Dietary Salt Accelerates Orthodontic Tooth Movement by Increased Osteoclast Activity

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
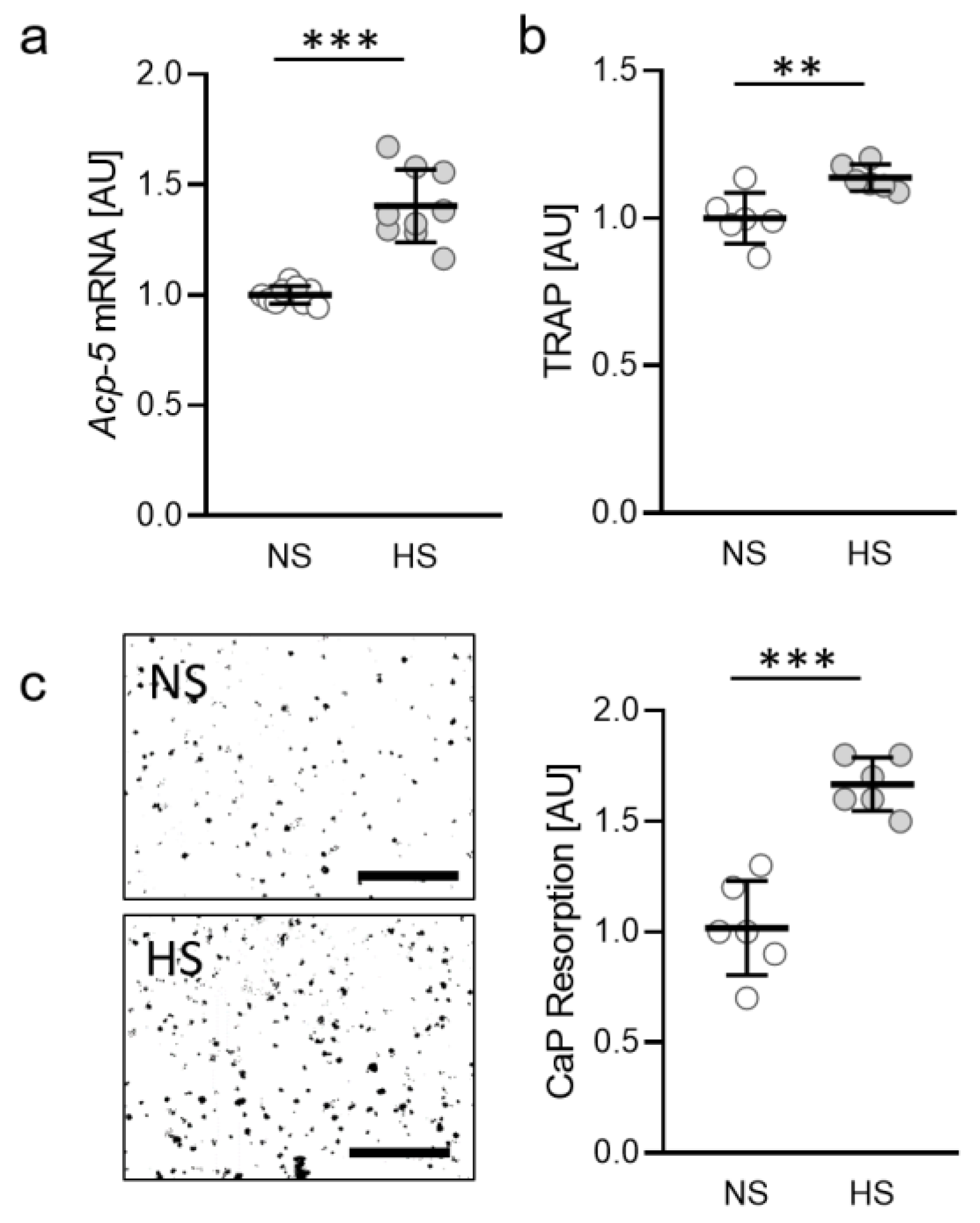
2.1. Impact of Salt on Osteoclast Activity
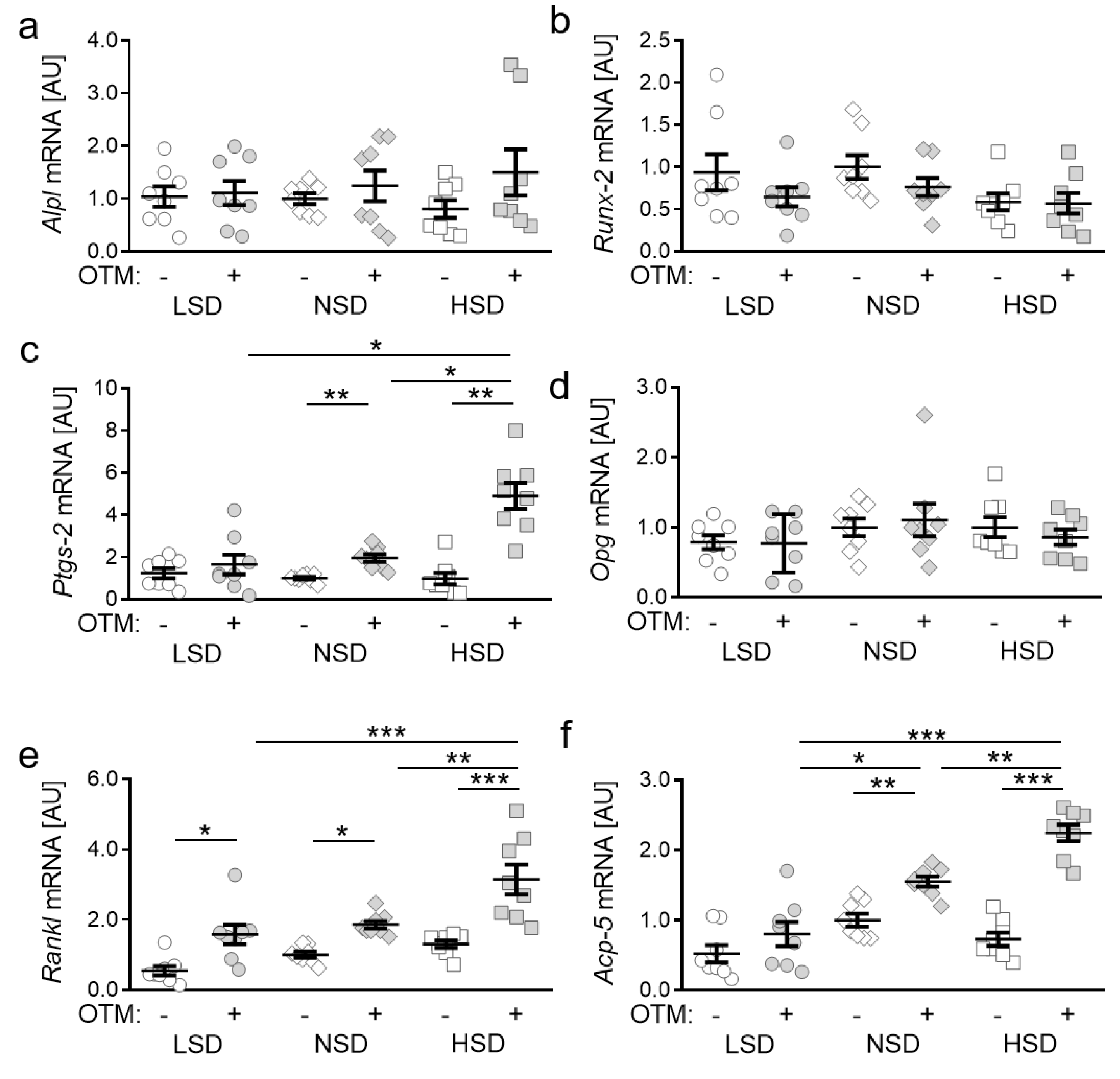
2.2. Impact of NaCl-Containing Diets on Expression of Genes Involved in Bone Remodelling
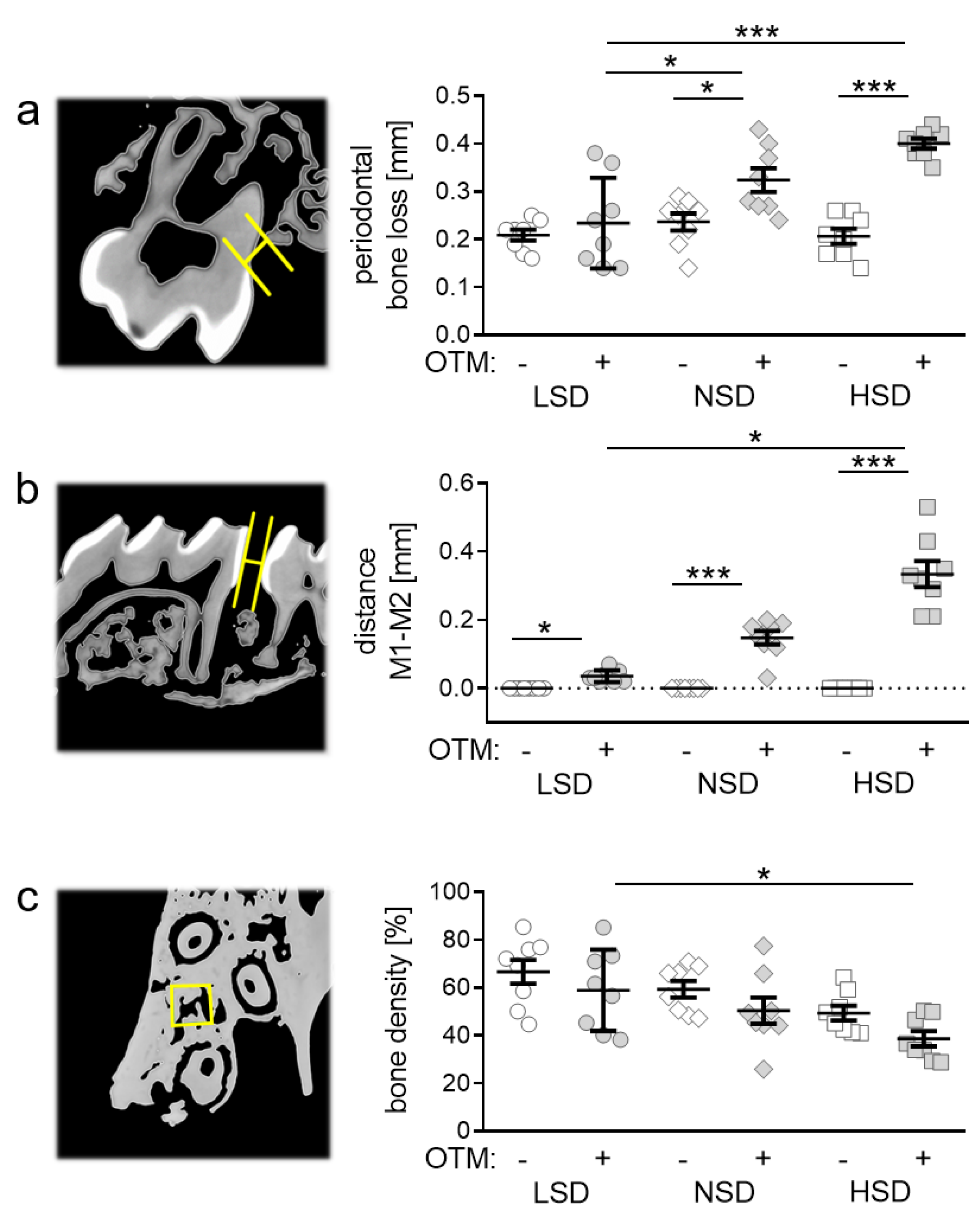
2.3. Effects of Different Salt Diets on Periodontal Bone Loss, Orthodontic Tooth Movement and Alveolar Bone Density
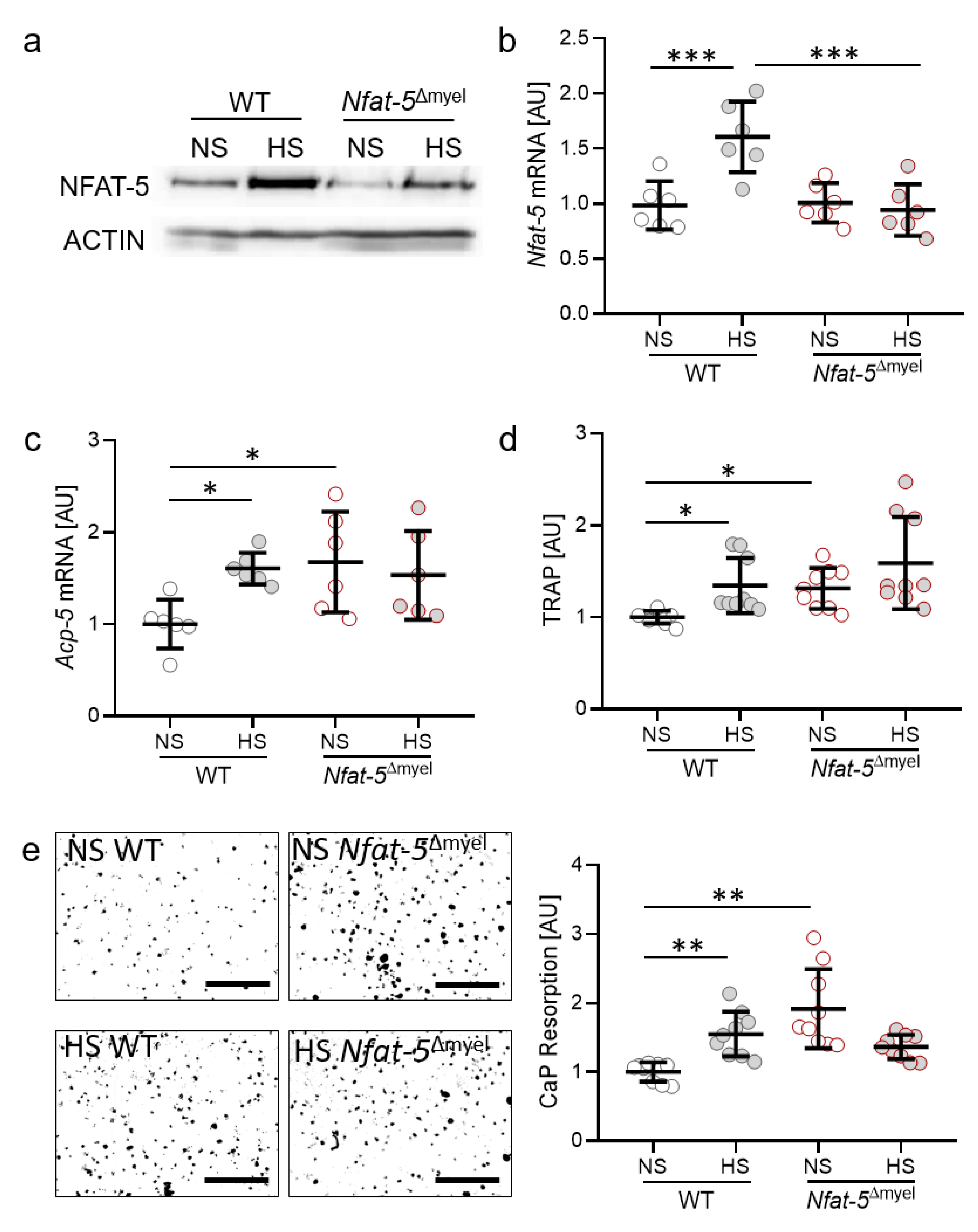
2.4. Impact of the Osmoprotective Transcription Factor NFAT-5 on Osteoclast Activity
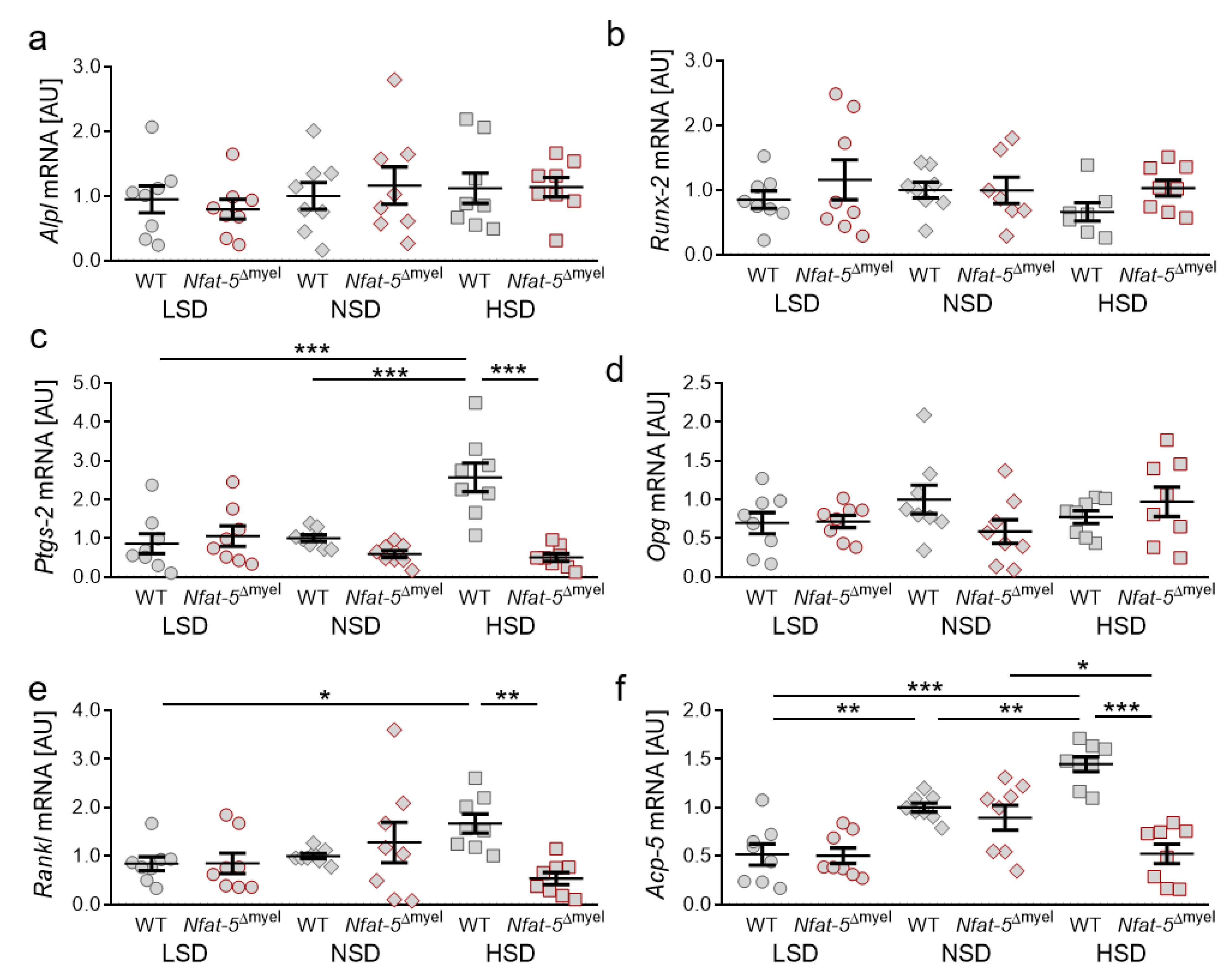
2.5. Impact of NFAT-5 Deletion in Myeloid Cells on the Expression of Genes Involved in Bone Remodelling
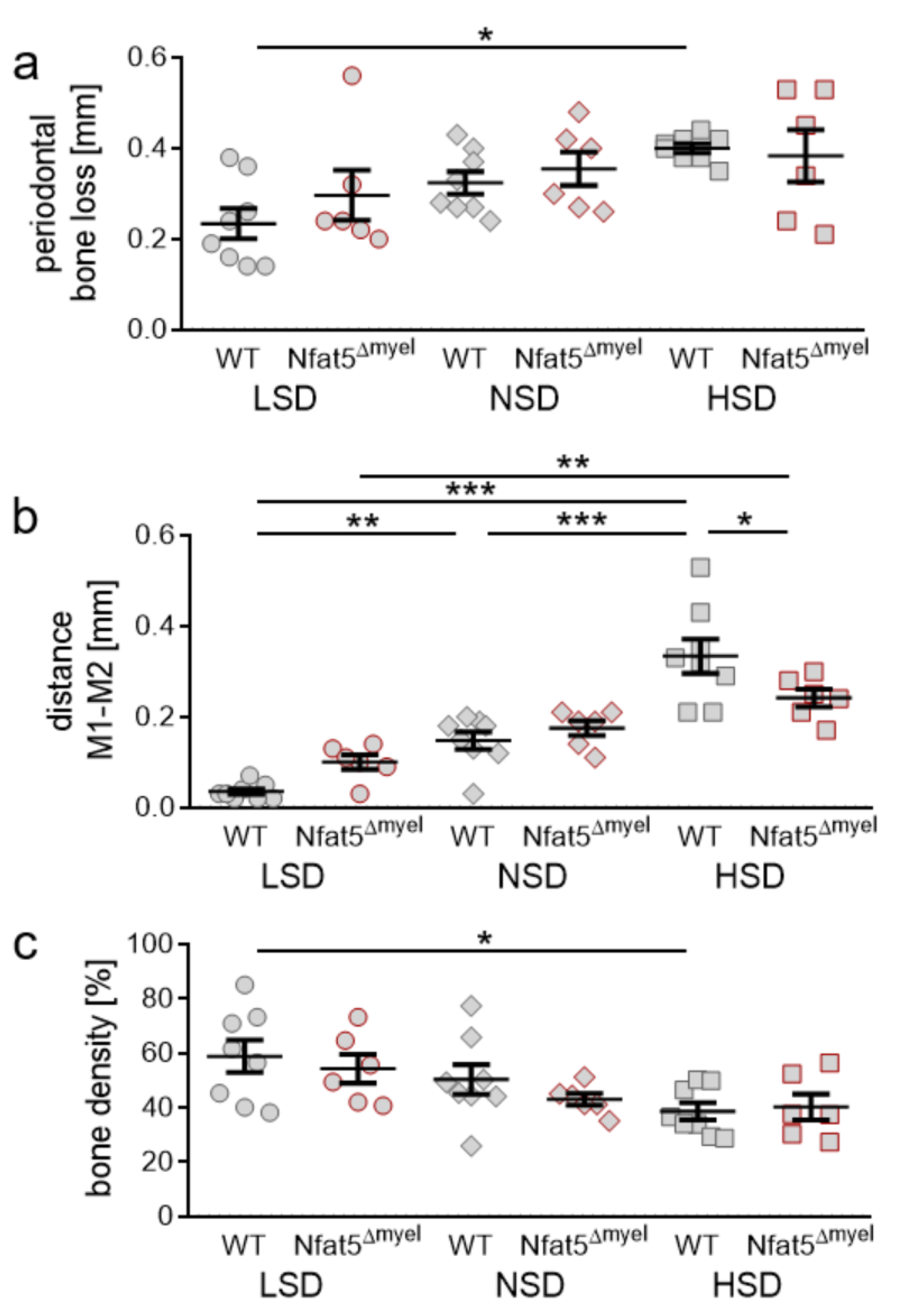
2.6. Impact of NFAT-5 Deletion in Myeloid Cells on Periodontal Bone Loss, Orthodontic Tooth Movement and Alveolar Bone Density
3. Discussion
4. Materials and Methods
4.1. Cell Culture Experiments
4.2. Animal Experiments
4.3. RNA Isolation from Cell Culture
4.4. RNA Isolation from Mouse Tissue and cDNA Synthesis
4.5. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.6. Tratrate-Resistant Acid Phosphatase (TRAP) Assay
4.7. Western Blot
4.8. Calcium Phosphate (CaP) Resorption Assay
