
Novel NFκB Inhibitor SC75741 Mitigates Chondrocyte Degradation and Prevents Activated Fibroblast Transformation by Modulating miR-21/GDF-5/SOX5 Signaling


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
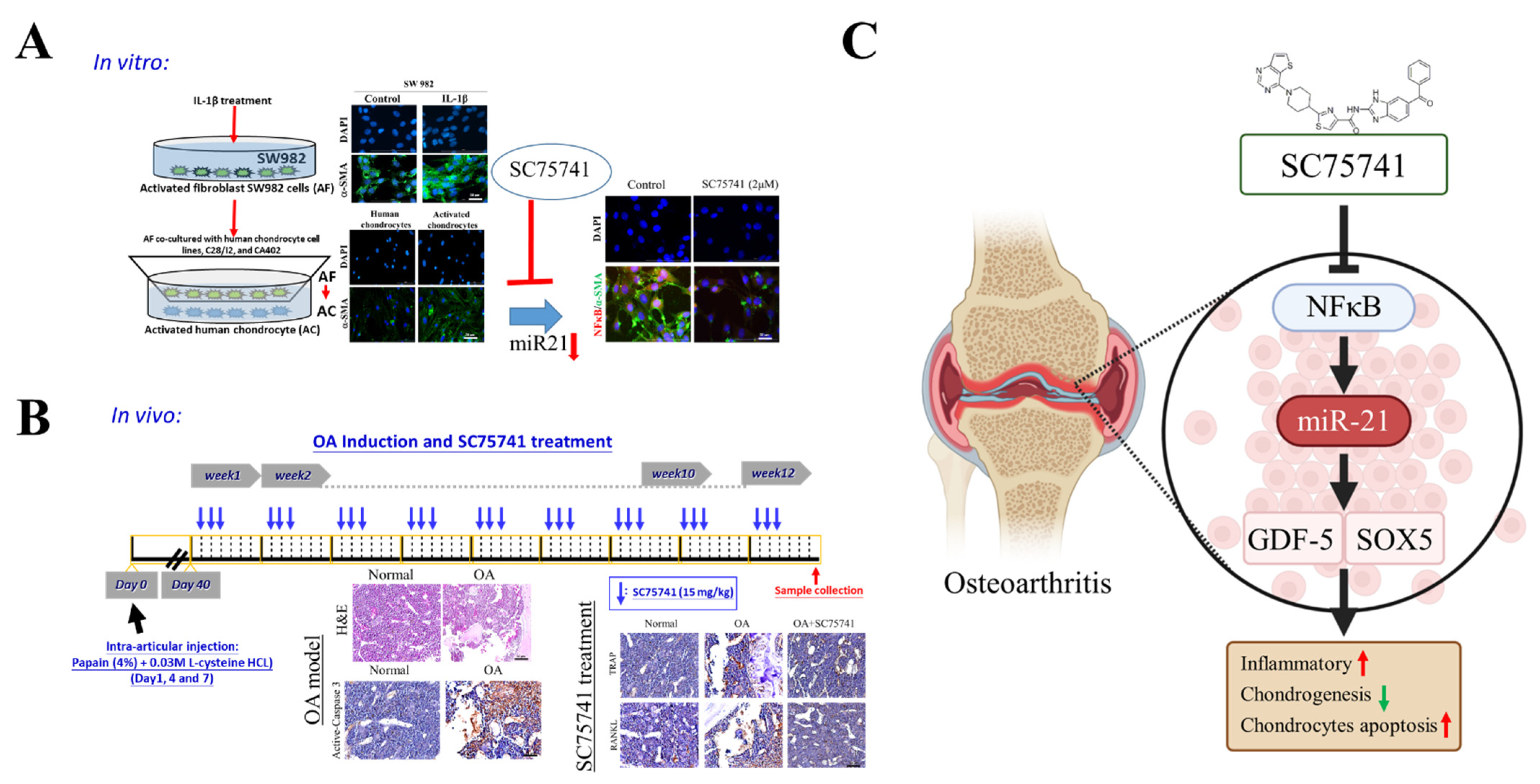
2.1. IL-1β-Activated Synovial Fibroblasts Exhibit Amplified Inflammatory Signaling in Chondrocytes
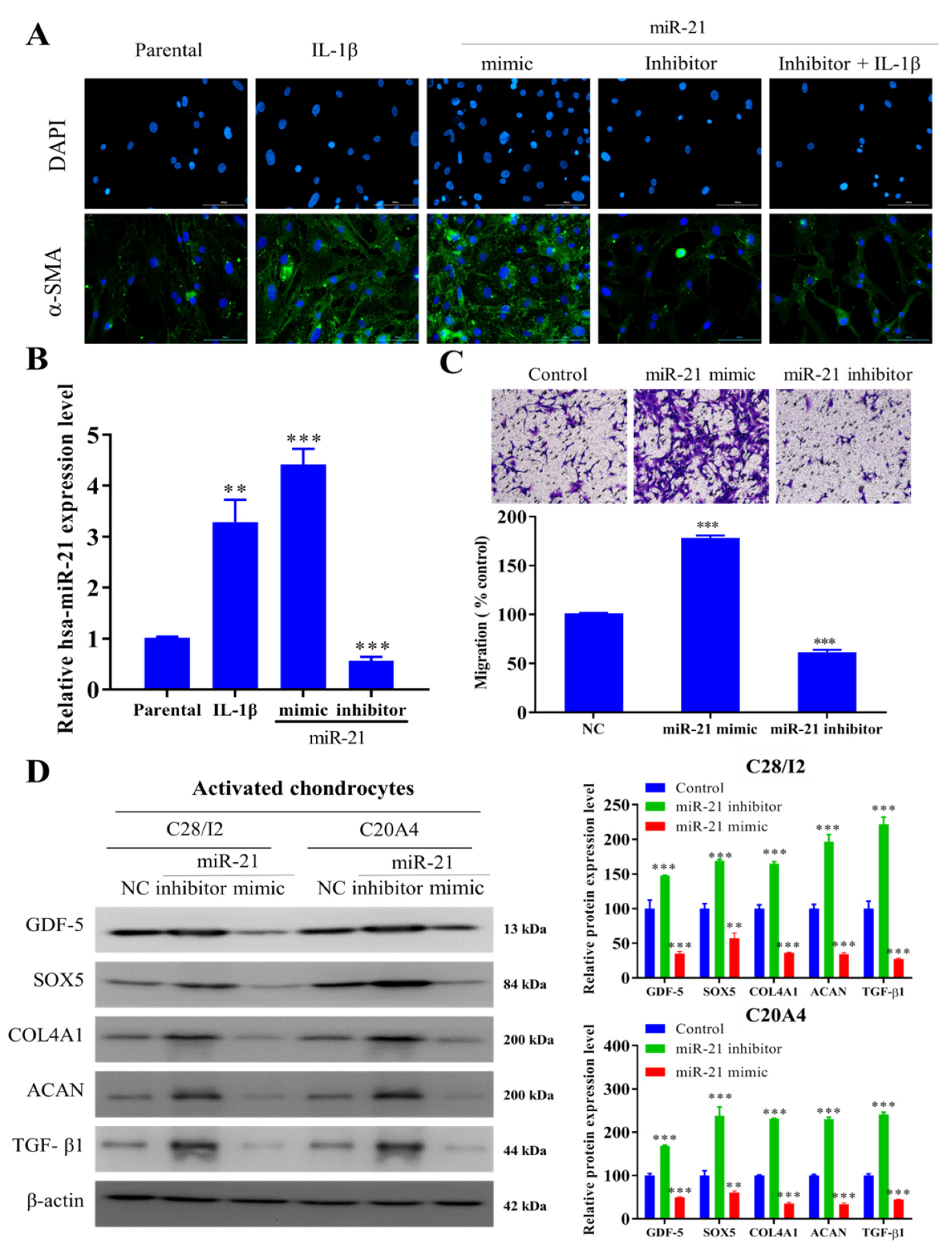
2.2. miR-21 Modulates the Activity of Activated Fibroblast-like Chondrocytes
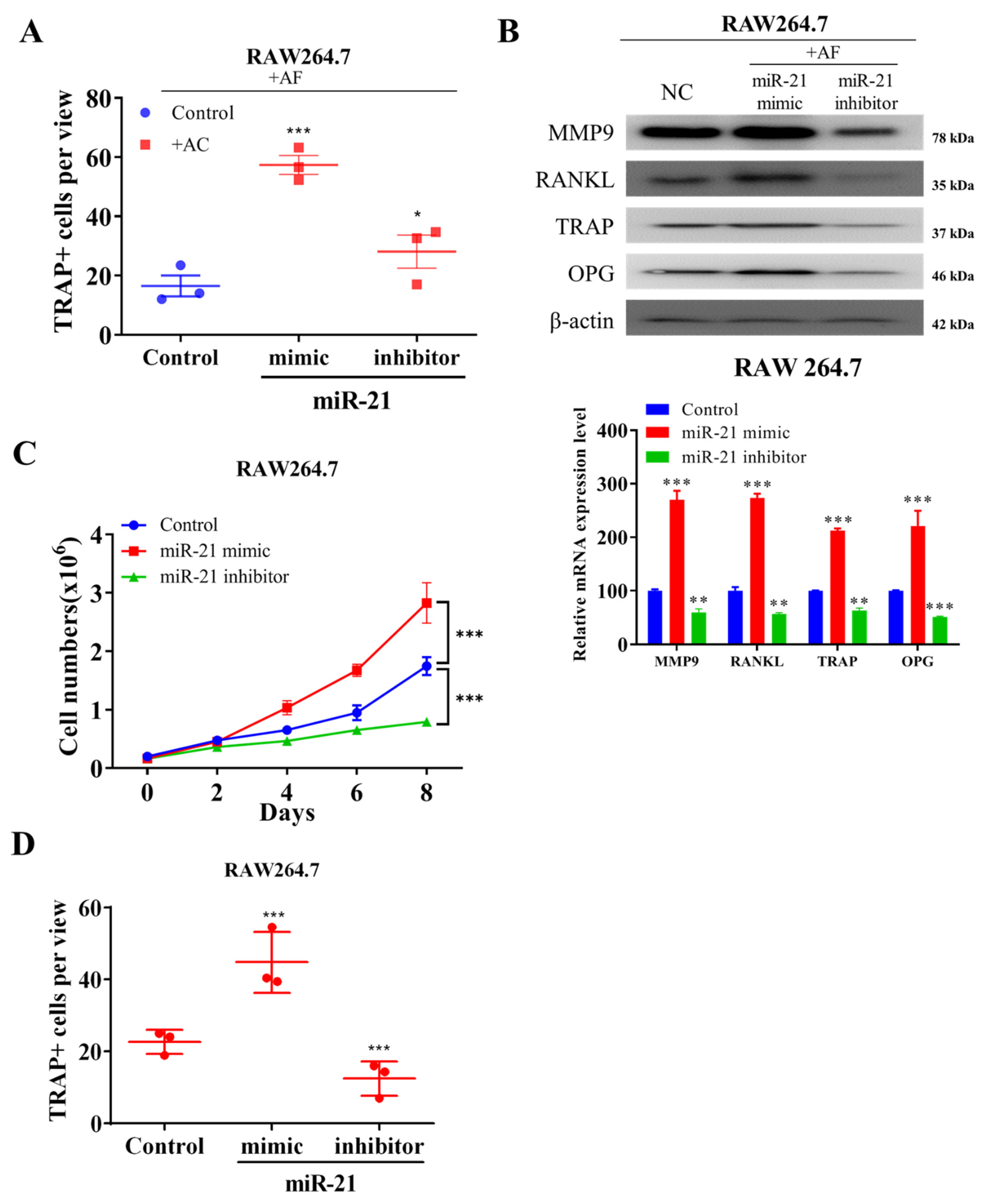
2.3. Ectopic Expression of miR-21 in Fibroblast-like Activated Chondrocytes Promote Osteoblast-Mediated Differentiation of Osteoclasts In Vitro
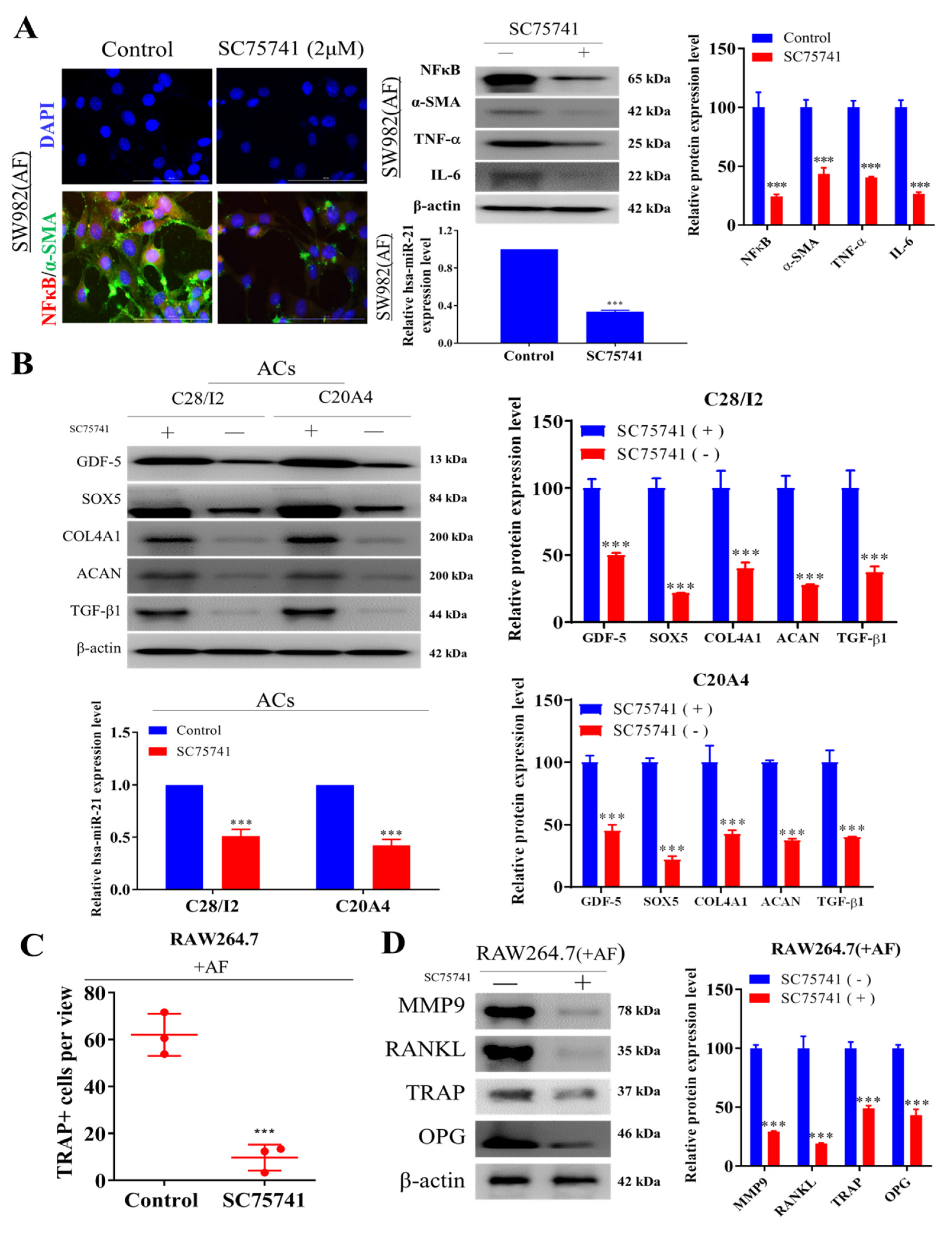
2.4. Inhibition of NF-κβ Dampens Cellular Transformation of Activated Fibroblasts, Chondrocytes, and Osteoclasts
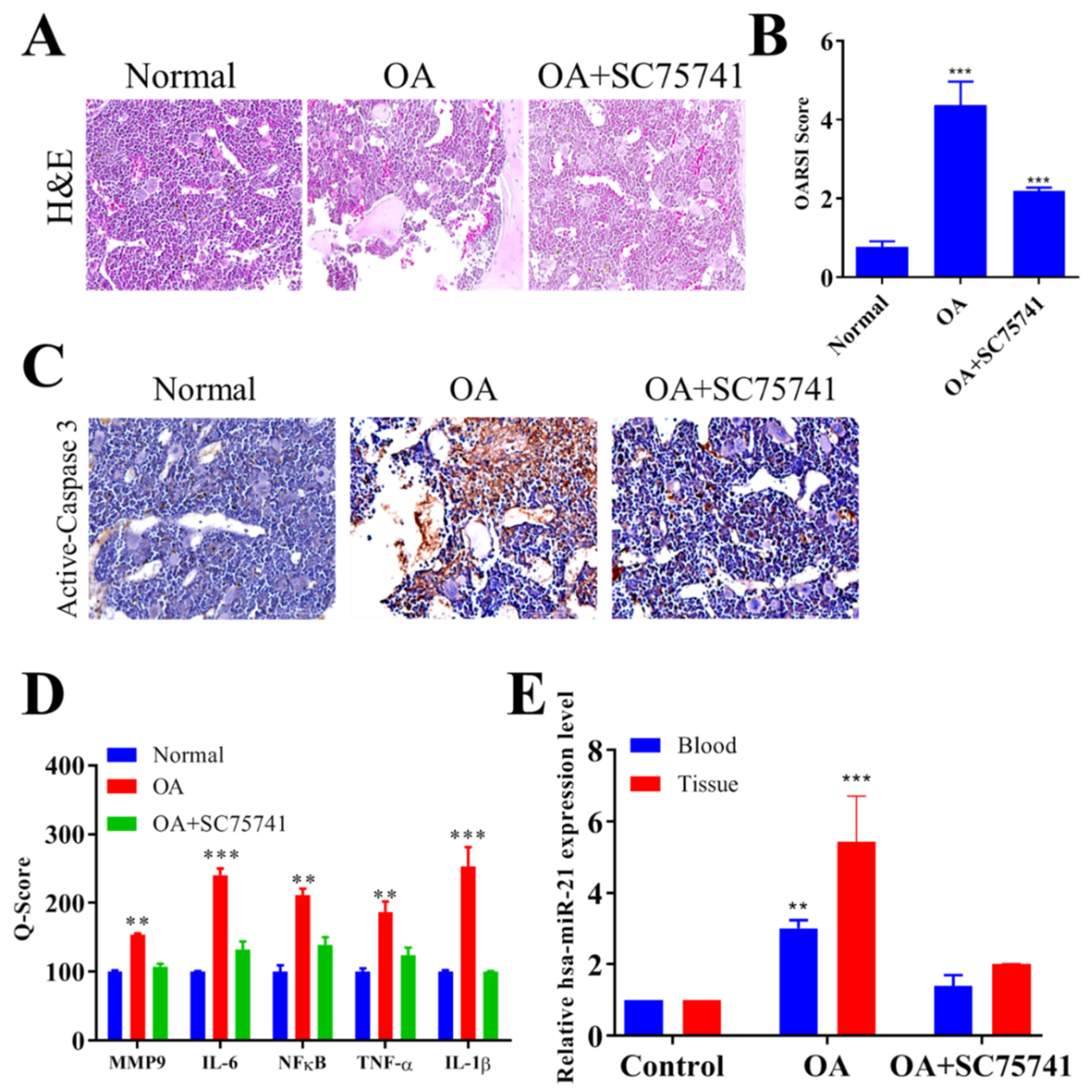
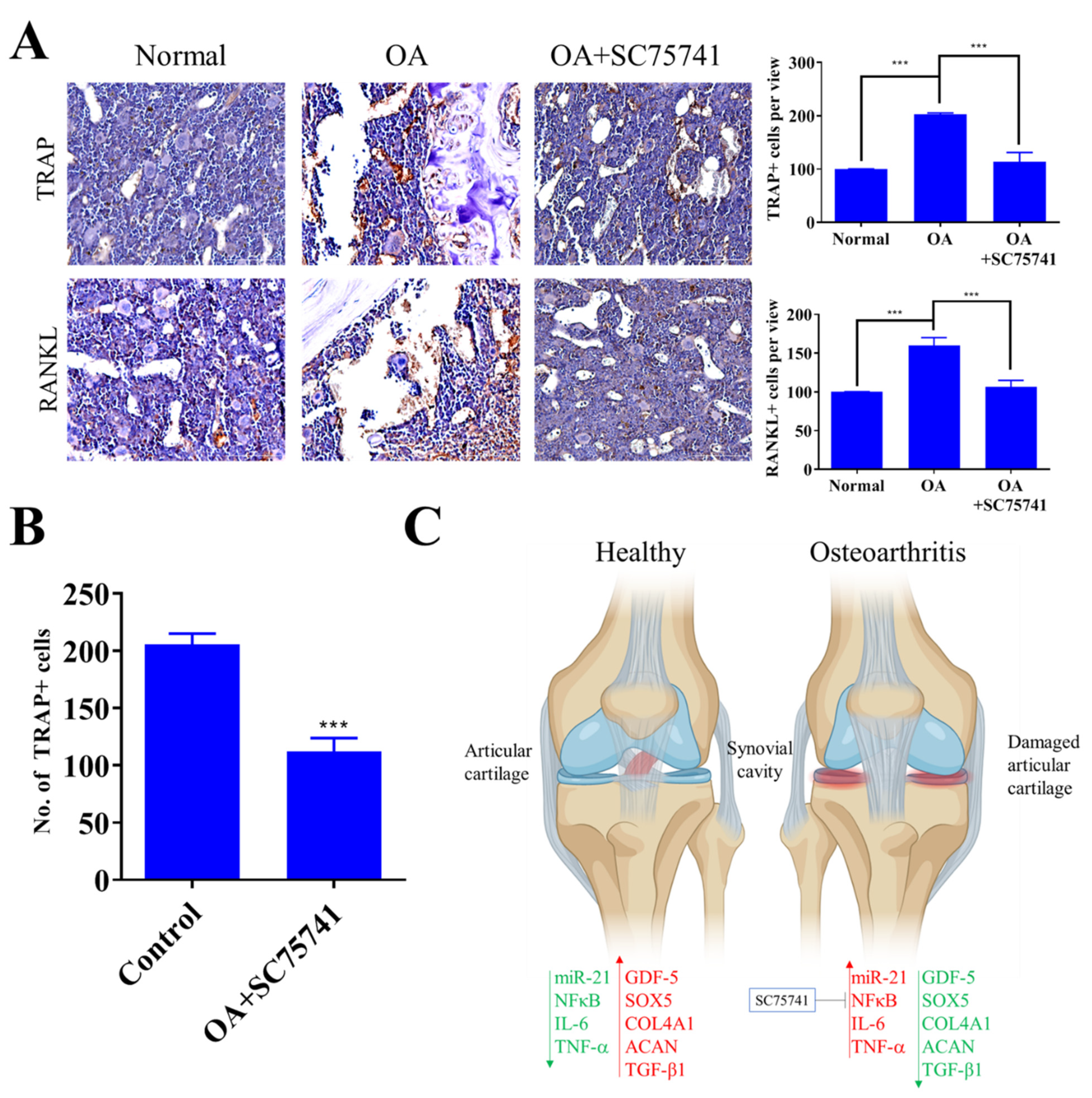
2.5. In Vivo Evaluation of the Therapeutic Potential of SC75741 in the OA Rat Model
2.6. In Vivo SC75741 Inhibited Chondroclast Differentiation and Activation
3. Discussion
4. Materials and Methods
4.1. Microarray Datasets Search
4.2. Cell Culture and In Vitro Osteoclastogenesis
4.3. Reagents
4.4. Cell Counting Kit-8 Cell Viability Assay
4.5. Total RNA Extraction and miR-21 Quantification by qRT-PCR and Transfections
4.6. Transwell Co-Culture and Migration Assays
4.7. Western Blot Assay
4.8. Immunofluorescence
4.9. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.10. In Vivo OA Rat Model
4.11. Histopathology, Immunohistochemical Staining and Scoring
4.12. Osteoarthritis Research Society International (OARSI) Score
