Inflammation in the Human Periodontium Induces Downregulation of the α1- and β1-Subunits of the sGC in Cementoclasts

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
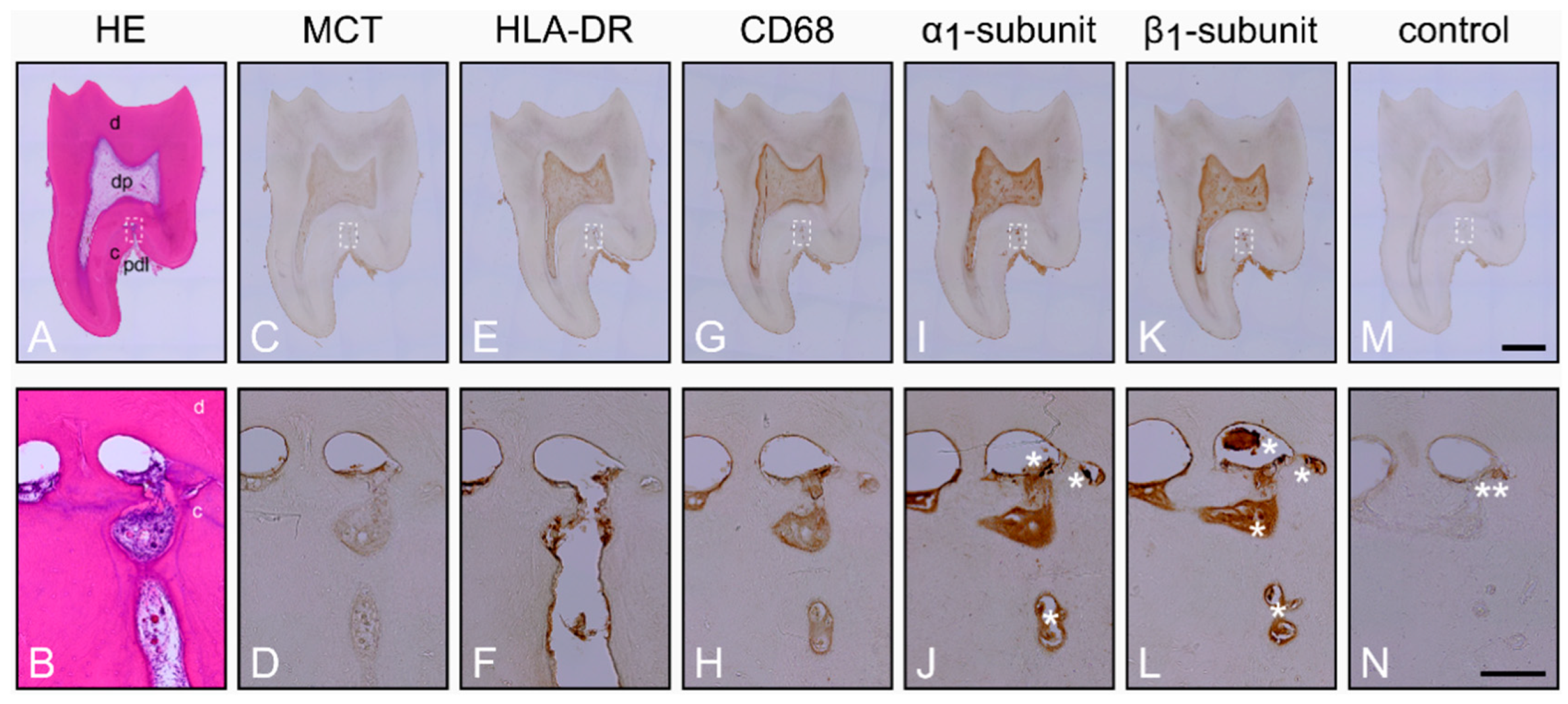
2.1. Characterization of the Healthy Periodontium and Presence of the α1- and β1-Subunits of sGC in Cementoclasts
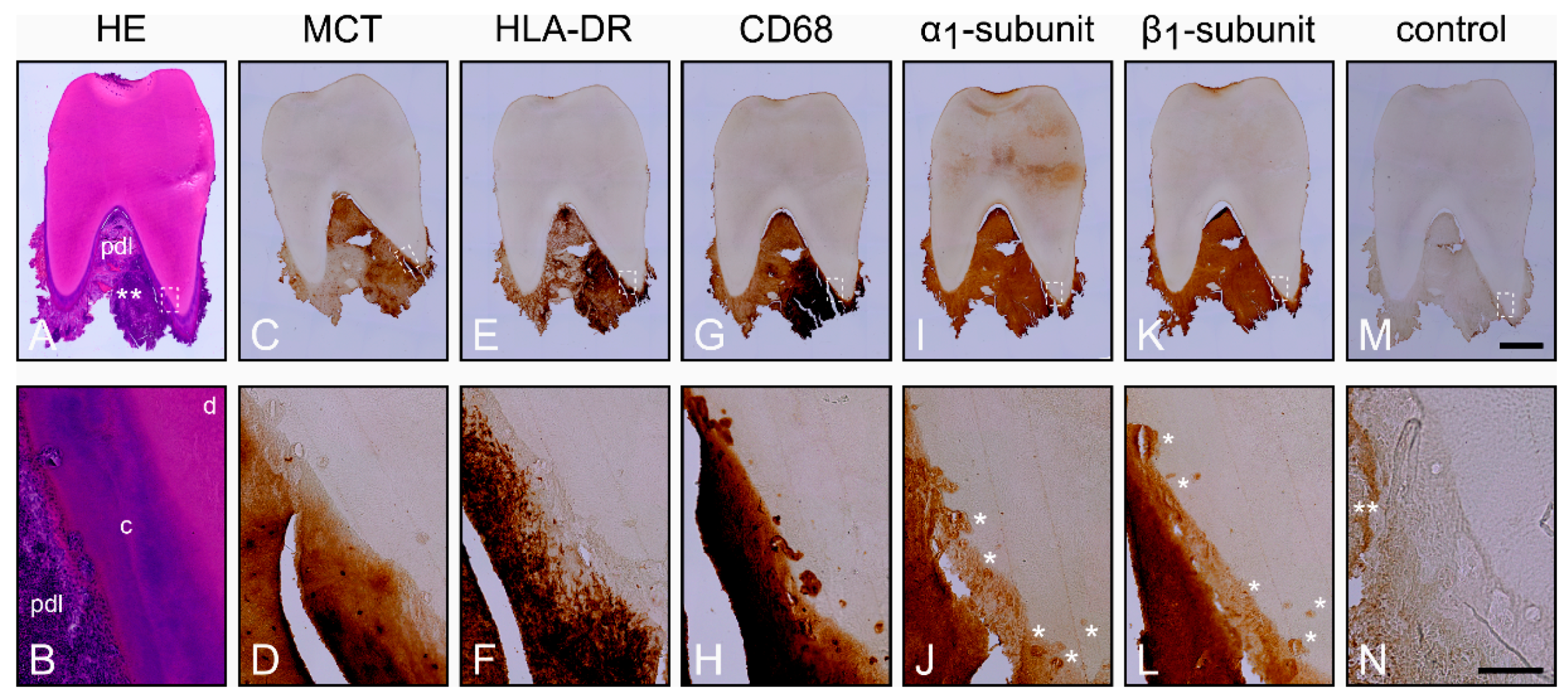
2.2. Characterization of the Inflamed Periodontium and Presence of the α1- and β1-Subunits of sGC in Cementoclasts
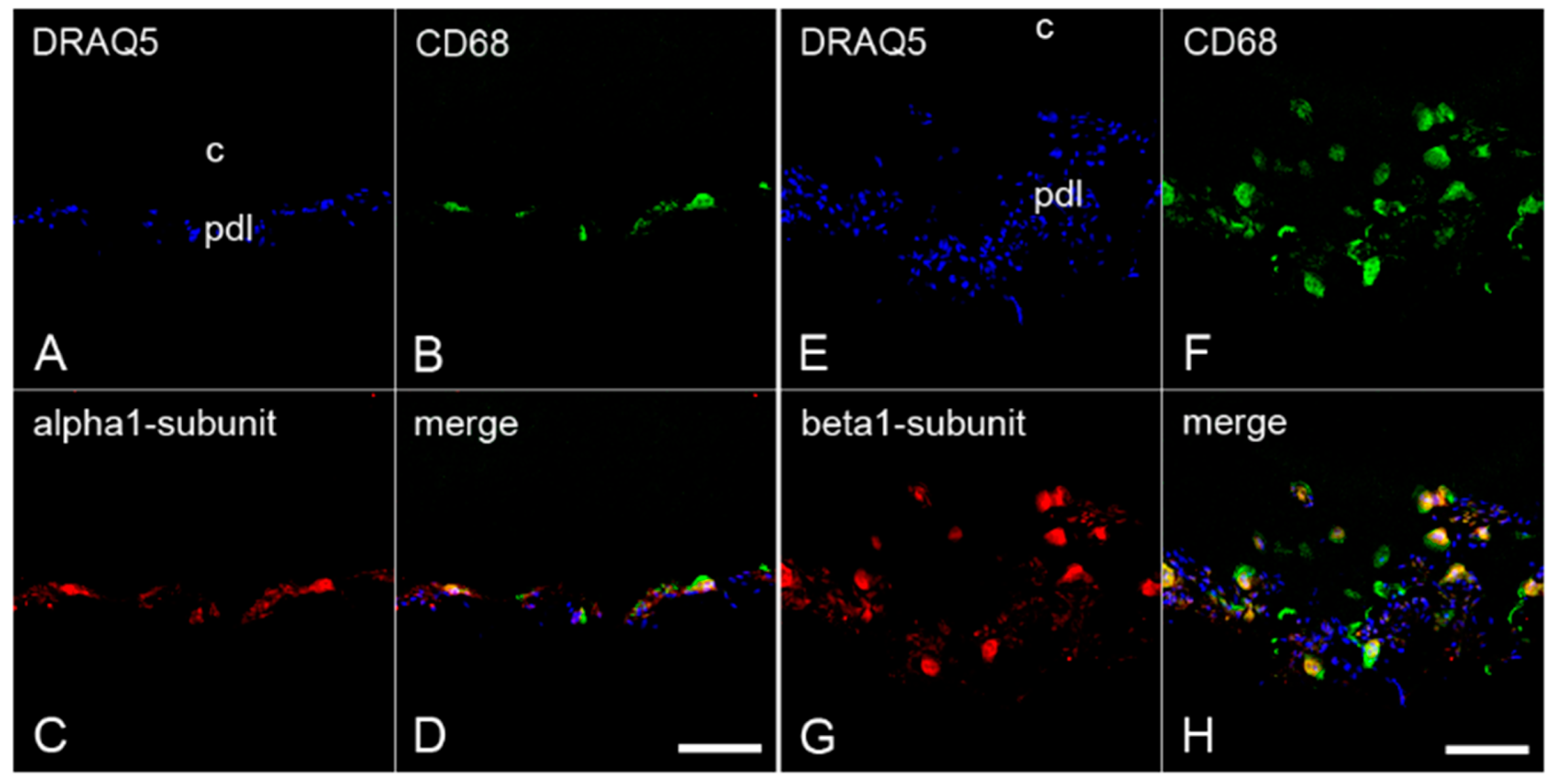
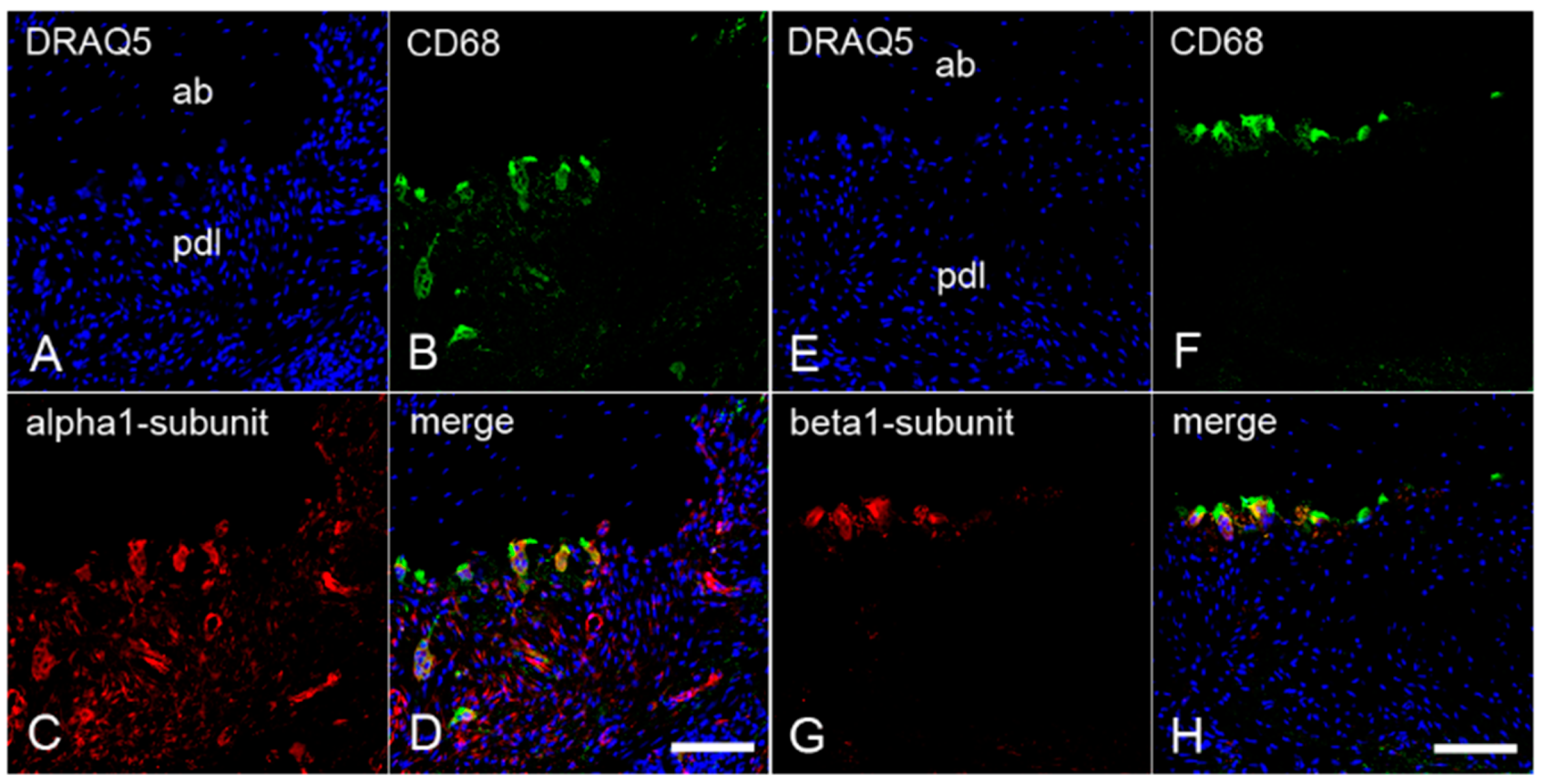
2.3. The Colocalization of α1- and β1-Subunits of sGC with CD68 in Cementoclasts of Inflamed Human Periodontium
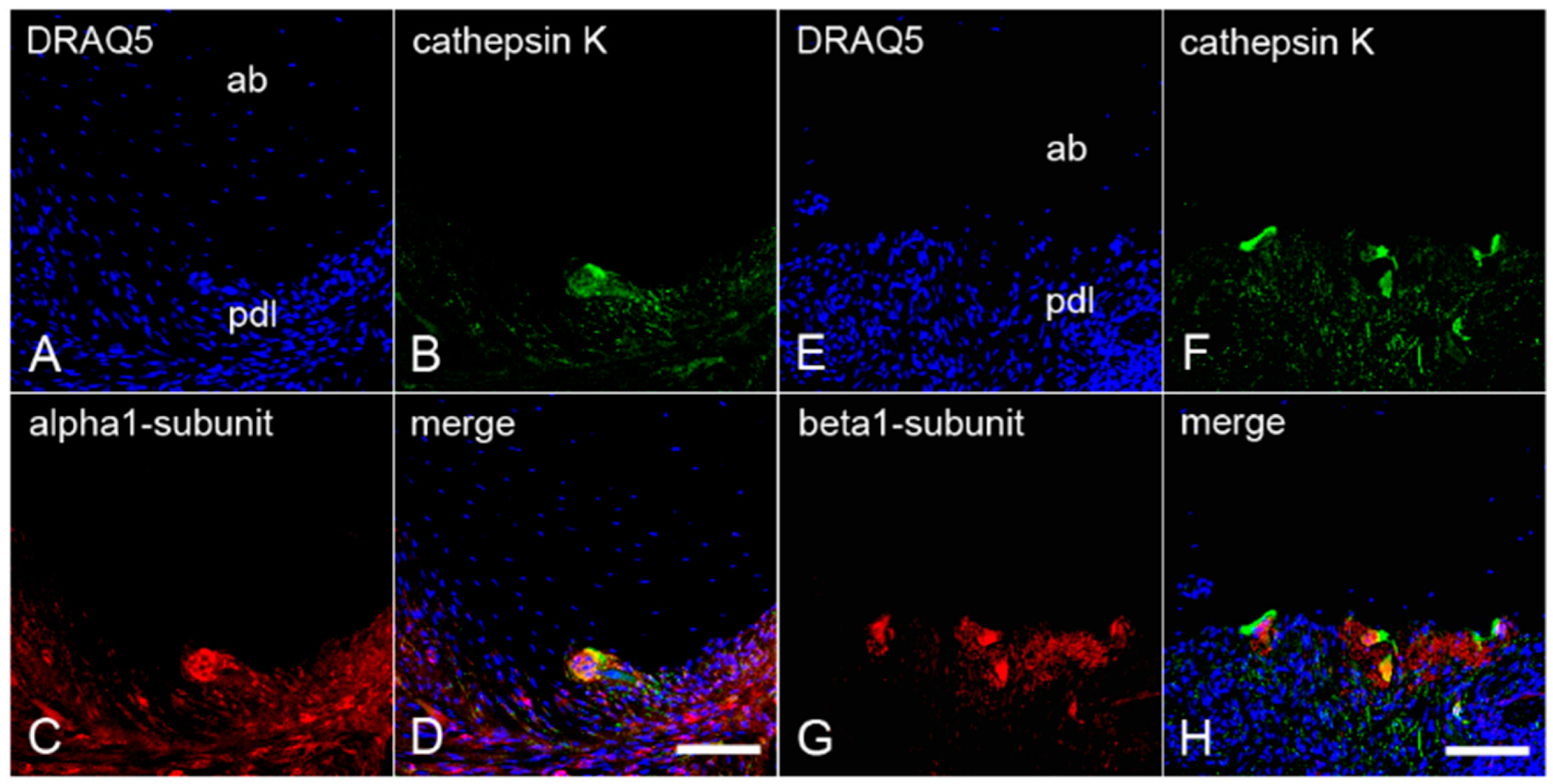
2.4. The Colocalization of α1- and β1-Subunits of sGC with Cathepsin K in Cementoclasts of Inflamed Human Periodontium
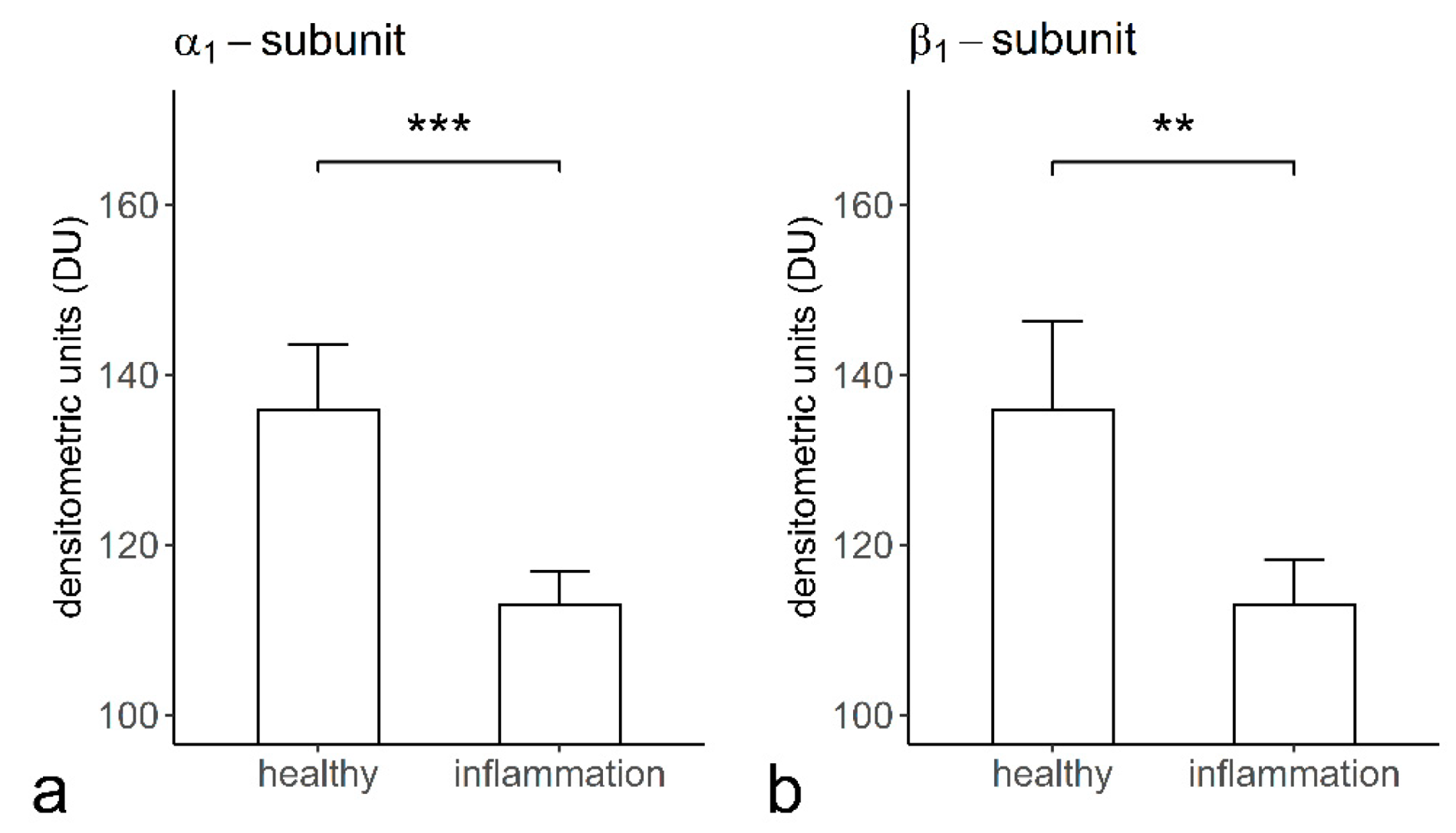
2.5. The Staining Intensities of α1- and β1-Subunits of sGC in Cementoclasts of the Healthy and Inflamed Human Periodontium
