
Towards a Cardoon (Cynara cardunculus var. altilis)-Based Biorefinery: A Case Study of Improved Cell Cultures via Genetic Modulation of the Phenylpropanoid Pathway

 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Development of a Method for the Stable Transformation of Cardoon
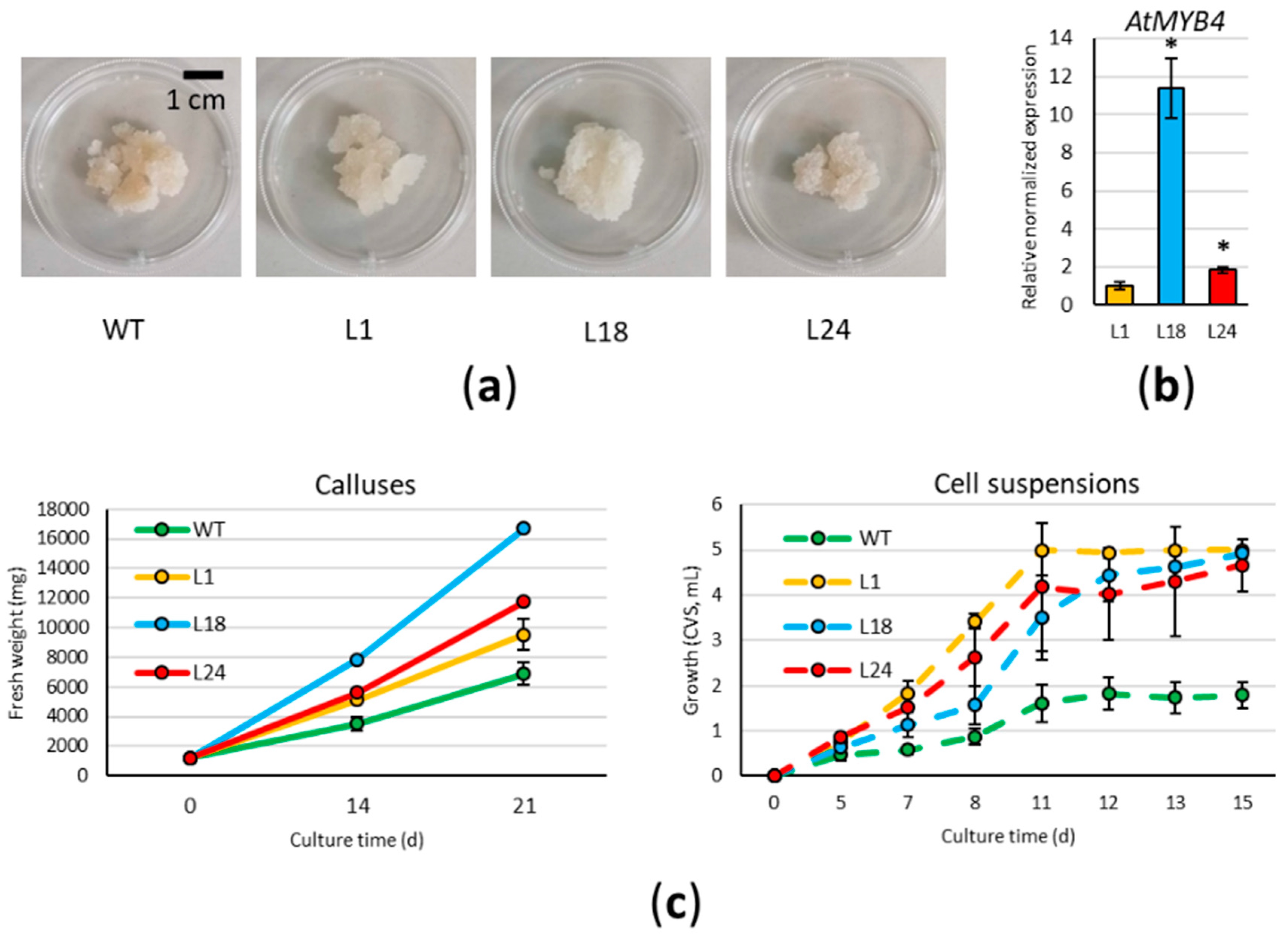
2.2. AtMYB4oe Lines Have Higher Growth Rates Than Wild Type
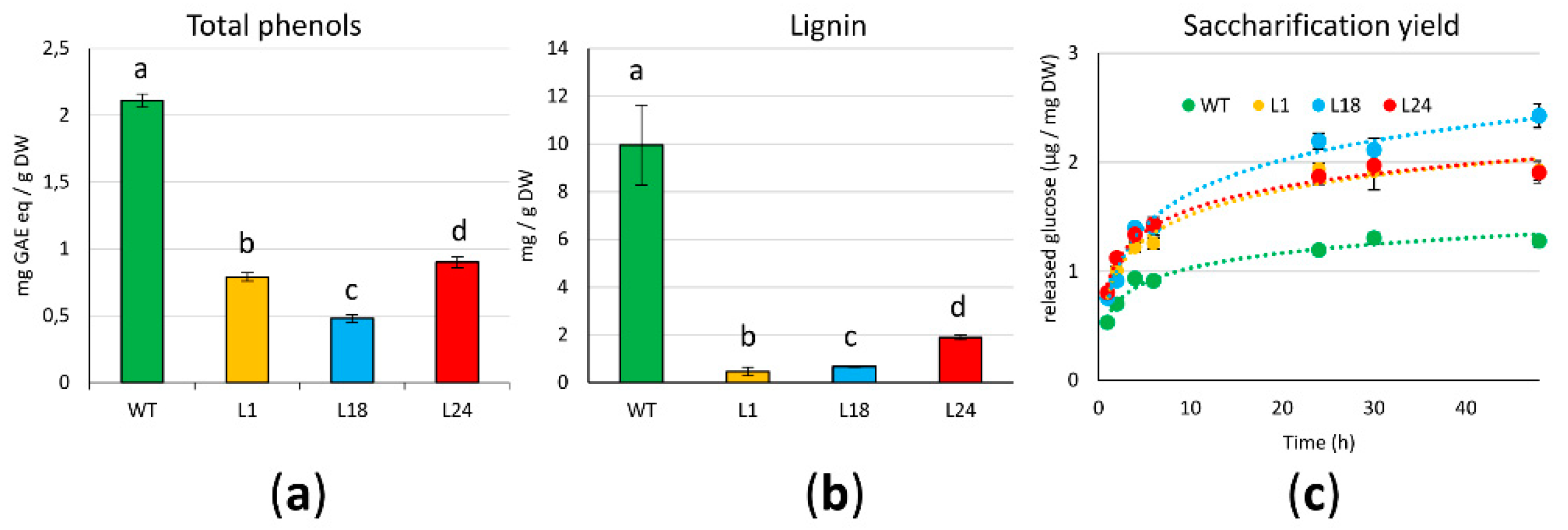
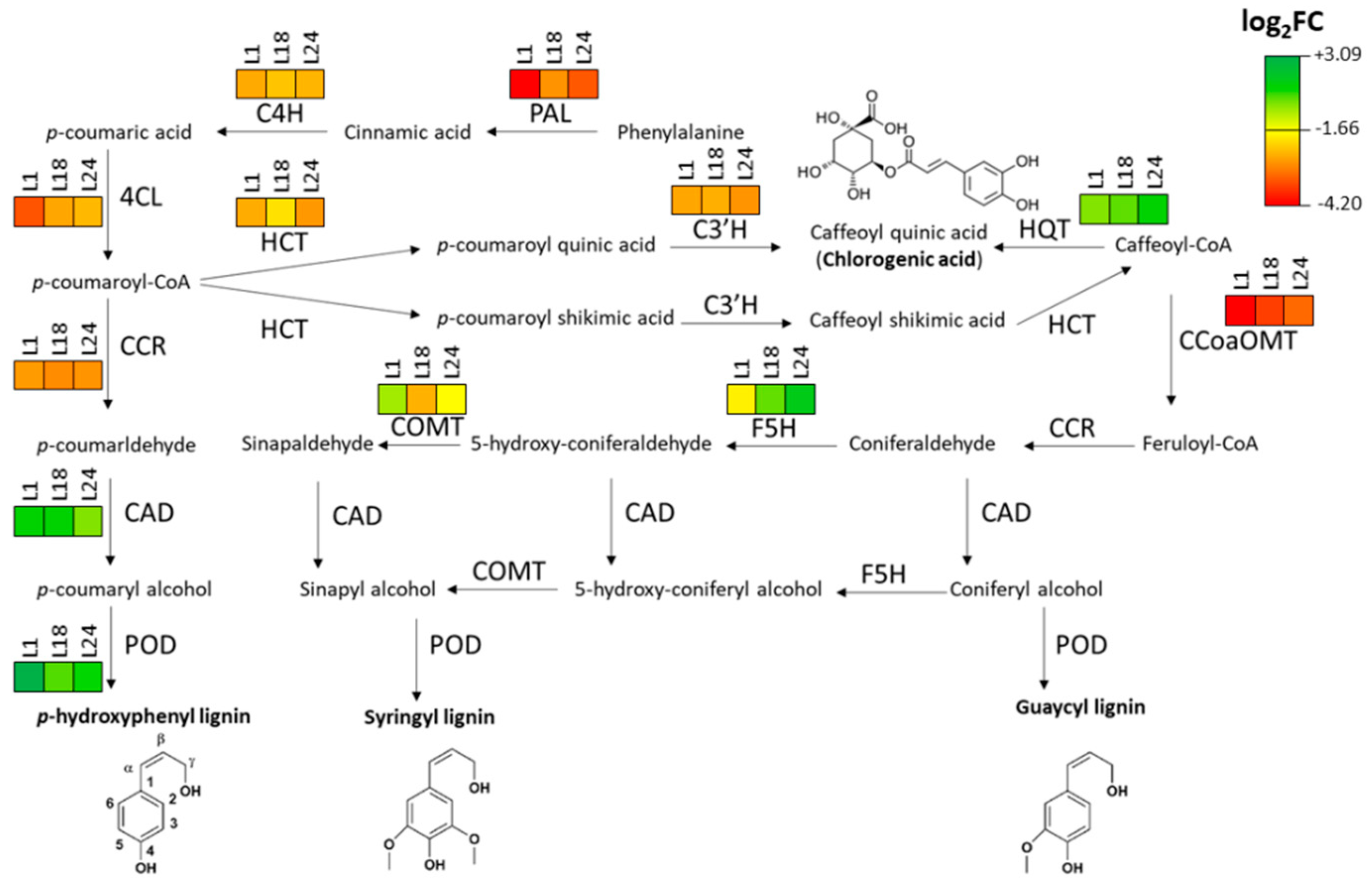
2.3. AtMYB4oe Lines Show Decreased Phenolic Compounds and Lignin Content and Enhanced Enzymatic Saccharification Efficiency
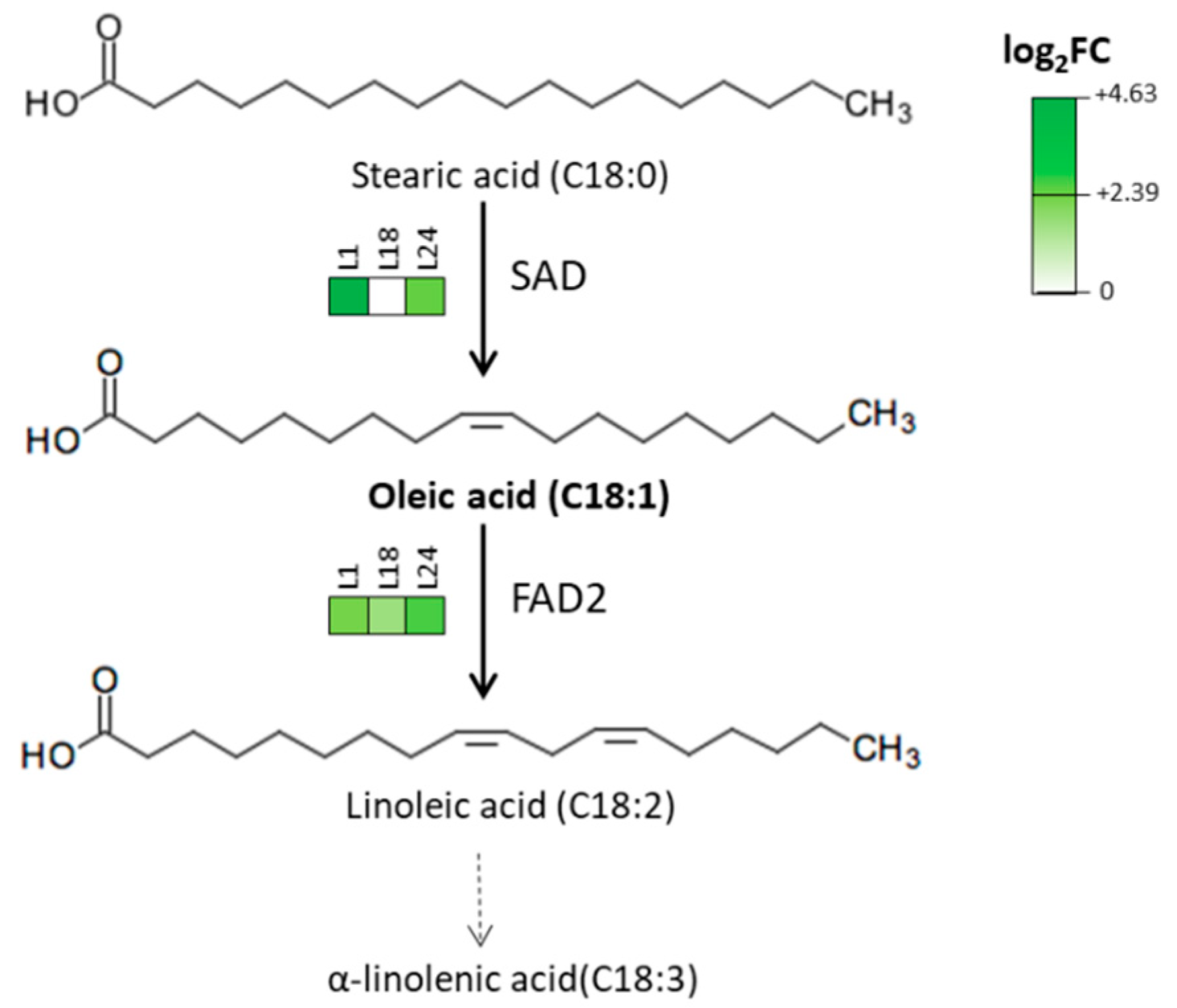
2.4. AtMYB4oe Lines Show a Different Profile of Polyphenolic Compounds and Fatty Acids Compared to Wild Type
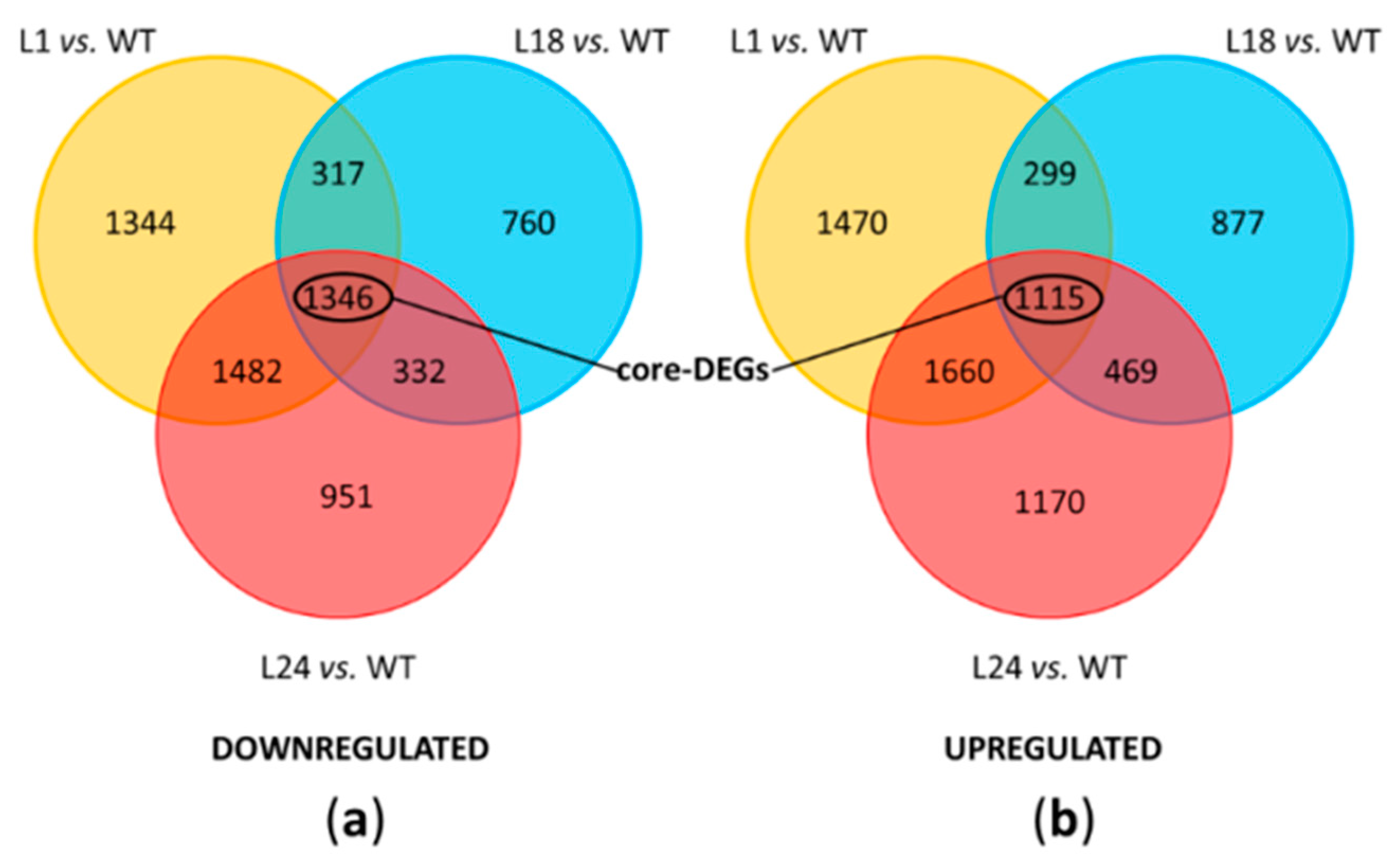
2.5. Transcriptomic Analysis of AtMYB4oe Lines
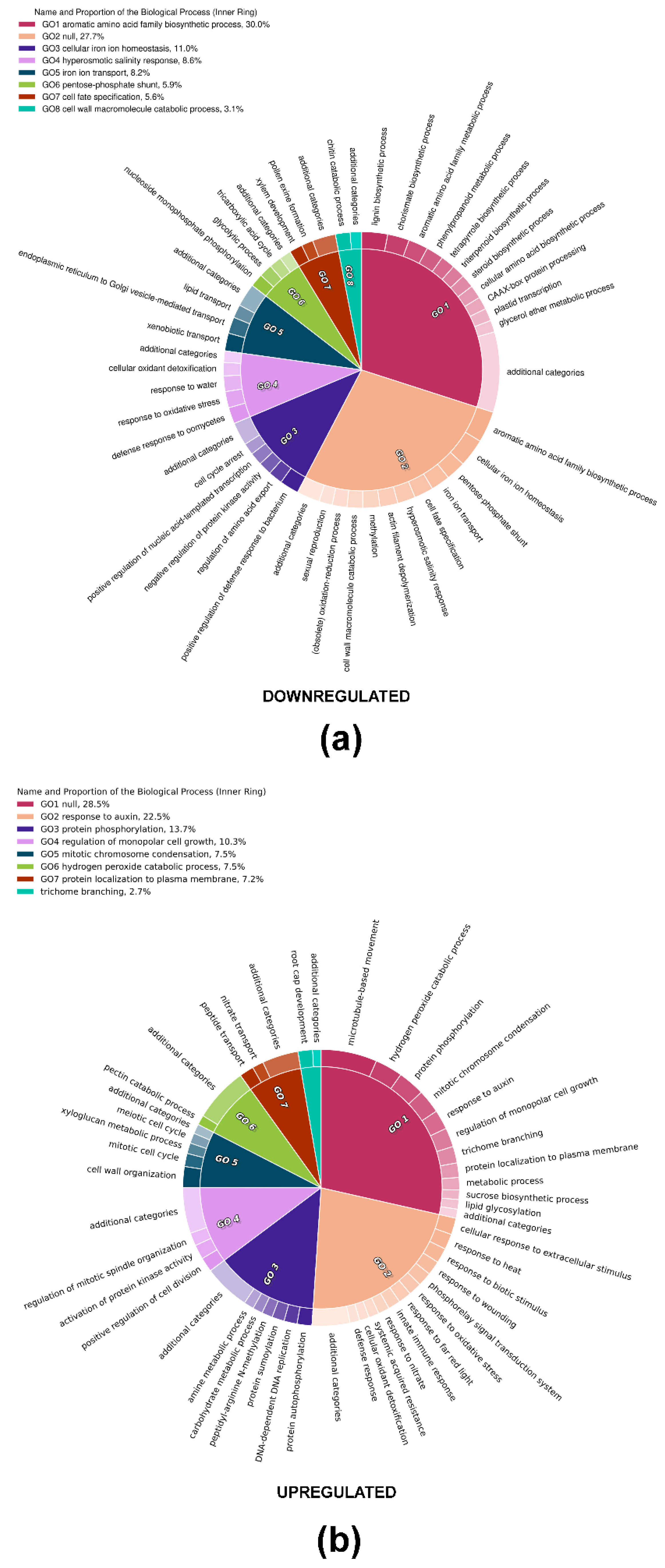
2.6. Gene Ontology Enrichment Analysis (GOEA)
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth
4.2. Construct Preparation
4.3. Cardoon Cell Suspension Transformation and Selection
4.4. Quantification of Total Phenols
4.5. Quantification of Total Lignin Content
4.6. Saccharification and Evaluation of Glucose Content of Cell Walls
4.7. Extraction of Polyphenols
4.8. Antioxidant Activity Determination
4.9. UHPLC-HRMS Analysis of Polyphenols
4.10. Oil Extraction
4.11. Fatty Acids Analysis
4.12. RNA Extraction, cDNA Preparation and Expression Analysis via q-PCR
4.13. RNA-Seq, Read Mapping, Gene Expression Evaluation, GOEA Analysis and TF Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barracosa, P.; Barracosa, M.; Pires, E. Cardoon as a Sustainable Crop for Biomass and Bioactive Compounds Production. Chem. Biodivers. 2019, 16, e1900498. [Google Scholar] [CrossRef]
- Basnizki, J.; Zohary, D. Breeding of Seed-Planted Artichoke. In Plant Breeding Reviews; John Wiley & Sons, Inc.: Oxford, UK, 2010; pp. 253–269. [Google Scholar]
- Acquadro, A.; Barchi, L.; Portis, E.; Mangino, G.; Valentino, D.; Mauromicale, G.; Lanteri, S. Genome reconstruction in Cynara cardunculus taxa gains access to chromosome-scale DNA variation. Sci. Rep. 2017, 7, 5617. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.M.; Simões, I. Cardoon-based rennets for cheese production. Appl. Microbiol. Biotechnol. 2018, 102, 4675–4686. [Google Scholar] [CrossRef] [PubMed]
- Toscano, V.; Sollima, L.; Genovese, C.; Melilli, M.G.; Raccuia, S.A. Pilot plant system for biodiesel and pellet production from cardoon: Technical and economic feasibility. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science, La Plata, Argentina, 17 November 2016; Volume 1147, pp. 429–442. [Google Scholar]
- Pappalardo, H.D.; Toscano, V.; Puglia, G.D.; Genovese, C.; Raccuia, S.A. Cynara cardunculus L. as a Multipurpose Crop for Plant Secondary Metabolites Production in Marginal Stressed Lands. Front. Plant Sci. 2020, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Gominho, J.; Curt, M.D.; Lourenço, A.; Fernández, J.; Pereira, H. Cynara cardunculus L. as a biomass and multi-purpose crop: A review of 30 years of research. Biomass Bioenergy 2018, 109, 257–275. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Docimo, T.; De Stefano, R.; Cappetta, E.; Piccinelli, A.L.; Celano, R.; De Palma, M.; Tucci, M. Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes. Plants 2020, 9, 554. [Google Scholar] [CrossRef]
- Gominho, J.; Lourenço, A.; Palma, P.; Lourenço, M.E.; Curt, M.D.; Fernández, J.; Pereira, H. Large scale cultivation of Cynara cardunculus L. for biomass production—A case study. Ind. Crop. Prod. 2011, 33, 1–6. [Google Scholar] [CrossRef]
