
Extended Cheese Whey Fermentation Produces a Novel Casein-Derived Antibacterial Polypeptide That Also Inhibits Gelatinases MMP-2 and MMP-9

 , , ,
, , ,
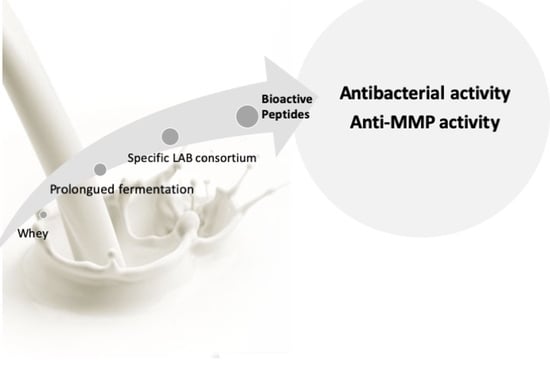
Abstract
:
1. Introduction
2. Results and Discussion
2.1. HPLC Peptide Profile throughout the 6-Day Fermentation
2.2. HPLC Peak 2 Presents High Antibacterial Activity
2.3. Peak 2 Is A Polypeptide Derived from Beta-Casein Proteolysis
2.4. Peak 2 Polypeptide Inhibits Other Pathogenic Bacteria
2.5. Peak 2 Polypeptide Inhibits Gelatinases MMP-2 and -9 and Reduces Colon Cancer Migration
3. Materials and Methods
3.1. LAB and Media
3.2. Whey Fermentation Conditions
3.3. Evaluation of Fermentation
3.4. Protein Quantification
3.5. Peptide Isolation through Ultrafiltration
3.6. Peptide Separation through HPLC
3.7. Antibacterial Activity of the Peptide Fractions
3.8. Mass Spectrometry (MS) Analysis
3.9. Polypeptide Identification
3.10. In Vitro Colon Cancer Cell Assays
3.10.1. HT29 Cell Cultures
3.10.2. Wound Healing Assay
3.10.3. MMP-9 and MMP-2 Catalytic Activity
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Recio, I.; Amigo, L. β-Lactoglobulin as source of bioactive peptides. Amino Acids 2008, 35, 257–265. [Google Scholar] [CrossRef]
- Madureira, A.; Pereira, C.; Gomes, A.; Pintado, M.; Malcata, F. Bovine whey proteins—Overview on their main biological properties. Food Res. Int. 2007, 40, 1197–1210. [Google Scholar] [CrossRef]
- Lopez-Exposito, I.L.; Recio, I. Protective effect of milk peptides: Antibacterial and antitumor properties. In Bioactive Components of Milk, Advances in Experimental Medicine and Biology; Bosze, Z., Ed.; Springer Science and Business Media: New York, NY, USA, 2008; Volume 606, pp. 271–293. [Google Scholar] [CrossRef]
- Korhonen, H. Milk-derived bioactive peptides: From science to applications. J. Funct. Foods 2009, 1, 177–187. [Google Scholar] [CrossRef]
- Tavares, T.G.; Malcata, F.X. Whey Proteins as Source of Bioactive Peptides Against Hypertension. In Bioactive Food Peptides in Health and Disease; Hernández-Ledesma, B., Hsieh, C., Eds.; Chapter 4; InTech: London, UK, 2013; pp. 75–114. Available online: https://www.intechopen.com/chapters/42430 (accessed on 13 April 2021).
- Rizzello, C.G.; Losito, I.; Gobbetti, M.; Carbonara, T.; De Bari, M.D.; Zambonii, P.G. Antibacterial Activities of Peptides from the Water-Soluble Extracts of Italian Cheese Varieties. J. Dairy Sci. 2005, 88, 2348–2360. [Google Scholar] [CrossRef] [Green Version]
- Jabbari, M.; Rahele, H.; Farshid, K.; Sahar, J. Antimicrobial Peptides from Milk Proteins: A Prospectus. Ann. Biol. Res. 2012, 3, 5313–5318. Available online: www.scholarsresearchlibrary.com (accessed on 5 July 2021).