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vina, E.R.; Kwoh, C.K. Epidemiology of osteoarthritis: Literature update. Curr. Opin. Rheumatol. 2018, 30, 160. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glyn-Jones, S.; Palmer, A.; Agricola, R.; Price, A.; Vincent, T.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Goldring, S.R.; Goldring, M.B. Clinical aspects, pathology and pathophysiology of osteoarthritis. J. Musculoskelet. Neuronal. Interact. 2006, 6, 376. [Google Scholar]
- Abusarah, J.; Benabdoune, H.; Shi, Q.; Lussier, B.; Martel-Pelletier, J.; Malo, M.; Fernandes, J.C.; de Souza, F.P.; Fahmi, H.; Benderdour, M. Elucidating the role of protandim and 6-gingerol in protection against osteoarthritis. J. Cell Biochem. 2017, 118, 1003–1013. [Google Scholar] [CrossRef]
- Scharstuhl, A.; Vitters, E.L.; van der Kraan, P.M.; van den Berg, W.B. Reduction of osteophyte formation and synovial thickening by adenoviral overexpression of transforming growth factor β/bone morphogenetic protein inhibitors during experimental osteoarthritis. Arthritis Rheum. 2003, 48, 3442–3451. [Google Scholar] [CrossRef]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.L.; Fang, C.; Liu, C.; Leslie, M.P.; Chang, E.; Cesare, P.E.D. Expression of bone morphogenetic proteins, receptors, and tissue inhibitors in human fetal, adult, and osteoarthritic articular cartilage. J. Orthop. Res. 2004, 22, 1188–1192. [Google Scholar] [CrossRef]
- Murphy, M.K.; Huey, D.J.; Hu, J.C.; Athanasiou, K.A. Tgf-β1, gdf-5, and bmp-2 stimulation induces chondrogenesis in expanded human articular chondrocytes and marrow-derived stromal cells. Stem Cells 2015, 33, 762–773. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef]
- Sellam, J.; Berenbaum, F. The role of synovitis in pathophysiology and clinical symptoms of osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, Y.; Wu, G.; Lin, B.; Li, Z.; Liu, X. Differential mirnaomics of the synovial membrane in knee osteoarthritis induced by bilateral anterior cruciate ligament transection in rats corrigendum in /10.3892/mmr.2019.10758. Mol. Med. Rep. 2018, 18, 4051–4057. [Google Scholar]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33. [Google Scholar] [CrossRef]