- Sorrentino, M.C.; Capozzi, F.; Amitrano, C.; Giordano, S.; Arena, C.; Spagnuolo, V. Performance of three cardoon cultivars in an industrial heavy metal-contaminated soil: Effects on morphology, cytology and photosynthesis. J. Hazard. Mater. 2018, 351, 131–137. [Google Scholar] [CrossRef]
- Capozzi, F.; Sorrentino, M.C.; Caporale, A.G.; Fiorentino, N.; Giordano, S.; Spagnuolo, V. Exploring the phytoremediation potential of Cynara cardunculus: A trial on an industrial soil highly contaminated by heavy metals. Environ. Sci. Pollut. Res. 2020, 27, 9075–9084. [Google Scholar] [CrossRef]
- Piscioneri, I.; Sharma, N.; Baviello, G.; Orlandini, S. Promising industrial energy crop, Cynara cardunculus: A potential source for biomass production and alternative energy. Energy Convers. Manag. 2000, 41, 1091–1105. [Google Scholar] [CrossRef]
- Fernández, J.; Curt, M.D.; Aguado, P.L. Industrial applications of Cynara cardunculus L. for energy and other uses. Ind. Crop. Prod. 2006, 24, 222–229. [Google Scholar] [CrossRef]
- Mantineo, M.; D’Agosta, G.M.; Copani, V.; Patanè, C.; Cosentino, S.L. Biomass yield and energy balance of three perennial crops for energy use in the semi-arid Mediterranean environment. Field Crop. Res. 2009, 114, 204–213. [Google Scholar] [CrossRef]
- Foti, S.; Mauromicale, G.; Raccuia, S.A.; Fallico, B.; Fanella, F.; Maccarone, E. Possible alternative utilization of Cynara spp. I. Biomass, grain yield and chemical composition of grain. Ind. Crop. Prod. 1999, 10, 219–228. [Google Scholar] [CrossRef]
- Ierna, A.; Sortino, O.; Mauromicale, G. Biomass, seed and energy yield of Cynara cardunculus L. as affected by environment and season. Agronomy 2020, 10, 1584. [Google Scholar] [CrossRef]
- Gominho, J.; Fernandez, J.; Pereira, H. Cynara cardunculus L. A new fibre crop for pulp and paper production. Ind. Crop. Prod. 2001, 13, 1–10. [Google Scholar] [CrossRef]
- Barbosa, C.H.; Andrade, M.A.; Vilarinho, F.; Castanheira, I.; Fernando, A.L.; Loizzo, M.R.; Silva, A.S. A new insight on cardoon: Exploring new uses besides cheese making with a view to zero waste. Foods 2020, 9, 564. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G.; Williamson, G. Phenolic acids and flavonoids in leaf and floral stem of cultivated and wild Cynara cardunculus L. genotypes. Food Chem. 2011, 126, 417–422. [Google Scholar] [CrossRef]
- Pinelli, P.; Agostini, F.; Comino, C.; Lanteri, S.; Portis, E.; Romani, A. Simultaneous quantification of caffeoyl esters and flavonoids in wild and cultivated cardoon leaves. Food Chem. 2007, 105, 1695–1701. [Google Scholar] [CrossRef]
- Graziani, G.; Docimo, T.; De Palma, M.; Sparvoli, F.; Izzo, L.; Tucci, M.; Ritieni, A. Changes in Phenolics and Fatty Acids Composition and Related Gene Expression during the Development from Seed to Leaves of Three Cultivated Cardoon Genotypes. Antioxidants 2020, 9, 1096. [Google Scholar] [CrossRef]
- Kollia, E.; Markaki, P.; Zoumpoulakis, P.; Proestos, C. Antioxidant activity of Cynara scolymus L. and Cynara cardunculus L. extracts obtained by different extraction techniques. Nat. Prod. Res. 2017, 31, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Falleh, H.; Ksouri, R.; Chaieb, K.; Karray-Bouraoui, N.; Trabelsi, N.; Boulaaba, M.; Abdelly, C. Phenolic composition of Cynara cardunculus L. organs, and their biological activities. Comptes Rendus Biol. 2008, 331, 372–379. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Mehmetçik, G.; Özdemirler, G.; Koçak-Toker, N.; Çevikbaş, U.; Uysal, M. Effect of pretreatment with artichoke extract on carbon tetrachloride-induced liver injury and oxidative stress. Exp. Toxicol. Pathol. 2008, 60, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Küçükgergin, C.; Aydın, A.F.; Özdemirler-Erata, G.; Mehmetçik, G.; Koçak-Toker, N.; Uysal, M. Effect of artichoke leaf extract on hepatic and cardiac oxidative stress in rats fed on high cholesterol diet. Biol. Trace Elem. Res. 2010, 135, 264–274. [Google Scholar] [CrossRef]