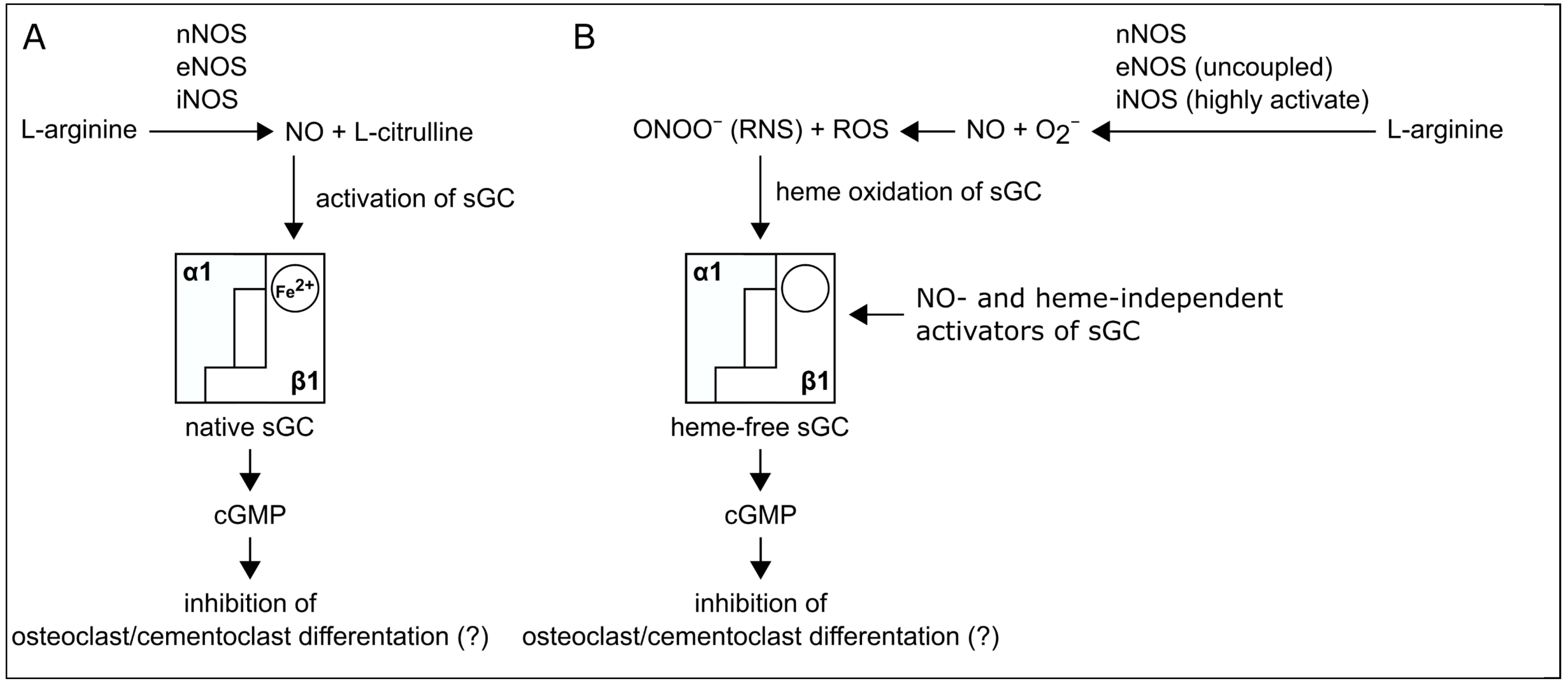
3. Discussion
4. Materials and Methods
4.1. Tissue Sample Collection
4.2. Histological Evaluation of the Inflammatory State
4.3. The Specificity of Antibodies against Human α1- and β1-Subunits of sGC
4.4. Immunohistochemical Avidin-Biotin-Peroxidase Complex Method
4.5. Immunofluorescence Double Staining Method
4.6. Controls
4.7. Quantification of Staining Intensities of α1- and β1-Subunits and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HE | Hematoxylin and Eosin |
| NO | Nitric Oxide |
| NOS | Nitric Oxide Synthase |
| sGC | soluble Guanylyl Cyclase |
| cGMP | cyclic Guanosine Monophosphate |
| GTP | Guanosine-5′-Triphosphate |
| PKG | protein kinase G |
| CNG | cyclic nucleotide-gated channel |
| PDE | phosphodiesterase |
| PDL | periodontal ligament |
| MCT | Mast Cell Tryptase |
| HLA-DR | Human Leukocyte Antigen-DR isotype |
| CD68 | Cluster of Differentiation 68 |
| DRAQ5 | Deep Red Anthraquinone 5 |
| ROS | Reactive Oxygen Species |
| RNS | Reactive Nitrogen Species |