4.9. Micro-Computed Tomography (µCT)-Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krishnan, V.; Davidovitch, Z. On a path to unfolding the biological mechanisms of orthodontic tooth movement. J. Dent. Res. 2009, 88, 597–608. [Google Scholar] [CrossRef]
- Feng, X.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Schinke, T.; Karsenty, G. The osteoblast: A sophisticated fibroblast under central surveillance. Science 2000, 289, 1501–1504. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Wong, B.R.; Rho, J.; Arron, J.; Robinson, E.; Orlinick, J.; Chao, M.; Kalachikov, S.; Cayani, E.; Bartlett, F.S.; Frankel, W.N.; et al. TRANCE is a novel ligand of the tumor necrosis factor receptor family that activates c-Jun N-terminal kinase in T cells. J. Biol. Chem. 1997, 272, 25190–25194. [Google Scholar] [CrossRef] [Green Version]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Kou, X.; Yang, R.; Liu, D.; Wang, X.; Luo, Q.; Song, Y.; Liu, F.; Yan, Y.; Gan, Y.; et al. M1-like Macrophage Polarization Promotes Orthodontic Tooth Movement. J. Dent. Res. 2015, 94, 1286–1294. [Google Scholar] [CrossRef]
- Wolf, M.; Lossdörfer, S.; Marciniak, J.; Römer, P.; Kirschneck, C.; Craveiro, R.; Deschner, J.; Jäger, A. CD8+ T cells mediate the regenerative PTH effect in hPDL cells via Wnt10b signaling. Innate Immun. 2016, 22, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef]