- Kukić, J.; Popović, V.; Petrović, S.; Mucaji, P.; Ćirić, A.; Stojković, D.; Soković, M. Antioxidant and antimicrobial activity of Cynara cardunculus extracts. Food Chem. 2008, 107, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.K.; Sharma, S. Recent updates on different methods of pretreatment of lignocellulosic feedstocks: A review. Bioresour. Bioprocess. 2017, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Halpin, C. Lignin engineering to improve saccharification and digestibility in grasses. Curr. Opin. Biotechnol. 2019, 56, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Maccarone, E.; Fallico, B.; Fanella, F.; Mauromicale, G.; Raccuia, S.A.; Foti, S. Possible alternative utilization of Cynara spp. II. Chemical characterization of their grain oil. Ind. Crop. Prod. 1999, 10, 229–237. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Rossi, M.; Russo, F.; Srirajaskanthan, R. Chapter 77—Table Olives: A Carrier for Delivering Probiotic Bacteria to Humans A2—Preedy, Victor R; Academic Press: New York, NY, USA, 2010; pp. 735–743. [Google Scholar]
- Wellenreuther, C.; Wolf, A. Innovative Feedstocks in Biodegradable Bio-Based Plastics: A Literature Review; Hamburgisches Welt-Wirtschafts-Archiv (HWWA): Hamburg, Germany, 2020. [Google Scholar]
- Koh, M.Y.; Tinia, T.I. A review of biodiesel production from Jatropha curcas L. oil. Renew. Sustain. Energy Rev. 2011, 15, 2240–2251. [Google Scholar] [CrossRef]
- Metzger, J.O.; Bornscheuer, U. Lipids as renewable resources: Current state of chemical and biotechnological conversion and diversification. Appl. Microbiol. Biotechnol. 2006, 71, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ierna, A.; Mauro, R.P.; Mauromicale, G. Biomass, grain and energy yield in Cynara cardunculus L. as affected by fertilization, genotype and harvest time. Biomass Bioenergy 2012, 36, 404–410. [Google Scholar] [CrossRef]
- Ottaiano, L.; Di Mola, I.; Impagliazzo, A.; Cozzolino, E.; Masucci, F.; Mori, M.; Fagnano, M. Yields and quality of biomasses and grain in Cynara cardunculus L. grown in southern Italy, as affected by genotype and environmental conditions. Ital. J. Agron. 2017, 12, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Pandino, G.; Lombardo, S.; Moglia, A.; Portis, E.; Lanteri, S.; Mauromicale, G. Leaf polyphenol profile and SSR-based fingerprinting of new segregant Cynara cardunculus genotypes. Front. Plant Sci. 2015, 5, 800. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curt, M.D.; Sánchez, G.; Fernández, J. The potential of Cynara cardunculus L. for seed oil production in a perennial cultivation system. Biomass Bioenergy 2002, 23, 33–46. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Piscioneri, I.; Sharma, N.; Melilli, M.G. Genetic variability in Cynara cardunculus L. domestic and wild types for grain oil production and fatty acids composition. Biomass Bioenergy 2011, 35, 3167–3173. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Cammareri, M.; Caretto, S.; Cavallaro, V.; Cominelli, E.; De Palma, M.; Docimo, T.; Giovinazzo, G.; Grandillo, S.; et al. Strategies to Modulate Specialized Metabolism in Mediterranean Crops: From Molecular Aspects to Field. Int. J. Mol. Sci. 2021, 22, 2887. [Google Scholar] [CrossRef]
- Nielsen, E.; Temporiti, M.E.E.; Cella, R. Improvement of phytochemical production by plant cells and organ culture and by genetic engineering. Plant Cell Rep. 2019, 38, 1199–1215. [Google Scholar] [CrossRef]
- Ferid, A.; Mohammed, A.; Khalivulla, S.I.; Korivi, M.; Abdul Razab, M.K.A. Plant Cell and Callus Cultures as an Alternative Source of Bioactive Compounds with Therapeutic Potential against Coronavirus Disease (COVID-19). IOP Conf. Ser. Earth Environ. Sci. 2020, 596, 012099. [Google Scholar] [CrossRef]