References
- Fleming, I. Molecular mechanisms underlying the activation of eNOS. Pflug. Arch. 2010, 459, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friebe, A.; Koesling, D. Regulation of nitric oxide-sensitive guanylyl cyclase. Circ. Res. 2003, 93, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Murad, F. Shattuck Lecture: Nitric oxide and cyclic GMP in cell signaling and drug development. N. Engl. J. Med. 2006, 355, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Stasch, J.P.; Schmidt, P.; Alonso-Alija, C.; Apeler, H.; Dembowsky, K.; Haerter, M.; Heil, M.; Minuth, T.; Perzborn, E.; Pleiss, U.; et al. NO- and haem-independent activation of soluble guanylyl cyclase: Molecular basis and cardiovascular implications of a new pharmacological principle. Br. J. Pharmacol. 2002, 136, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Stasch, J.P.; Schmidt, P.M.; Nedvetsky, P.I.; Nedvetskaya, T.Y.; HS, A.K.; Meurer, S.; Deile, M.; Taye, A.; Knorr, A.; Lapp, H.; et al. Targeting the heme-oxidized nitric oxide receptor for selective vasodilatation of diseased blood vessels. J. Clin. Investig. 2006, 116, 2552–2561. [Google Scholar] [CrossRef] [Green Version]
- Russwurm, M.; Behrends, S.; Harteneck, C.; Koesling, D. Functional properties of a naturally occurring isoform of soluble guanylyl cyclase. Biochem. J. 1998, 335, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabel, U.; Weeger, M.; La, M.; Schmidt, H.H. Human soluble guanylate cyclase: Functional expression and revised isoenzyme family. Biochem. J. 1998, 335, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Evgenov, O.V.; Pacher, P.; Schmidt, P.M.; Haskó, G.; Schmidt, H.H.; Stasch, J.P. NO-independent stimulators and activators of soluble guanylate cyclase: Discovery and therapeutic potential. Nat. Rev. Drug Discov. 2006, 5, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Friebe, A.; Koesling, D. The function of NO-sensitive guanylyl cyclase: What we can learn from genetic mouse models. Nitric Oxide 2009, 21, 149–156. [Google Scholar] [CrossRef]