- Santos, M.I.S.; Martins, S.R.; Pedroso, L.; Sousa, I.; Ferreira, M.A.S.S. Potential bioactivity of whey fermented extract as sanitizer of organic grown lettuce. Food Control. 2015, 50, 477–481. [Google Scholar] [CrossRef]
- Santos, M.I.S.; Lima, A.I.; Monteiro, S.A.V.S.; Ferreira, R.M.S.B.; Pedroso, L.; Sousa, I.; Ferreira, M.A.S.S. Preliminary study on the effect of fermented cheese whey on Listeria monocytogenes, Escherichia coli O157:H7 and Salmonella Goldcoast populations inoculated onto fresh organic lettuce. Foodborne Pathog. Dis. 2016, 13, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.I.S.; Fradinho, P.; Martins, S.; Lima, A.I.; Ferreira, R.M.S.B.; Pedroso, L.; Ferreira, M.A.S.S.; Sousa, I. A novel way for whey: Cheese whey fermentation produces an effective and environmentally safe alternative to chlorine. Appl. Sci. 2019, 9, 2800. [Google Scholar] [CrossRef] [Green Version]
- Baugh, M.D.; Perry, M.J.; Hollander, A.P.; Davies, D.R.; Cross, S.S.; Lobo, A.J.; Taylor, C.J.; Evans, G.S. Matrix metalloproteinase levels are elevated in inflammatory bowel disease. Gastroenterology 1999, 117, 814–822. [Google Scholar] [CrossRef]
- Lee, K.; Vansaun, M.N.; Shim, J.H.; Matrisian, L.M.; Gorden, D.L. Increased metastases are associated with inflammation and matrix metalloproteinase-9 activity at incision sites in a murine model of peritoneal dissemination of colorectal cancer. J. Surg. Res. 2013, 180, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Markle, B.; May, E.J.; Majumdar, A.P.N. Do nutraceutics play a role in the prevention and treatment of colorectal cancer? Cancer Metastasis Rev. 2010, 29, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Nagase, H. Progress in matrix metalloproteinase research. Mol. Asp. Med. 2009, 29, 290–308. [Google Scholar] [CrossRef] [Green Version]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Abdelazeem, M.A.; El-Sayed, M. The pattern of CD44 and matrix metalloproteinase 9 expression is a useful predictor of ulcerative colitis–associated dysplasia and neoplasia. Ann. Diagn. Pathol. 2015, 19, 369–374. [Google Scholar] [CrossRef]
- Kofla-Dłubacz, A.; Matusiewicz, M.; Krzesiek, E.; Noga, L.; Iwańczak, B. Metalloproteinase-3 and -9 as novel markers in the evaluation of ulcerative colitis activity in children. Adv. Clin. Exp. Med. 2014, 23, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Siloşi, I.; Boldeanu, M.V.; Mogoantă, S.; Ghiluşi, M.; Cojocaru, M.; Biciuşcă, V.; Cojocaru, I.M.; Avrămescu, C.S.; Gheonea, D.I.; Siloşi, C.A.; et al. Matrix metalloproteinases (MMP-3 and MMP-9) implication in the pathogenesis of inflammatory bowel disease (IBD). Rom. J. Morphol. Embryol. 2014, 55, 1317–1324. [Google Scholar]
- Matusiewicz, M.; Neubauer, K.; Mierzchala-Pasierb, M.; Gamian, A.; Krzystek-Korpacka, M. Matrix metalloproteinase-9: Its interplay with angiogenic factors in inflammatory bowel diseases. Dis. Markers 2014, 2014, 643645. [Google Scholar] [CrossRef]
