
Overexpression of ZNT1 and NRAMP4 from the Ni Hyperaccumulator Noccaea caerulescens Population Monte Prinzera in Arabidopsis thaliana Perturbs Fe, Mn, and Ni Accumulation

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. The Expression of NRAMP4 and ZNT1 in Ni hyperaccumulator N. caerulescens MP
2.2. Expression of ZNT1/ZIP4 and NRAMP4 from N. caerulescens and A. thaliana Has Contrasting Effects in Yeast
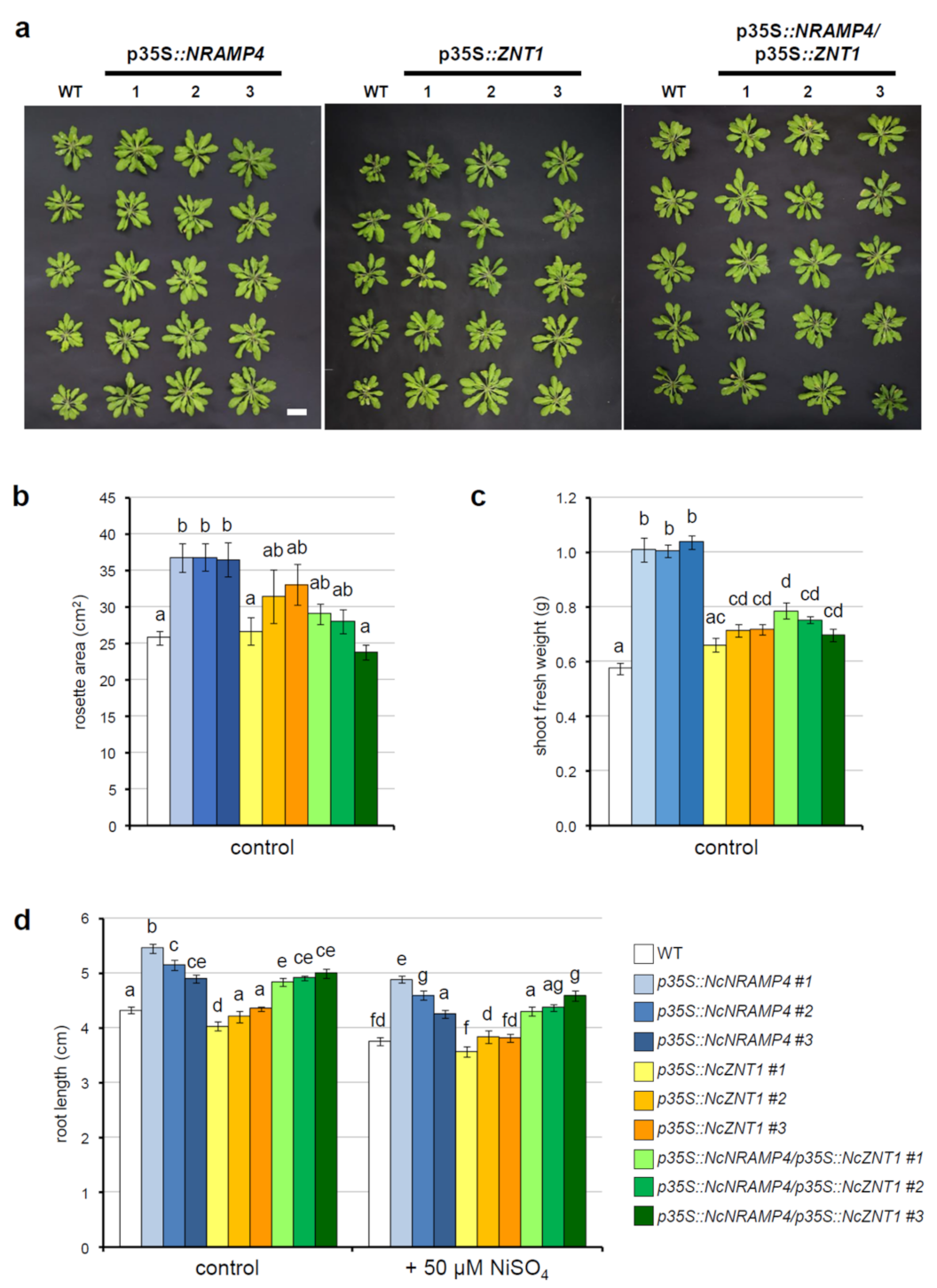
2.3. Expression of NcNRAMP4 and NcZNT1 in A. thaliana Alters Plant Growth and Metal Accumulation
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. ZNT1 and NRAMP4 Expression Analysis in N. caerulescens and T. arvense
4.3. Isolation of NcNRAMP4, NcZNT1, AtNRAMP4 and AtZIP4 Coding Sequences, Plasmid DNA Constructs Preparation and Yeast Transformation
4.4. Analysis of Ni Tolerance and Accumulation in Yeast
4.5. Plasmid DNA Constructs Preparation and A. thaliana Transformation
4.6. Analysis of A. thaliana Transgenic Lines
4.7. Metal Quantification in Yeast and A. thaliana Transgenic Lines
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krämer, U. Metal Hyperaccumulation in Plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Manara, A.; Fasani, E.; Furini, A.; DalCorso, G. Evolution of the metal hyperaccumulation and hypertolerance traits. Plant Cell Environ. 2020, 43, 2969–2986. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Tanikawa, R.; Ishida, S.; Yoshida, J.; Mizuno, T.; Nakanishi, H.; Furuta, N. Elevated Expression of Vacuolar Nickel Transporter Gene IREG2 Is Associated with Reduced Root-to-Shoot Nickel Translocation in Noccaea japonica. Front. Plant Sci. 2020, 11, 610. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ji, X.; Luo, X. Visualizing Hotspots and Future Trends in Phytomining Research Through Scientometrics. Sustainability 2020, 12, 4593. [Google Scholar] [CrossRef]
- Novo, L.A.B.; Castro, P.M.L.; Alvarenga, P.; da Silva, E.F. Phytomining of Rare and Valuable Metals. In Phytoremediation—Management of Environmental Contaminants; 2017; Volume 5, pp. 469–486. [Google Scholar] [CrossRef]
- Kozhevnikova, A.; Seregin, I.V.; Schat, H. Accumulation of Nickel by Excluder Thlaspi arvense and Hyperaccumulator Noccaea caerulescens upon Short-Term and Long-Term Exposure. Russ. J. Plant Physiol. 2020, 67, 303–311. [Google Scholar] [CrossRef]
- Van Der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2012, 362, 319–334. [Google Scholar] [CrossRef]
- Assunção, A.G.L.; Bookum, W.M.; Nelissen, H.J.M.; Vooijs, R.; Schat, H.; Ernst, W.H.O. Differential metal-specific tolerance and accumulation patterns among Thlaspi caerulescens populations originating from different soil types. New Phytol. 2003, 159, 411–419. [Google Scholar] [CrossRef]