- Scanzello, C.R.; Goldring, S.R. The role of synovitis in osteoarthritis pathogenesis. Bone 2012, 51, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.D. Suppl 1: The normal synovium. Open Rheumatol. J. 2011, 5, 100. [Google Scholar] [CrossRef]
- Carr, H.L.; Turner, J.D.; Major, T.; Scheel-Toellner, D.; Filer, A. New developments in transcriptomic analysis of synovial tissue. Front. Med. 2020, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houard, X.; Goldring, M.B.; Berenbaum, F. Homeostatic mechanisms in articular cartilage and role of inflammation in osteoarthritis. Curr. Rheumatol. Rep. 2013, 15, 375. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.X.; Lin, L.; Wang, H.J.; Wei, X.L.; Fu, X.; Zhang, J.Y.; Yu, C.L. Suppression of early experimental osteoarthritis by in vivo delivery of the adenoviral vector-mediated nf-kappabp65-specific sirna. Osteoarthr. Cartil. 2008, 16, 174–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Nakasa, T.; Hikata, T.; Asahara, H. Molecular network of cartilage homeostasis and osteoarthritis. Med. Res. Rev. 2008, 28, 464–481. [Google Scholar] [CrossRef]
- Belluzzi, E.; Stocco, E.; Pozzuoli, A.; Granzotto, M.; Porzionato, A.; Vettor, R.; De Caro, R.; Ruggieri, P.; Ramonda, R.; Rossato, M.; et al. Contribution of infrapatellar fat pad and synovial membrane to knee osteoarthritis pain. Biomed. Res. Int. 2019, 2019, 6390182. [Google Scholar] [CrossRef]
- Jiang, L.-F.; Fang, J.-H.; Wu, L.-D. Role of infrapatellar fat pad in pathological process of knee osteoarthritis: Future applications in treatment. World J. Clin. Cases 2019, 7, 2134. [Google Scholar] [CrossRef]