- Tsukasaki, M.; Takayanagi, H. Osteoimmunology: Evolving concepts in bone-immune interactions in health and disease. Nat. Rev. Immunol. 2019, 19, 626–642. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Li, Z.; Quan, H.; Xiao, L.; Zhao, J.; Wang, Y.; Liu, J.; Gou, Y.; An, S.; Huang, Y.; et al. Osteoimmunology in orthodontic tooth movement. Oral Dis. 2015, 21, 694–704. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guideline: Sodium Intake for Adults and Children; World Health Organization: Geneva, Switzerland, 2012; ISBN 9789241504836. [Google Scholar]
- Titze, J.; Rittweger, J.; Dietsch, P.; Krause, H.; Schwind, K.H.; Engelke, K.; Lang, R.; Kirsch, K.A.; Luft, F.C.; Hilgers, K.F. Hypertension, sodium retention, calcium excretion and osteopenia in Dahl rats. J. Hypertens. 2004, 22, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Wiig, H.; Schröder, A.; Neuhofer, W.; Jantsch, J.; Kopp, C.; Karlsen, T.V.; Boschmann, M.; Goss, J.; Bry, M.; Rakova, N.; et al. Immune cells control skin lymphatic electrolyte homeostasis and blood pressure. J. Clin. Investig. 2013, 123, 2803–2815. [Google Scholar] [CrossRef]
- Machnik, A.; Neuhofer, W.; Jantsch, J.; Dahlmann, A.; Tammela, T.; Machura, K.; Park, J.K.; Beck, F.X.; Müller, D.N.; Derer, W.; et al. Macrophages regulate salt-dependent volume and blood pressure by a vascular endothelial growth factor-C-dependent buffering mechanism. Nat. Med. 2009, 15, 545–552. [Google Scholar] [CrossRef]