- Kozhevnikova, A.D.; Seregin, I.V.; Aarts, M.G.; Schat, H. Intra-specific variation in zinc, cadmium and nickel hyper-tolerance and hyperaccumulation capacities in Noccaea caerulescens. Plant Soil 2020, 452, 479–498. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Physiol. Plant. 2020. [Google Scholar] [CrossRef]
- Van de Mortel, J.E.; Almar Villanueva, L.; Schat, H.; Kwekkeboom, J.; Coughlan, S.; Moerland, P.D.; Aarts, M.G. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2006, 142, 1127–1147. [Google Scholar] [CrossRef] [Green Version]
- Van de Mortel, J.E.; Schat, H.; Moerland, P.D.; Van Themaat, E.V.L.; Van Der Ent, S.J.O.E.R.D.; Blankestijn, H.; Aarts, M.G. Expression differences for genes involved in lignin, glutathione and sulphate metabolism in response to cadmium in Arabidopsis thaliana and the related Zn/Cd-hyperaccumulator Thlaspi caerulescens. Plant Cell Environ. 2008, 31, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Halimaa, P.; Lin, Y.-F.; Ahonen, V.H.; Blande, D.; Clemens, S.; Gyenesei, A.; Häikiö, E.; Kärenlampi, S.O.; Laiho, A.; Aarts, M.G.M.; et al. Gene Expression Differences between Noccaea caerulescens Ecotypes Help to Identify Candidate Genes for Metal Phytoremediation. Environ. Sci. Technol. 2014, 48, 3344–3353. [Google Scholar] [CrossRef] [PubMed]
- Foroughi, S.; Baker, A.J.M.; Roessner, U.; Johnson, A.A.T.; Bacic, A.; Callahan, D.L. Hyperaccumulation of zinc by Noccaea caerulescens results in a cascade of stress responses and changes in the elemental profile. Metallomics 2014, 6, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Pence, N.S.; Larsen, P.B.; Ebbs, S.; Letham, D.L.D.; Lasat, M.M.; Garvin, D.F.; Eide, D.; Kochian, L. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA 2000, 97, 4956–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, M.; Craft, E.; Yamaji, N.; Koyama, E.; Ma, J.F.; Kochian, L. Characterization of the high affinity Zn transporter from Noccaea caerulescens, NcZNT1, and dissection of its promoter for its role in Zn uptake and hyperaccumulation. New Phytol. 2012, 195, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Oomen, R.J.F.J.; Wu, J.; Lelievre, F.; Blanchet, S.; Richaud, P.; Barbier-Brygoo, H.; Aarts, M.G.; Thomine, S. Functional char-acterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens. New Phytol. 2009, 181, 637–650. [Google Scholar] [CrossRef]
- Visioli, G.; Gullì, M.; Marmiroli, N. Noccaea caerulescens populations adapted to growth in metalliferous and non-metalliferous soils. Environ. Exp. Bot 2014, 105, 10–17. [Google Scholar] [CrossRef]