- Ramachandra Rao, S.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Anwar, M.; Chen, L.; Xiao, Y.; Wu, J.; Zeng, L.; Li, H.; Wu, Q.; Hu, Z. Recent Advanced Metabolic and Genetic Engineering of Phenylpropanoid Biosynthetic Pathways. Int. J. Mol. Sci. 2021, 22, 9544. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Fornalé, S.; Shi, X.; Chai, C.; Encina, A.; Irar, S.; Capellades, M.; Fuguet, E.; Torres, J.-L.; Rovira, P.; Puigdomènech, P.; et al. ZmMYB31 directly represses maize lignin genes and redirects the phenylpropanoid metabolic flux. Plant J. 2010, 64, 633–644. [Google Scholar] [CrossRef]
- Sonbol, F.M.; Fornalé, S.; Capellades, M.; Encina, A.; Touriño, S.; Torres, J.L.; Rovira, P.; Ruel, K.; Puigdomènech, P.; Rigau, J.; et al. The maize ZmMYB42 represses the phenylpropanoid pathway and affects the cell wall structure, composition and degradability in Arabidopsis thaliana. Plant Mol. Biol. 2009, 70, 283–296. [Google Scholar] [CrossRef]
- Ma, Q.H.; Wang, C.; Zhu, H.H. TaMYB4 cloned from wheat regulates lignin biosynthesis through negatively controlling the transcripts of both cinnamyl alcohol dehydrogenase and cinnamoyl-CoA reductase genes. Biochimie 2011, 93, 1179–1186. [Google Scholar] [CrossRef]
- Wei, Q.; Zhang, F.; Sun, F.; Luo, Q.; Wang, R.; Hu, R.; Chen, M.; Chang, J.; Yang, G.; He, G. A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants. Plant Sci. 2017, 265, 112–123. [Google Scholar] [CrossRef]
- Rao, X.; Chen, X.; Shen, H.; Ma, Q.; Li, G.; Tang, Y.; Pena, M.; York, W.; Frazier, T.P.; Lenaghan, S.; et al. Gene regulatory networks for lignin biosynthesis in switchgrass (Panicum virgatum). Plant Biotechnol. J. 2019, 17, 580–593. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, X.; Ran, L.; Li, C.; Fan, D.; Luo, K. PtoMYB156 is involved in negative regulation of phenylpropanoid metabolism and secondary cell wall biosynthesis during wood formation in poplar. Sci. Rep. 2017, 7, 41209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Takada, R.; Tobimatsu, Y.; Takeda, Y.; Suzuki, S.; Yamamura, M.; Osakabe, K.; Osakabe, Y.; Sakamoto, M.; Umezawa, T. OsMYB108 loss-of-function enriches p-coumaroylated and tricin lignin units in rice cell walls. Plant J. 2019, 98, 14290. [Google Scholar] [CrossRef]
- Fornalé, S.; Lopez, E.; Salazar-Henao, J.E.; Fernández-Nohales, P.; Rigau, J.; Caparros-Ruiz, D. AtMYB7, a new player in the regulation of UV-sunscreens in arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, Y.; Guo, L.; Gong, N.; Pang, Y.; Jiang, W.; Liu, Y.; Jiang, X.; Zhao, L.; Wang, Y.; et al. Functional characterization of tea (Camellia sinensis) MYB4a transcription factor using an integrative approach. Front. Plant Sci. 2017, 8, 943. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.C.; Wu, J.; Guan, M.L.; Zhao, C.H.; Geng, P.; Zhao, Q. Arabidopsis MYB4 plays dual roles in flavonoid biosynthesis. Plant J. 2020, 101, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Nuno de Sousa Sampaio, P.; Neto, H.; Poejo, P.; Serrazina, S.M.T.; Pais, M.S.S. Overexpression and characterization of cyprosin B in transformed suspension cells of Cynara cardunculus. Plant Cell Tissue Organ Cult. 2010, 101, 311–321. [Google Scholar] [CrossRef]
- Menin, B.; Moglia, A.; Comino, C.; Lanteri, S.; Van Herpen, T.W.J.M.; Beekwilder, J. In vitro callogenesis and agrobacterium-mediated transformation of globe artichoke. Acta Hortic. 2012, 961, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Maroufi, A.; Karimi, M.; Mehdikhanlou, K.; De Loose, M. Inulin chain length modification using a transgenic approach opening new perspectives for chicory. 3 Biotech 2018, 8, 349. [Google Scholar] [CrossRef]