- Titze, J.; Machnik, A. Sodium sensing in the interstitium and relationship to hypertension. Curr. Opin. Nephrol. Hypertens. 2010, 19, 385–392. [Google Scholar] [CrossRef]
- Schröder, A.; Nazet, U.; Neubert, P.; Jantsch, J.; Spanier, G.; Proff, P.; Kirschneck, C. Sodium-chloride-induced effects on the expression profile of human periodontal ligament fibroblasts with focus on simulated orthodontic tooth movement. Eur. J. Oral Sci. 2019, 127, 386–395. [Google Scholar] [CrossRef]
- Schröder, A.; Neubert, P.; Titze, J.; Bozec, A.; Neuhofer, W.; Proff, P.; Kirschneck, C.; Jantsch, J. Osteoprotective action of low-salt diet requires myeloid cell-derived NFAT5. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Jantsch, J.; Schatz, V.; Friedrich, D.; Schröder, A.; Kopp, C.; Siegert, I.; Maronna, A.; Wendelborn, D.; Linz, P.; Binger, K.J.; et al. Cutaneous Na+ storage strengthens the antimicrobial barrier function of the skin and boosts macrophage-driven host defense. Cell Metab. 2015, 21, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Schröder, A.; Leikam, A.; Käppler, P.; Neubert, P.; Jantsch, J.; Neuhofer, W.; Deschner, J.; Proff, P.; Kirschneck, C. Impact of salt and the osmoprotective transcription factor NFAT-5 on macrophages during mechanical strain. Immunol. Cell Biol. 2020. [Google Scholar] [CrossRef]