- Marcu, K.B.; Otero, M.; Olivotto, E.; Borzi, R.M.; Goldring, M.B. Nf-kappab signaling: Multiple angles to target oa. Curr. Drug Targets 2010, 11, 599–613. [Google Scholar] [CrossRef]
- Nam, J.; Aguda, B.D.; Rath, B.; Agarwal, S. Biomechanical thresholds regulate inflammation through the nf-kappab pathway: Experiments and modeling. PLoS ONE 2009, 4, e5262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, R.; Liu, N.; Luo, S.; Huang, W.; Zha, Z.; Yang, J. Microrna-9 regulates the development of knee osteoarthritis through the nf-kappab1 pathway in chondrocytes. Medicine 2016, 95, e4315. [Google Scholar] [CrossRef]
- Roman-Blas, J.A.; Jimenez, S.A. Nf-kappab as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthr. Cartil. 2006, 14, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V. The functions of animal micrornas. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; He, A.; Wen, Y.; Xiao, X.; Hao, J.; Zhang, F.; Guo, X. Comparison of microrna expression profiles of kashin-beck disease, osteoarthritis and rheumatoid arthritis. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-L.; Wu, Y.-B.; Liu, J.-T.; Li, C.-Y. Upregulation of mir-98 inhibits apoptosis in cartilage cells in osteoarthritis. Genet. Test. Mol. Biomark. 2016, 20, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Cuadra, V.M.B.; González-Huerta, N.C.; Romero-Cordoba, S.; Hidalgo-Miranda, A.; Miranda-Duarte, A. Altered expression of circulating microrna in plasma of patients with primary osteoarthritis and in silico analysis of their pathways. PLoS ONE 2014, 9, e97690. [Google Scholar]
- Leban, J.; Baierl, M.; Mies, J.; Trentinaglia, V.; Rath, S.; Kronthaler, K.; Wolf, K.; Gotschlich, A.; Seifert, M.H. A novel class of potent nf-kappab signaling inhibitors. Bioorganic Med. Chem. Lett. 2007, 17, 5858–5862. [Google Scholar] [CrossRef] [PubMed]