- Mizuno, T.; Usui, K.; Horie, K.; Nosaka, S.; Mizuno, N.; Obata, H. Cloning of three ZIP/Nramp transporter genes from a Ni hyperaccumulator plant Thlaspi japonicum and their Ni2+-transport abilities. Plant Physiol. Biochem. 2005, 43, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Vincenzi, S.; Marmiroli, M.; Marmiroli, N. Correlation between phenotype and proteome in the Ni hyperaccumulator Noccaea caerulescens subsp. caerulescens. Environ. Exp. Bot. 2012, 77, 156–164. [Google Scholar] [CrossRef]
- Visioli, G.; Menta, C.; Gardi, C.; Conti, F.D. Metal toxicity and biodiversity in serpentine soils: Application of bioassay tests and microarthropod index. Chemosphere 2013, 90, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Maestri, E.; Pirondini, A.; Visioli, G.; Marmiroli, N. Trade-off between genetic variation and ecological adaptation of metallicolous and non-metallicolous Noccaea and Thlaspi species. Environ. Exp. Bot. 2013, 96, 1–10. [Google Scholar] [CrossRef]
- Seregin, I.; Kozhevnikova, A. Physiological role of nickel and its toxic effects on higher plants. Russ. J. Plant Physiol. 2006, 53, 257–277. [Google Scholar] [CrossRef]
- Jaffré, T.; Reeves, R.D.; Baker, A.J.M.; Schat, H.; van der Ent, A. The discovery of nickel hyperaccumulation in the New Caledonian tree Pycnandra acuminata 40 years on: An introduction to a Virtual Issue. New Phytol. 2018, 218, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Broadhurst, C.L.; Tappero, R.V.; Maugel, T.K.; Erbe, E.F.; Sparks, D.L.; Chaney, R.L. Interaction of nickel and manganese in accumulation and localization in leaves of the Ni hyperaccumulators Alyssum murale and Alyssum corsicum. Plant Soil 2008, 314, 35–48. [Google Scholar] [CrossRef]
- Nishida, S.; Tsuzuki, C.; Kato, A.; Aisu, A.; Yoshida, J.; Mizuno, T. AtIRT1, the Primary Iron Uptake Transporter in the Root, Mediates Excess Nickel Accumulation in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1433–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruotolo, R.; Marchini, G.; Ottonello, S. Membrane transporters and protein traffic networks differentially affecting metal tolerance: A genomic phenotyping study in yeast. Genome Biol. 2008, 9, R67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanquar, V.; Lelièvre, F.; Barbier-Brygoo, H.; Thomine, S. Regulation and function of AtNRAMP4 metal transporter protein. Soil Sci. Plant Nutr. 2004, 50, 1141–1150. [Google Scholar] [CrossRef]
- Thomine, S.; Wang, R.; Ward, J.; Crawford, N.M.; Schroeder, J.I. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA 2000, 97, 4991–4996. [Google Scholar] [CrossRef] [Green Version]