- Kalyanaraman, H.; Ramdani, G.; Joshua, J.; Schall, N.; Boss, G.R.; Cory, E.; Sah, R.L.; Casteel, D.E.; Pilz, R.B. A novel, direct NO donor regulates osteoblast and osteoclast functions and increases bone mass in ovariectomized mice. J. Bone Miner. Res. 2017, 32, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, H.; Schall, N.; Pilz, R.B. Nitric oxide and cyclic GMP functions in bone. Nitric Oxide 2018, 76, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Taubman, M.A.; Valverde, P.; Han, X.; Kawai, T. Immune response: The key to bone resorption in periodontal disease. J. Periodontol. 2005, 76, 2033–2041. [Google Scholar] [CrossRef]
- Cochran, D.L. Inflammation and bone loss in periodontal disease. J. Periodontol. 2008, 79, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Belibasakis, G.N.; Bostanci, N. The RANKL-OPG system in clinical periodontology. J. Clin. Periodontol. 2012, 39, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sczepanik, F.S.C.; Grossi, M.L.; Casati, M.; Goldberg, M.; Glogauer, M.; Fine, N.; Tenenbaum, H.C. Periodontitis is an inflammatory disease of oxidative stress: We should treat it that way. Periodontol. 2000 2020, 84, 45–68. [Google Scholar] [CrossRef]
- Arana-Chavez, V.E.; Bradaschia-Correa, V. Clastic cells: Mineralized tissue resorption in health and disease. Int. J. Biochem. Cell Biol. 2009, 41, 446–450. [Google Scholar] [CrossRef]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]
- Blair, H.C.; Teitelbaum, S.L.; Ghiselli, R.; Gluck, S. Osteoclastic bone resorption by a polarized vacuolar proton pump. Science 1989, 245, 855–857. [Google Scholar] [CrossRef]
- Horton, M.A.; Dorey, E.L.; Nesbitt, S.A.; Samanen, J.; Ali, F.E.; Stadel, J.M.; Nichols, A.; Greig, R.; Helfrich, M.H. Modulation of vitronectin receptor-mediated osteoclast adhesion by Arg-Gly-Asp peptide analogs: A structure-function analysis. J. Bone Miner. Res. 1993, 8, 239–247. [Google Scholar] [CrossRef]
- Novack, D.V.; Teitelbaum, S.L. The osteoclast: Friend or foe? Annu. Rev. Pathol. 2008, 3, 457–484. [Google Scholar] [CrossRef] [PubMed]