- Spoerlein, C.; Mahal, K.; Schmidt, H.; Schobert, R. Effects of chrysin, apigenin, genistein and their homoleptic copper(II) complexes on the growth and metastatic potential of cancer cells. J. Inorg. Biochem. 2013, 127, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.M.; Sousa, A.J.; Hawley, S.P.; Vong, L.; Gareau, M.G.; Kumar, S.A.; Johnson-Henry, K.C.; Sherman, P.M. Matrix metalloproteinase 9 contributes to gut microbe homeostasis in a model of infectious colitis. BMC Microbiol. 2012, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, D.; Jena, R.; Choudhury, P.K.; Pattnaik, R.; Mohapatra, S.; Saini, M.R. Milk Derived Antimicrobial Bioactive Peptides: A Review. Int. J. Food Prop. 2015, 19, 837–846. [Google Scholar] [CrossRef]
- Expósito, I.L.; Recio, I. Antibacterial activity of peptides and folding variants from milk proteins. Int. Dairy J. 2006, 16, 1294–1305. [Google Scholar] [CrossRef]
- Florisa, R.; Recio, I.; Berkhout, B.; Visser, S. Antibacterial and antiviral effects of milk proteins and derivatives thereof. Curr. Pharm. Des. 2003, 9, 1257–1275. [Google Scholar] [CrossRef]
- Khan, M.U.; Pirzadeh, M.; Förster, C.Y.; Shityakov, S.; Shariati, M.A. Role of Milk-Derived Antibacterial Peptides in Modern Food Biotechnology: Their Synthesis, Applications and Future Perspectives. Biomolecules 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucht, H.D.; Raida, M.; Adermann, K.; Mägert, H.J.; Forssmann, W.G. Casocidin-I: A casein-alpha s2 derived peptide exhibits antibacterial activity. FEBS Lett. 1995, 372, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Baranyi, M.; Thomas, U.; Pellegrini, A. Antibacterial activity of casein-derived peptides isolated from rabbit (Oryctolagus cuniculus) milk. J. Dairy Res. 2003, 70, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Algaron, F.; Rizzello, C.G.; Fox, P.F.; Monnet, V.; Gobbetti, M. Angiotensin I-Converting-Enzyme-Inhibitory and Antibacterial Peptides from Lactobacillus helveticus PR4 Proteinase-Hydrolyzed Caseins of Milk from Six Species. Appl. Environ. Microbiol. 2003, 69, 5297–5305. [Google Scholar] [CrossRef] [Green Version]
- Chronopoulou, R.; Xylouri, E.; Fegeros, K.; Politis, I. The effect of two bovine beta-casein peptides on various functional properties of porcine macrophages and neutrophils: Differential roles of protein kinase A and exchange protein directly activated by cyclic AMP-1. Br. J. Nutr. 2006, 96, 553–561. [Google Scholar]
- Tagliabue, A.; Rappuoli, R. Changing priorities in vaccinology: Antibiotic resistance moving to the top. Front. Immunol. 2018, 9, 1068. [Google Scholar] [CrossRef]
- Bull, M.J.; Plummer, N.T. Part 1: The human gut microbiome in health and disease. Integr. Med. 2014, 13, 17–22. [Google Scholar]
- Alam, M.T.; Amos, G.C.; Murphy, A.R.; Murch, S.; Wellington, E.M.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 2020, 12, 1–8. [Google Scholar] [CrossRef]
- Sanches, S. Identificação de Péptidos Produzidos Pela Fermentação Láctica de Subprodutos das Indústrias de Lacticínios com Capacidade Inibidora das Gelatinases, MMP-9 e MMP-2. Master’s Thesis, Universidade do Porto, Porto, Portugal, 2015. Available online: https://hdl.handle.net/10216/79113 (accessed on 23 June 2021).