- Ainasoja, M.M.; Pohjala, L.L.; Tammela, P.S.M.; Somervuo, P.J.; Vuorela, P.M.; Teeri, T.H. Comparison of transgenic Gerbera hybrida lines and traditional varieties shows no differences in cytotoxicity or metabolic fingerprints. Transgenic Res. 2008, 17, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Tamagnone, L.; Merida, A.; Parr, A.; Mackay, S.; Culianez-Macia, F.A.; Roberts, K.; Martin, C. The AmMYB308 and AmMYB330 transcription factors from antirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenic tobacco. Plant Cell 1998, 10, 135–154. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; He, X.; Poovaiah, C.R.; Wuddineh, W.A.; Ma, J.; Mann, D.G.J.; Wang, H.; Jackson, L.; Tang, Y.; Neal Stewart, J.; et al. Functional characterization of the switchgrass (Panicum virgatum) R2R3-MYB transcription factor PvMYB4 for improvement of lignocellulosic feedstocks. New Phytol. 2012, 193, 121–136. [Google Scholar] [CrossRef]
- Bonawitz, N.D.; Soltau, W.L.; Blatchley, M.R.; Powers, B.L.; Hurlock, A.K.; Seals, L.A.; Weng, J.K.; Stout, J.; Chapple, C. REF4 and RFR1, subunits of the transcriptional coregulatory complex mediator, are required for phenylpropanoid homeostasis in Arabidopsis. J. Biol. Chem. 2012, 287, 5434–5445. [Google Scholar] [CrossRef] [Green Version]
- Muro-Villanueva, F.; Mao, X.; Chapple, C. Linking phenylpropanoid metabolism, lignin deposition, and plant growth inhibition. Curr. Opin. Biotechnol. 2019, 56, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Valdiani, A.; Hansen, O.K.; Nielsen, U.B.; Johannsen, V.K.; Shariat, M.; Georgiev, M.I.; Omidvar, V.; Ebrahimi, M.; Tavakoli Dinanai, E.; Abiri, R. Bioreactor-based advances in plant tissue and cell culture: Challenges and prospects. Crit. Rev. Biotechnol. 2019, 39, 20–34. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Fevereiro, P.; Cabral, J.M.S.; Novais, J.M.; Salomé, M.; Pais, S. Callus and suspension cultures for biomass production of Cynara cardunculus (Compositae). Biotechnol. Lett. 1987, 9, 213–218. [Google Scholar] [CrossRef]
- Cordeiro, M.C.; Pais, M.S.; Brodelius, P.E. Cynara cardunculus subsp.flavescens (Cardoon): In Vitro Culture, and the Production of Cyprosins—Milk-Clotting Enzymes. Biotechnol. Agric. For. 1998, 41, 132–153. [Google Scholar] [CrossRef]
- van Damme, D.; de Rybel, B.; Gudesblat, G.; Demidov, D.; Grunewald, W.; de Smet, I.; Houben, A.; Beeckman, T.; Russinova, E. Arabidopsis α Aurora kinases function in formative cell division plane orientation. Plant Cell 2011, 23, 4013–4024. [Google Scholar] [CrossRef] [Green Version]
- Himanen, K.; Boccardi, T.M.; De Rycke, R.; Odeny, O.P.; Van Lijsebettens, M. Is HUB1 a hub for plant fitness? Plant Signal. Behav. 2012, 7, 1537–1540. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhang, Q.; Yan, X.; Zhang, D.; Zhou, Q.; Shen, Y.; Anupol, N.; Wang, X.; Bao, M.; Larkin, R.M.; et al. A conservative pathway for coordination of cell wall biosynthesis and cell cycle progression in plants. Plant J. 2021, 106, 630–648. [Google Scholar] [CrossRef]
- Mu, R.L.; Cao, Y.R.; Liu, Y.F.; Lei, G.; Zou, H.F.; Liao, Y.; Wang, H.W.; Zhang, W.K.; Ma, B.; Du, J.Z.; et al. An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis. Cell Res. 2009, 19, 1291–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownfield, L.; Hafidh, S.; Borg, M.; Sidorova, A.; Mori, T.; Twell, D. A plant germline-specific integrator of sperm specification and cell cycle progression. PLoS Genet. 