- Binger, K.J.; Gebhardt, M.; Heinig, M.; Rintisch, C.; Schroeder, A.; Neuhofer, W.; Hilgers, K.; Manzel, A.; Schwartz, C.; Kleinewietfeld, M.; et al. High salt reduces the activation of IL-4- and IL-13-stimulated macrophages. J. Clin. Investig. 2015, 125, 4223–4238. [Google Scholar] [CrossRef] [PubMed]
- Neuhofer, W. Role of NFAT5 in inflammatory disorders associated with osmotic stress. Curr. Genom. 2010, 11, 584–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.Y.; Lee-Kwon, W.; Kwon, H.M. The evolving role of TonEBP as an immunometabolic stress protein. Nat. Rev. Nephrol. 2020, 16, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Bauer, K.; Spanier, G.; Proff, P.; Wolf, M.; Kirschneck, C. Expression kinetics of human periodontal ligament fibroblasts in the early phases of orthodontic tooth movement. J. Orofac. Orthop. 2018, 79, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Kitada, K.; Daub, S.; Zhang, Y.; Klein, J.D.; Nakano, D.; Pedchenko, T.; Lantier, L.; LaRocque, L.M.; Marton, A.; Neubert, P.; et al. High salt intake reprioritizes osmolyte and energy metabolism for body fluid conservation. J. Clin. Investig. 2017, 127, 1944–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakova, N.; Kitada, K.; Lerchl, K.; Dahlmann, A.; Birukov, A.; Daub, S.; Kopp, C.; Pedchenko, T.; Zhang, Y.; Beck, L.; et al. Increased salt consumption induces body water conservation and decreases fluid intake. J. Clin. Investig. 