- Pottier, M.; Oomen, R.; Picco, C.; Giraudat, J.; Scholz-Starke, J.; Richaud, P.; Carpaneto, A.; Thomine, S. Identification of mutations allowing Natural Resistance Associated Macrophage Proteins (NRAMP) to discriminate against cadmium. Plant J. 2015, 83, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-F.; Hassan, Z.; Talukdar, S.; Schat, H.; Aarts, M.G.M. Expression of the ZNT1 Zinc Transporter from the Metal Hyperaccumulator Noccaea caerulescens Confers Enhanced Zinc and Cadmium Tolerance and Accumulation to Arabidopsis thaliana. PLoS ONE 2016, 11, e0149750. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.H.; Marschner, H. The Mineral Nutrition of Higher Plants. J. Ecol. 1988, 76, 1250. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, R.; Ghaderian, S.M.; Krämer, U. Interference of nickel with copper and iron homeostasis contributes to metal toxicity symptoms in the nickel hyperaccumulator plant Alyssum inflatum. New Phytol. 2009, 184, 566–580. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Sharma, C.P. Effect of heavy metals Co2+, Ni2+ and Cd2+ on growth and metabolism of cabbage. Plant Sci. 2002, 163, 753–758. [Google Scholar] [CrossRef]
- Nishida, S.; Aisu, A.; Mizuno, T. Induction ofIRT1by the nickel-induced iron-deficient response in Arabidopsis. Plant Signal. Behav. 2012, 7, 329–331. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water Culture Method for Growing Plants without Soil; Circular no. 347; University of California College of Agriculture: Berkeley, CA, USA, 1938. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2–DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Agatep, R.; Kirkpatrick, R.D.; Parchaliuk, D.L.; Woods, R.A.; Gietz, R.D. Transformation of Saccharomyces cerevisiae by the lithium acetate/single-stranded carrier DNA/polyethylene glycol protocol. Tech. Tips Online 1998, 3, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Sending, L.N.; Filippone, M.P.; Orce, I.G.; Rigano, L.; Enrique, R.; Peña, L.; Vojnov, A.A.; Marano, M.R.; Castagnaro, A.P. Transient expression of pepper Bs2 gene in Citrus limona an approach to evaluate its utility for management of citrus canker disease. Plant Pathol 2012, 61, 648–657. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullì, M.; Marchi, L.; Fragni, R.; Buschini, A.; Visioli, G. Epigenetic modifications preserve the hyperaccumulator Noccaea caerulescens from Ni genotoxicity. Environ. Mol. Mutagen. 2018, 59, 464–475. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | +20 µM NiSO4 | |||||
|---|---|---|---|---|---|---|