2009, 5, e1000430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, S.; Coppens, F.; Lee, E.; Donner, T.J.; Xie, Z.; Van Isterdael, G.; Dhondt, S.; De Winter, F.; De Rybel, B.; Vuylsteke, M.; et al. Developmental regulation of CYCA2s contributes to tissue-specific proliferation in Arabidopsis. EMBO J. 2011, 30, 3430–3441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Lee, E.; Lucas, J.R.; Morohashi, K.; Li, D.; Murray, J.A.H.; Sack, F.D.; Grotewolda, E. Regulation of cell proliferation in the stomatal lineage by the Arabidopsis MYB FOUR LIPS via direct targeting of core cell cycle genes. Plant Cell 2010, 22, 2306–2321. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Li, L.; Lou, Y.; Zhu, C.; Li, X.; Gao, Z. A regulatory network driving shoot lignification in rapidly growing bamboo. Plant Physiol. 2021, 187, 900–916. [Google Scholar] [CrossRef] [PubMed]
- Sablowski, R.; Carnier Dornelas, M. Interplay between cell growth and cell cycle in plants. J. Exp. Bot. 2014, 65, 2703–2714. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.M.; Mota, T.R.; Grandis, A.; de Morais, G.R.; de Lucas, R.C.; Polizeli, M.L.T.M.; Marchiosi, R.; Buckeridge, M.S.; Ferrarese-Filho, O.; dos Santos, W.D. Lignin plays a key role in determining biomass recalcitrance in forage grasses. Renew. Energy 2020, 147, 2206–2217. [Google Scholar] [CrossRef]
- Wang, N.; Wang, K.Y.; Li, G.; Guo, W.; Liu, D. Expression and characterization of camel chymosin in Pichia pastoris. Protein Expr. Purif. 2015, 111, 75–81. [Google Scholar] [CrossRef]
- Trajtemberg, S.P.; Apóstolo, N.M.; Fernández, G. Calluses of Cynara cardunculus Var. Cardunculus cardoon (Asteraceae): Determination of cynarine and chlorogenic acid by automated high-performance capillary electrophoresis. Vitr. Cell. Dev. Biol. Plant 2006, 42, 534–537. [Google Scholar] [CrossRef]
- Correa, S.M.; Atehortúa, L. Lipid Profile of In Vitro Oil Produced through Cell Culture of Jatropha curcas. J. AOAC Int. 2012, 95, 1161–1169. [Google Scholar] [CrossRef]
- Hernández, M.L.; Sicardo, M.D.; Alfonso, M.; Martínez-Rivas, J.M. Transcriptional Regulation of Stearoyl-Acyl Carrier Protein Desaturase Genes in Response to Abiotic Stresses Leads to Changes in the Unsaturated Fatty Acids Composition of Olive Mesocarp. Front. Plant Sci. 2019, 10, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kummerow, F.A. The negative effects of hydrogenated trans fats and what to do about them. Atherosclerosis 2009, 205, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharm. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Lee, K.-R.; Jeon, I.; Yu, H.; Kim, S.-G.; Kim, H.-S.; Ahn, S.-J.; Lee, J.; Lee, S.-K.; Kim, H.U. Increasing Monounsaturated Fatty Acid Contents in Hexaploid Camelina sativa Seed Oil by FAD2 Gene Knockout Using CRISPR-Cas9. Front. Plant Sci. 2021, 0, 1279. [Google Scholar] [CrossRef]
- Damude, H.G.; Kinney, A.J. Enhancing Plant Seed Oils for Human Nutrition. Plant Physiol. 2008, 147, 962. [Google Scholar] [CrossRef] [Green Version]
- Beló, A.; Zheng, P.; Luck, S.; Shen, B.; Meyer, D.J.; Li, B.; Tingey, S.; Rafalski, A. Whole genome scan detects an allelic variant of fad2 associated with increased oleic acid levels in maize. Mol. Genet. Genom. 2008, 279, 1–10. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.D.; Zhang, Y.Y.; Yang, J.; Qi, G.X.; Guo, D.Q.; Xing, G.J.; Yao, Y.; Xu, W.J.; Li, H.Y.; et al. Changes in oleic acid content of transgenic soybeans by antisense RNA mediated posttranscriptional gene silencing. Int. J. Genom. 2014, 2014, 921950. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Jung, S.; Moore, K.; Powell, G.; Ainsworth, C.; Abbott, A. High-oleate peanut mutants result from a MITE insertion into the FAD2 gene. Theor. Appl. Genet. 2004, 108, 1492–1502. [Google Scholar] [CrossRef]
- Cheng, T.; Zhao, P.; Ren, Y.; Zou, J.; Sun, M.-X. AtMIF1 increases seed oil content by attenuating GL2 inhibition. New Phytol. 2021, 229, 2152–2162. [Google Scholar] [CrossRef]