- Haasbach, E.; Reiling, S.J.; Ehrhardt, C.; Droebner, K.; Rückle, A.; Hrincius, E.R.; Leban, J.; Strobl, S.; Vitt, D.; Ludwig, S.; et al. The nf-kappab inhibitor sc75741 protects mice against highly pathogenic avian influenza a virus. Antivir. Res. 2013, 99, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Hamberg, M.; Backes, C.; Fehlmann, T.; Hart, M.; Meder, B.; Meese, E.; Keller, A. Mirtargetlink–mirnas, genes and interaction networks. Int. J. Mol. Sci. 2016, 17, 564. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Zhang, Z.; Guo, W.; Ma, H.; Xu, B.; Mu, W.; Amat, A.; Cao, L. Isoliquiritigenin blunts osteoarthritis by inhibition of bone resorption and angiogenesis in subchondral bone. Sci. Rep. 2018, 8, 1721. [Google Scholar] [CrossRef] [PubMed]
- Lampiasi, N.; Russo, R.; Kireev, I.; Strelkova, O.; Zhironkina, O.; Zito, F. Osteoclasts differentiation from murine raw 264.7 cells stimulated by rankl: Timing and behavior. Biology 2021, 10, 117. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. Nf-κb signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. Nf-κb signaling pathways in osteoarthritic cartilage destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Markopoulos, G.S.; Roupakia, E.; Tokamani, M.; Alabasi, G.; Sandaltzopoulos, R.; Marcu, K.B.; Kolettas, E. Roles of nf-κb signaling in the regulation of mirnas impacting on inflammation in cancer. Biomedicines 2018, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.N.; Li, Y. Mir-21 and let-7 in the ras and nf-κb pathways. Microrna 2012, 1, 65–69. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Lee, S.H.; Na, H.-S.; Jung, K.; Choi, J.; Cho, K.H.; Lee, C.-Y.; Kim, S.J.; Park, S.-H.; Shin, D.-Y.; et al. Kartogenin inhibits pain behavior, chondrocyte inflammation, and attenuates osteoarthritis progression in mice through induction of il-10. Sci. Rep. 2018, 8, 13832. [Google Scholar] [CrossRef]