2017, 127, 1932–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albano, G.; Dolder, S.; Siegrist, M.; Mercier-Zuber, A.; Auberson, M.; Stoudmann, C.; Hofstetter, W.; Bonny, O.; Fuster, D.G. Increased bone resorption by osteoclast-specific deletion of the sodium/calcium exchanger isoform 1 (NCX1). Pflugers Arch. 2017, 469, 225–233. [Google Scholar] [CrossRef]
- Gupta, A.; Guo, X.L.; Alvarez, U.M.; Hruska, K.A. Regulation of sodium-dependent phosphate transport in osteoclasts. J. Clin. Investig. 1997, 100, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, W.; Siegrist, M.; Simonin, A.; Bonny, O.; Fuster, D.G. Sodium/hydrogen exchanger NHA2 in osteoclasts: Subcellular localization and role in vitro and in vivo. Bone 2010, 47, 331–340. [Google Scholar] [CrossRef]
- Schröder, A.; Gubernator, J.; Nazet, U.; Spanier, G.; Jantsch, J.; Proff, P.; Kirschneck, C. Effects of sodium chloride on the gene expression profile of periodontal ligament fibroblasts during tensile strain. J. Orofac. Orthop. 2020, 81, 360–370. [Google Scholar] [CrossRef]
- Wu, L.; Luthringer, B.J.C.; Feyerabend, F.; Zhang, Z.; Machens, H.G.; Maeda, M.; Taipaleenmäki, H.; Hesse, E.; Willumeit-Römer, R.; Schilling, A.F. Increased levels of sodium chloride directly increase osteoclastic differentiation and resorption in mice and men. Osteoporos. Int. 2017, 28, 3215–3228. [Google Scholar] [CrossRef] [PubMed]