- Gou, M.; Hou, G.; Yang, H.; Zhang, X.; Cai, Y.; Kai, G.; Liu, C.-J. The MYB107 Transcription Factor Positively Regulates Suberin Biosynthesis. Plant Physiol. 2017, 173, 1045. [Google Scholar] [CrossRef] [Green Version]
- Meï, C.; Michaud, M.; Cussac, M.; Albrieux, C.; Gros, V.; Maréchal, E.; Block, M.A.; Jouhet, J.; Rébeillé, F. Levels of polyunsaturated fatty acids correlate with growth rate in plant cell cultures. Sci. Rep. 2015, 5, 15207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidabadi, S.S.; Mohan Jain, S. Cellular, molecular, and physiological aspects of in vitro plant regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Comino, C.; Moglia, A.; Repetto, A.; Tavazza, R. Globe Artichoke Tissue Culture and Its Biotechnological Application. Globe Artichoke Genome 2019, 41–64. [Google Scholar] [CrossRef]
- Blom, T.J.M.; Kreis, W.; van Iren, F.; Libbenga, K.R. A non-invasive method for the routine-estimation of fresh weight of cells grown in batch suspension cultures. Plant Cell Rep. 1992, 11, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAYTM vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Haas, J.H.; Ream, W.; Manulis, S. Universal PCR Primers for Detection of Phytopathogenic. Microbiology 1995, 61, 2879–2884. [Google Scholar]
- Gabotti, D.; Locatelli, F.; Cusano, E.; Baldoni, E.; Genga, A.; Pucci, L.; Consonni, R.; Mattana, M. Cell suspensions of Cannabis sativa (var. futura): Effect of elicitation on metabolite content and antioxidant activity. Molecules 2019, 24, 4056. [Google Scholar] [CrossRef] [Green Version]
- Docimo, T.; Consonni, R.; Coraggio, I.; Mattana, M. Early phenylpropanoid biosynthetic steps in Cannabis sativa: Link between genes and metabolites. Int. J. Mol. Sci. 2013, 14, 13626–13644. [Google Scholar] [CrossRef] [Green Version]
- Acker, R.; Vanholme, R.; Piens, K.; Boerjan, W. Saccharification Protocol for Small-scale Lignocellulosic Biomass Samples to Test Processing of Cellulose into Glucose. Bio-Protocol 2016, 6, e1701. [Google Scholar] [CrossRef] [Green Version]
- Kostić, M.D.; Joković, N.M.; Stamenković, O.S.; Rajković, K.M.; Milić, P.S.; Veljković, V.B. Optimization of hempseed oil extraction by n-hexane. Ind. Crop. Prod. 2013, 48, 133–143. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnante, G.; D’Amore, R.; Blanco, E.; Pierri, C.L.; de Palma, M.; Luo, J.; Tucci, M.; Martin, C. Novel hydroxycinnamoyl-coenzyme a quinate transferase genes from artichoke are involved in the synthesis of chlorogenic acid. Plant Physiol. 2010, 153, 1224–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Rau, A.; Gallopin, M.; Celeux, G.; Jaffrézic, F. Data-based filtering for replicated high-throughput transcriptome sequencing experiments. Bioinformatics 2013, 29, 2146–2152. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, I.; Lugmayr, A.; Siira, S.J.; Rackham, O.; Filipovska, A. CirGO: An alternative circular way of visualising gene ontology terms. BMC Bioinform. 2019, 20, 84. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | L1 | L18 | L24 | ||
|---|---|---|---|---|---|
| Polyphenols (µg/g DW) | 5-iFQA | 21.50 ± 1.33a | 15.41 ± 0.43b | 0.11 ± 0.04c | 4.26 ± 0.12d |
| 1,5-DiCQA (cynarin) | 3345.79 ± 112.23a | 532.70 ± 11.55b | 2.50 ± 0.81c | 352.98 ± 21.71d | |
| 3,4-DiCQA | 3152.41 ± 26.67a | 443.88 ± 12.43b | 103.58 ± 8.23c | 1615.65 ± 34.56d | |
| 5-FQA | 98.87 ± 11.34a | 5.56 ± 0.43b | 0.83 ± 0.011c | 76.44 ± 11.34d | |
| 3-FQA | 3.09 ± 0.55a | 2.71 ± 0.91b | 0.10 ± 0.01c | 10.20 ± 0.23d | |
| 3-CQA (CGA) | 319.38 ± 22.45a | 424.66 ± 11.32b | 97.28 ± 3.56c | 513.48 ± 13.57d | |
| p-coumaric acid | 3.50 ± 0.02a | 3.75 ± 0.91a | 1.50 ± 0.01b | 3.85 ± 0.02a | |
| Quercetin | 3.35 ± 0.02a | 4.30 ± 0.02b | 0.55 ± 0.01c | 4.35 ± 0.02b | |