- Loeser, R.F. Aging and osteoarthritis: The role of chondrocyte senescence and aging changes in the cartilage matrix. Osteoarthr. Cartil. 2009, 17, 971–979. [Google Scholar] [CrossRef] [Green Version]
- Greene, M.A.; Loeser, R.F. Aging-related inflammation in osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1966–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, K.; Kubo, M.; Imai, S.; Toyoda, F.; Maeda, T.; Okumura, N.; Matsuura, H.; Matsusue, Y. The cox-2 selective blocker etodolac inhibits tnfα-induced apoptosis in isolated rabbit articular chondrocytes. Int. J. Mol. Sci. 2013, 14, 19705–19715. [Google Scholar] [CrossRef] [PubMed]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis. Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Confalone, E.; D’Alessio, G.; Furia, A. Il-6 induction by tnfα and il-1β in an osteoblast-like cell line. Int. J. Biomed. Sci. 2010, 6, 135–140. [Google Scholar] [PubMed]
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. Biomed. Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [Green Version]
- Schuerwegh, A.J.; Dombrecht, E.J.; Stevens, W.J.; Van Offel, J.F.; Bridts, C.H.; De Clerck, L.S. Influence of pro-inflammatory (IL-1 alpha, IL-6, TNF-alpha, IFN-gamma) and anti-inflammatory (il-4) cytokines on chondrocyte function. Osteoarthr. Cartil. 2003, 11, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Rigoglou, S.; Papavassiliou, A.G. The nf-kappab signalling pathway in osteoarthritis. Int. J. Biochem. Cell Biol. 2013, 45, 2580–2584. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Thielen, N.G.M.; van der Kraan, P.M.; van Caam, A.P.M. Tgfβ/bmp signaling pathway in cartilage homeostasis. Cells 2019, 8, 969. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Zhang, Z.; Liao, W.; Xu, D.; Fu, M.; Kang, Y. Screening of gene signatures for rheumatoid arthritis and osteoarthritis based on bioinformatics analysis. Mol. Med. Rep. 2016, 14, 1587–1593. [Google Scholar] [CrossRef] [Green Version]
- Rodan, G.A. Bone homeostasis. Proc. Natl. Acad. Sci. USA 1998, 95, 13361–13362. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.-S.; Hu, Y.; Xie, P.-L.; Cao, J.; Wang, Z.-X.; Liu, J.-H.; Yin, H.; Huang, J.; Tan, Y.-J.; Luo, J.; et al. Omentin-1 prevents inflammation-induced osteoporosis by downregulating the pro-inflammatory cytokines. Bone Res. 2018, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrhardt, C.; Rückle, A.; Hrincius, E.R.; Haasbach, E.; Anhlan, D.; Ahmann, K.; Banning, C.; Reiling, S.J.; Kühn, J.; Strobl, S.; et al. The nf-κb inhibitor sc75741 efficiently blocks influenza virus propagation and confers a high barrier for development of viral resistance. Cell. Microbiol. 2013, 15, 1198–1211. [Google Scholar] [CrossRef] [PubMed]
- Waldstein, W.; Perino, G.; Gilbert, S.L.; Maher, S.A.; Windhager, R.; Boettner, F. Oarsi osteoarthritis cartilage histopathology assessment system: A biomechanical evaluation in the human knee. J. Orthop. Res. 2016, 34, 135–140. [Google Scholar] [CrossRef]
- Hulejová, H.; Baresová, V.; Klézl, Z.; Polanská, M.; Adam, M.; Senolt, L. Increased level of cytokines and matrix metalloproteinases in osteoarthritic subchondral bone. Cytokine 2007, 38, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Odgren, P.R.; Witwicka, H.; Reyes-Gutierrez, P. The cast of clasts: Catabolism and vascular invasion during bone growth, repair, and disease by osteoclasts, chondroclasts, and septoclasts. Connect. Tissue Res. 2016, 57, 161–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. Ncbi geo: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2012, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar]