- WALDO, C.M.; ROTHBLATT, J.M. Histologic response to tooth movement in the laboratory rat; procedure and preliminary observations. J. Dent. Res. 1954, 33, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Maria, S.M.; Prukner, C.; Sheikh, Z.; Mueller, F.; Barralet, J.E.; Komarova, S.V. Reproducible quantification of osteoclastic activity: Characterization of a biomimetic calcium phosphate assay. J. Biomed. Mater. Res. Part B Appl. Biomater. 2014, 102, 903–912. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | Number | 5′-Forward Primer-3′ | 5′-Reverse Primer-3′ |
|---|---|---|---|---|
| Acp-5 | acid phosphatase-5, tartrate resistant | NM_007388.3 | ATACGGGGTCACTGCCTACC | TCGTTGATGTCGCACAGAGG |
| Alpl | alkaline phosphatase, tartrate resistant | NM_007431.3 | GGGGTACAAGGCTAGATGGC | AGTTCAGTGCGGTTCCAGAC |
| Ptgs-2 | prostaglandin endoperoxide synthase-2 | NM_011198.4 | TCCCTGAAGCCGTACACATC | TCCCCAAAGATAGCATCTGGAC |
| Eef1a1 | eukaryotic translation elongation factor 1 alpha 1 | NM_010106.2 | AAAACATGATTACAGGCACATCCC | GCCCGTTCTTGGAGATACCAG |
| Nfat-5 | nuclear factor of activated T cells-5 | NM_133957.3 | AAATGACCTGTAGTTCTCTGCTTC | GCTGTCGGTGACTGAGGTAG |
| Polr2a | polymerase (RNA) II (DNA directed) polypeptide A | NM_001291068.1 | CGGATGGTGTGAGCCTGATG | GTTCTCGCTCCAGAGCCTTC |
| Opg | osteoprotegerin | NM_008764.3 | CCTTGCCCTGACCACTCTTAT | CACACACTCGGTTGTGGGT |
| Rankl | receptor activator of NF-kB ligand | NM_011613.3 | AAACGCAGATTTGCAGGACTC | CCCCACAATGTGTTGCAGTTC |