- Romano, M.; De Francesco, F.; Zarantonello, L.; Ruffolo, C.; Ferraro, G.A.; Zanus, G.; Giordano, A.; Bassi, N.; Cillo, U. From inflammation to cancer in inflammatory bowel disease: Molecular perspectives. Anticancer Res. 2016, 36, 1447–1460. [Google Scholar]
- Mohanty, D.P.; Mohapatra, S.; Misra, S.; Sahu, P.S. Milk derived bioactive peptides and their impact on human health–A review. Saudi J. Biol. Sci. 2016, 23, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Vandooren, J.; Van Den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef]
- Araújo, R.F., Jr.; Lira, G.A.; Vilaça, J.A.; Guedes, H.G.; Leitão, M.C.A.; Lucena, H.F.; Ramos, C.C.O. Prognostic and diagnostic implications of MMP-2, MMP-9, and VEGF-α expressions in colorectal cancer. Pathol. Res. Pract. 2015, 211, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, A.; Danesi, F.; Dardevet, D.; Dupont, D.; Fernandez, A.S.; Gille, D.; Dos Santos, C.N.; Pinto, P.; Re, R.; Rémond, D.; et al. Dairy products and inflammation: A review of the clinical evidence. Crit. Rev. Food Sci. Nutr. 2017, 57, 2497–2525. [Google Scholar] [CrossRef] [PubMed]
- DeMan, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of lactobacilli. J. Appl. Bacterial. 1960, 1, 23–130. [Google Scholar] [CrossRef]
- Lowry, O.H.; Brock, M.J.; Lopez, J.A. Protein measurement with Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Bouhdid, S.; Abrini, J.; Amensour, M.; Zhiri, A.; Espuny, M.J.; Manresa, A. Functional and ultrastructural changes in Pseudomonas aeruginosa and Staphylococcus aureus cells induced by Cinnamomum verum essential oil. J. Appl. Microbiol. 2010, 109, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.; Mota, J.; Monteiro, S.; Ferreira, R. Legume seeds and colorectal cancer revisited: Protease inhibitors reduce MMP-9 activity and colon cancer cell migration. Food Chem. 2016, 197, 30–38. [Google Scholar] [CrossRef]
- Toth, M.; Sohail, A.; Fridman, R. Assessment of Gelatinases (MMP-2 and MMP-9) by Gelatin Zymography. In Metastasis Research Protocols. Methods in Molecular Biology (Methods and Protocols); Dwek, M., Brooks, S., Schumacher, U., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 878, pp. 121–135. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC (μg/mL) | ||
|---|---|---|
| Days of Fermentation | L. monocytogenes | E. coli 0157 |
| 1 | ND | 25.0 |
| 2 | ND | 25.0 |
| 3 | ND | 25.0 |
| 4 | 25.0 | 25.0 |
| 5 | 25.0 | 25.0 |
| 6 | 0.20 | 0.40 |
| MIC (μg/mL) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Peak 1 | Peak 2 | Peak 3 | Peak 4 | Peak 5 | Peak 6 | Peak 7 | Peak 8 | Peak 9 | Peak 10 | Peak 11 | |
| L. monocytogenes | ND | 3 | ND | 50 | ND | ND | ND | ND | ND | ND | ND |
| E. coli | ND | 6 | 50 | ND | ND | ND | ND | ND | ND | ND | ND |
| N | Name | Peptides (95%) |
|---|---|---|
| 1 | Beta-casein OS=Bos taurus GN=CSN2 PE=1 SV=2 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, M.I.; Lima, A.; Mota, J.; Rebelo, P.; Ferreira, R.B.; Pedroso, L.; Ferreira, M.A.; Sousa, I. Extended Cheese Whey Fermentation Produces a Novel Casein-Derived Antibacterial Polypeptide That Also Inhibits Gelatinases MMP-2 and MMP-9. Int. J. Mol. Sci. 2021, 22, 11130. https://doi.org/10.3390/ijms222011130
Santos MI, Lima A, Mota J, Rebelo P, Ferreira RB, Pedroso L, Ferreira MA, Sousa I. Extended Cheese Whey Fermentation Produces a Novel Casein-Derived Antibacterial Polypeptide That Also Inhibits Gelatinases MMP-2 and MMP-9. International Journal of Molecular Sciences. 2021; 22(20):11130. https://doi.org/10.3390/ijms222011130
Chicago/Turabian StyleSantos, Maria Isabel, Ana Lima, Joana Mota, Patrícia Rebelo, Ricardo Boavida Ferreira, Laurentina Pedroso, Maria Adélia Ferreira, and Isabel Sousa. 2021. "Extended Cheese Whey Fermentation Produces a Novel Casein-Derived Antibacterial Polypeptide That Also Inhibits Gelatinases MMP-2 and MMP-9" International Journal of Molecular Sciences 22, no. 20: 11130. https://doi.org/10.3390/ijms222011130