| Genotype | Fe | Mn | Ni | Fe | Mn | Ni |
| WT | 0.09 ± 0.01 a | 0.17 ± 0.01 a | - | 0.02 ± 0.00 a | 0.53 ± 0.07 ab | 0.02 ± 0.00 a |
| p35S::NRAMP4 | 0.12 ± 0.00 a | 0.23 ± 0.01 ab | - | 0.02 ± 0.00 a | 0.42 ± 0.07 a | 0.02 ± 0.00 a |
| p35S::ZNT1 | 0.11 ± 0.00 a | 0.24 ± 0.00 b | - | 0.02 ± 0.00 a | 0.85 ± 0.09 b | 0.03 ± 0.00 a |
| p35S::NRAMP4/p35S::ZNT1 | 0.21 ± 0.01 b | 0.20 ± 0.01 a | - | 0.02 ± 0.00 a | 0.65 ± 0.11 ab | 0.02 ± 0.00 a |
| A. thaliana transformation | ||
|---|---|---|
| NcNRAMP4 | #F | GGGGACAAGTTTGTACAAAAAGCAGGCTATGTCGGAGACGGAGAGAGA |
| #R | GGGGACCACTTTGTACAAGAAAGCTGGGTCTAATTGCAAGGAGTGTACGT | |
| NcZNT1 | #F | GGATCCATGATCATCGCCGATCTTCTTTG |
| #R | CTCGAGCTAAGCCCAAATGGCGAGTG | |
| Yeast complementation | ||
| NcNRAMP4 | #F | GGATCCATGTCGGGAGACTGATAGAGAG |
| #R | GAATTCCTAATTGCAAGGAGTGTACGT | |
| NcZNT1 | #F | GGATCCATGATCATCGCCGATCTTCTT |
| #R | GAATTCCTAAGCCCCAAATGGCGAGTG | |
| AtNRMAP4 | #F | GGATCCATGTCGGAGACTGATAGAGAGCG |
| #R | GAATTCCTCACTCATCATCCCTCTGTGG | |
| AtZIP4 | #F | GGATCCATGGCTTCTTCTACCACTAAA |
| #R | CTCGAGCTAAGCCCAAATGGCGAGAGCA | |
| Expression analysis by Real Time—RT PCR | ||
| Nc-Ta NRAMP4 | #F_N4RT | TTCCCGATACTCTACATATGG |
| #R_N4RT | CCATCGCATGTACCATGAGC | |
| Nc-Ta ZNT1 | #F_ZNTRT | TCAACTCGCATAGCCCTGG |
| #R_ZNTRT | AGCCTCACATTACAACTCATC | |
| ACT2/8 | #F_ACT | AACATTGTGCTCAGTGGTGG |
| #R_ACT | GACCTTAATCTTCATGCTGCT | |
| Ubi10 | #F_UBI10 | GGACAAGGAAGGTATTCCTC |
| #R_UBI10 | CTCCTTCTGGATGTTGTAGTC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasani, E.; DalCorso, G.; Zorzi, G.; Agrimonti, C.; Fragni, R.; Visioli, G.; Furini, A. Overexpression of ZNT1 and NRAMP4 from the Ni Hyperaccumulator Noccaea caerulescens Population Monte Prinzera in Arabidopsis thaliana Perturbs Fe, Mn, and Ni Accumulation. Int. J. Mol. Sci. 2021, 22, 11896. https://doi.org/10.3390/ijms222111896
Fasani E, DalCorso G, Zorzi G, Agrimonti C, Fragni R, Visioli G, Furini A. Overexpression of ZNT1 and NRAMP4 from the Ni Hyperaccumulator Noccaea caerulescens Population Monte Prinzera in Arabidopsis thaliana Perturbs Fe, Mn, and Ni Accumulation. International Journal of Molecular Sciences. 2021; 22(21):11896. https://doi.org/10.3390/ijms222111896
Chicago/Turabian StyleFasani, Elisa, Giovanni DalCorso, Gianluca Zorzi, Caterina Agrimonti, Rosaria Fragni, Giovanna Visioli, and Antonella Furini. 2021. "Overexpression of ZNT1 and NRAMP4 from the Ni Hyperaccumulator Noccaea caerulescens Population Monte Prinzera in Arabidopsis thaliana Perturbs Fe, Mn, and Ni Accumulation" International Journal of Molecular Sciences 22, no. 21: 11896. https://doi.org/10.3390/ijms222111896