| Runx-2 | runt related transcription factor-2 | NM_009820.5 | GACGTGCCCAGGCGTATTTC | CACCTGCCTGGCTCTTCTTAC |
| Sdha | succinate dehydrogenase complex, subunit A | NM_023281.1 | AACACTGGAGGAAGCACACC | AGTAGGAGCGGATAGCAGGAG |
| Ywhaz | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta | NM_011740.3 | AATGCTTCGCAACCAGAAAGC | TGGTATGCTTGCTGTGACTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, A.; Gubernator, J.; Leikam, A.; Nazet, U.; Cieplik, F.; Jantsch, J.; Neubert, P.; Titze, J.; Proff, P.; Kirschneck, C. Dietary Salt Accelerates Orthodontic Tooth Movement by Increased Osteoclast Activity. Int. J. Mol. Sci. 2021, 22, 596. https://doi.org/10.3390/ijms22020596
Schröder A, Gubernator J, Leikam A, Nazet U, Cieplik F, Jantsch J, Neubert P, Titze J, Proff P, Kirschneck C. Dietary Salt Accelerates Orthodontic Tooth Movement by Increased Osteoclast Activity. International Journal of Molecular Sciences. 2021; 22(2):596. https://doi.org/10.3390/ijms22020596
Chicago/Turabian StyleSchröder, Agnes, Joshua Gubernator, Alexandra Leikam, Ute Nazet, Fabian Cieplik, Jonathan Jantsch, Patrick Neubert, Jens Titze, Peter Proff, and Christian Kirschneck. 2021. "Dietary Salt Accelerates Orthodontic Tooth Movement by Increased Osteoclast Activity" International Journal of Molecular Sciences 22, no. 2: 596. https://doi.org/10.3390/ijms22020596