A.; Cruz-Velázquez, J.; et al. Cancer progression mediated by horizontal gene transfer in an in vivo model. PLoS ONE 2012, 7, e52754. [Google Scholar] [CrossRef] [Green Version]
- Gaiffe, E.; Prétet, J.L.; Launay, S.; Jacquin, E.; Saunier, M.; Hetzel, G.; Oudet, P.; Mougin, C. Apoptotic HPV positive cancer cells exhibit transforming properties. PLoS ONE 2012, 7, e36766. [Google Scholar] [CrossRef]
- Takei, H.; Araki, A.; Watanabe, H.; Ichinose, A.; Sendo, F. Rapid killing of human neutrophils by the potent activator phorbol 12-myristate 13-acetate (PMA) accompanied by changes different from typical apoptosis or necrosis. J. Leukoc. Biol. 1996, 59, 229–240. [Google Scholar] [CrossRef]
- Prosser, K.E.; Chang, S.W.; Saraci, F.; Le, P.H.; Walsby, C.J. Anticancer copper pyridine benzimidazole complexes: ROS generation, biomolecule interactions, and cytotoxicity. J. Inorg. Biochem. 2017, 167, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Ting, R.; Thomas, J.M.; Lermer, L.; Perrin, D.M. Substrate specificity and kinetic framework of a DNAzyme with an expanded chemical repertoire: A putative RNaseA mimic that catalyzes RNA hydrolysis independent of a divalent metal cation. Nucleic Acids Res. 2004, 32, 6660–6672. [Google Scholar] [CrossRef] [PubMed]
- Park, J.G.; Kramer, B.S.; Carmichael, J.; Minna, J.D.; Gazdar, A.F.; Steinberg, S.M.; Collins, J.M. Chemosensitivity testing of human colorectal carcinoma cell lines using a tetrazolium-based colorimetric assay. Cancer Res. 1987, 47, 5875–5879. [Google Scholar] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Schumperli, D.; Howard, B.H.; Rosenberg, M. Efficient expression of Escherichia coli galactokinase gene in mammalian cells. Proc. Natl. Acad. Sci. USA 1982, 79, 257–261. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekseeva, L.; Sen’kova, A.; Savin, I.; Zenkova, M.; Mironova, N. Human Recombinant DNase I (Pulmozyme®) Inhibits Lung Metastases in Murine Metastatic B16 Melanoma Model That Correlates with Restoration of the DNase Activity and the Decrease SINE/LINE and c-Myc Fragments in Blood Cell-Free DNA. Int. J. Mol. Sci. 2021, 22, 12074. https://doi.org/10.3390/ijms222112074
Alekseeva L, Sen’kova A, Savin I, Zenkova M, Mironova N. Human Recombinant DNase I (Pulmozyme®) Inhibits Lung Metastases in Murine Metastatic B16 Melanoma Model That Correlates with Restoration of the DNase Activity and the Decrease SINE/LINE and c-Myc Fragments in Blood Cell-Free DNA. International Journal of Molecular Sciences. 2021; 22(21):12074. https://doi.org/10.3390/ijms222112074
Chicago/Turabian StyleAlekseeva, Ludmila, Aleksandra Sen’kova, Innokenty Savin, Marina Zenkova, and Nadezhda Mironova. 2021. "Human Recombinant DNase I (Pulmozyme®) Inhibits Lung Metastases in Murine Metastatic B16 Melanoma Model That Correlates with Restoration of the DNase Activity and the Decrease SINE/LINE and c-Myc Fragments in Blood Cell-Free DNA" International Journal of Molecular Sciences 22, no. 21: 12074. https://doi.org/10.3390/ijms222112074