| Quercetin-glucoside | 4.00 ± 0.65a | 5.25 ± 1.23b | 1.25 ± 0.03c | 5.40 ± 0.34b | |
| Kaempferol | 3.55 ± 0.65a | 4.30 ± 0.91b | 1.05 ± 0.03c | 4.55 ± 0.83d | |
| Kaempferol-3-O-glucoside | 1.15 ± 0.02a | 1.50 ± 0.03b | 0.40 ± 0.04c | 1.60 ± 0.04b | |
| Naringin | 1.20 ± 0.01a | 1.75 ± 0.01b | 0.41 ± 0.03c | 1.800.03b | |
| Luteolin | 1.50 ± 0.03a | 2.35 ± 0.02b | 0.60 ± 0.04c | 2.55 ± 0.04d | |
| Myricetin | 2.55 ± 0.34a | 2.96 ± 0.32b | 0.73 ± 0.11c | 3.04 ± 0.12b | |
| Apigenin | 0.20 ± 0.03a | 0.25 ± 0.02b | 0.01 ± 0.003c | 0.30 ± 0.01d | |
| Total polyphenols | 6962.05a | 1451.33b | 210.92c | 2600.46d | |
| Antiox. activity (TEAC) | DPPH | 83.7 ± 0.61a | 22.17 ± 0.42b | 16.96 ± 0.63c | 35.43 ± 5.82d |
| ABTS | 71.33 ± 0.20a | 19.16 ± 0.07b | 12.54 ± 0.26c | 23.61 ± 0.22d | |
| FRAP | 66.81 ± 0.98a | 30.84 ± 0.21b | 11.09 ± 0.08c | 27.90 ± 0.54d | |
| Oil % | 11.38 | 8.24 | 10.35 | 7.25 | |
| Fatty acids (%) | Palmitic (C16:0) | 22.29 ± 1.12ab | 21.71 ± 0.58a | 24.06 ± 0.35b | 19.46 ± 0.38c |
| Stearic (C18:0) | 2.85 ± 0.31ab | 3.14 ± 0.54ab | 2.29 ± 0.07a | 3.73 ± 0.26b | |
| Oleic (C18:1) | 3.43 ± 1.91a | 10.30 ± 0.56b | 3.49 ± 0.27a | 12.35 ± 0.16c | |
| Linoleic (C18:2) | 19.38 ± 3.74a | 45.82 ± 0.85b | 41.04 ± 0.61c | 41.34 ± 0.39c | |
| Linolenic (C18:3) | 42.60 ± 6.34a | 10.16 ± 0.38b | 20.73 ± 0.74c | 14.46 ± 0.06d | |
| Arachidic (C20:0) | 0.80 ± 0.08a | 0.88 ± 0.01a | 0.84 ± 0.05a | 1.04 ± 0.05b | |
| Lignoceric (C24:0) | 2.42 ± 0.30a | 3.26 ± 1.59a | 2.41 ± 0.58a | 2.40 ± 0.68a | |
| Nervonic (C24:1) | 1.23 ± 0.14a | 0.18 ± 0.03b | 0.52 ± 0.47ab | 0.66 ± 0.12ab | |
| Total SFA% | 30.54 ± 0.78ab | 30.29 ± 0.76a | 32.07 ± 0.43b | 28.00 ± 0.10c | |
| Total MUFA % | 4.65 ± 1.777a | 10.48 ± 0.30b | 4.02 ± 0.21a | 13.01 ± 0.28c | |
| Total PUFA % | 61.98 ± 2.88a | 55.98 ± 0.80b | 61.77 ± 0.41a | 55.81 ± 0.42b | |
| Others % | 2.83 | 3.25 | 2.14 | 3.18 |
| Number of Core-DEGs | ||
|---|---|---|
| TF Family | Downregulated | Upregulated |
| AP2 | 1 | 3 |
| bHLH | 7 | 11 |
| bZIP | 5 | 4 |
| C2H2 | 1 | 6 |
| C3H | 2 | - |
| Dof | 3 | 1 |
| ERF | 2 | 6 |
| FAR1 | 1 | 2 |
| G2-like | 1 | 3 |
| GRAS | 3 | 7 |
| HB-other | 2 | - |
| HSF | 1 | 1 |
| LBD | 3 | 4 |
| MIKC_MADS | 4 | 2 |
| M-type MADS | 2 | 2 |
| MYB | 1 | 4 |
| MYB_related | 6 | 5 |
| NAC | 14 | 3 |
| NF-YA | 1 | - |
| Nin-like | 1 | - |
| S1Fa-like | 1 | - |
| TALE | 2 | 3 |
| TCP | 1 | - |
| Trihelix | 1 | 1 |
| WOX | 2 | - |
| WRKY | 9 | 6 |
| ZF-HD | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paolo, D.; Locatelli, F.; Cominelli, E.; Pirona, R.; Pozzo, S.; Graziani, G.; Ritieni, A.; De Palma, M.; Docimo, T.; Tucci, M.; et al. Towards a Cardoon (Cynara cardunculus var. altilis)-Based Biorefinery: A Case Study of Improved Cell Cultures via Genetic Modulation of the Phenylpropanoid Pathway. Int. J. Mol. Sci. 2021, 22, 11978. https://doi.org/10.3390/ijms222111978
Paolo D, Locatelli F, Cominelli E, Pirona R, Pozzo S, Graziani G, Ritieni A, De Palma M, Docimo T, Tucci M, et al. Towards a Cardoon (Cynara cardunculus var. altilis)-Based Biorefinery: A Case Study of Improved Cell Cultures via Genetic Modulation of the Phenylpropanoid Pathway. International Journal of Molecular Sciences. 2021; 22(21):11978. https://doi.org/10.3390/ijms222111978
Chicago/Turabian StylePaolo, Dario, Franca Locatelli, Eleonora Cominelli, Raul Pirona, Sara Pozzo, Giulia Graziani, Alberto Ritieni, Monica De Palma, Teresa Docimo, Marina Tucci, and et al. 2021. "Towards a Cardoon (Cynara cardunculus var. altilis)-Based Biorefinery: A Case Study of Improved Cell Cultures via Genetic Modulation of the Phenylpropanoid Pathway" International Journal of Molecular Sciences 22, no. 21: 11978. https://doi.org/10.3390/ijms222111978