- Hayman, A.R. Tartrate-resistant acid phosphatase (trap) and the osteoclast/immune cell dichotomy. Autoimmunity 2008, 41, 218–223. [Google Scholar] [CrossRef]
- Murat, N.; Karadam, B.; Ozkal, S.; Karatosun, V.; Gidener, S. Quantification of papain-induced rat osteoarthritis in relation to time with the mankin score. Acta Orthop. Traumatol. Turc. 2007, 41, 233–237. [Google Scholar] [PubMed]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The oarsi histopathology initiative—Recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, P.-W.; Yadav, V.K.; Pikatan, N.W.; Fong, I.-H.; Lin, I.-H.; Yeh, C.-T.; Lee, W.-H. Novel NFκB Inhibitor SC75741 Mitigates Chondrocyte Degradation and Prevents Activated Fibroblast Transformation by Modulating miR-21/GDF-5/SOX5 Signaling. Int. J. Mol. Sci. 2021, 22, 11082. https://doi.org/10.3390/ijms222011082
Weng P-W, Yadav VK, Pikatan NW, Fong I-H, Lin I-H, Yeh C-T, Lee W-H. Novel NFκB Inhibitor SC75741 Mitigates Chondrocyte Degradation and Prevents Activated Fibroblast Transformation by Modulating miR-21/GDF-5/SOX5 Signaling. International Journal of Molecular Sciences. 2021; 22(20):11082. https://doi.org/10.3390/ijms222011082
Chicago/Turabian StyleWeng, Pei-Wei, Vijesh Kumar Yadav, Narpati Wesa Pikatan, Iat-Hang Fong, I-Hsin Lin, Chi-Tai Yeh, and Wei-Hwa Lee. 2021. "Novel NFκB Inhibitor SC75741 Mitigates Chondrocyte Degradation and Prevents Activated Fibroblast Transformation by Modulating miR-21/GDF-5/SOX5 Signaling" International Journal of Molecular Sciences 22, no. 20: 11082. https://doi.org/10.3390/ijms222011082