- Wheal, B.D.; Beach, R.J.; Tanabe, N.; Dixon, S.J.; Sims, S.M. Subcellular elevation of cytosolic free calcium is required for osteoclast migration. J. Bone Miner. Res. 2014, 29, 725–734. [Google Scholar] [CrossRef] [PubMed]
- van’t Hof, R.J.; Macphee, J.; Libouban, H.; Helfrich, M.H.; Ralston, S.H. Regulation of bone mass and bone turnover by neuronal nitric oxide synthase. Endocrinology 2004, 145, 5068–5074. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yu, X.; Collin-Osdoby, P.; Osdoby, P. RANKL stimulates inducible nitric-oxide synthase expression and nitric oxide production in developing osteoclasts. An autocrine negative feedback mechanism triggered by RANKL-induced interferon-beta via NF-kappaB that restrains osteoclastogenesis and bone resorption. J. Biol. Chem. 2006, 281, 15809–15820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowik, C.W.; Nibbering, P.H.; van de Ruit, M.; Papapoulos, S.E. Inducible production of nitric oxide in osteoblast-like cells and in fetal mouse bone explants is associated with suppression of osteoclastic bone resorption. J. Clin. Investig. 1994, 93, 1465–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.S.; Williams, J.P.; Jordan, S.E.; Cornwell, T.; Blair, H.C. Nitric oxide regulation of cGMP production in osteoclasts. J. Cell. Biochem. 1999, 73, 478–487. [Google Scholar] [CrossRef]
- Wang, J.W.; Yeh, C.B.; Chou, S.J.; Lu, K.C.; Chu, T.H.; Chen, W.Y.; Chien, J.L.; Yen, M.H.; Chen, T.H.; Shyu, J.F. YC-1 alleviates bone loss in ovariectomized rats by inhibiting bone resorption and inducing extrinsic apoptosis in osteoclasts. J. Bone Miner. Metab. 2018, 36, 508–518. [Google Scholar] [CrossRef]
- Martin, E.; Lee, Y.C.; Murad, F. YC-1 activation of human soluble guanylyl cyclase has both heme-dependent and heme-independent components. Proc. Natl. Acad. Sci. USA 2001, 98, 12938–12942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matejka, M.; Partyka, L.; Ulm, C.; Solar, P.; Sinzinger, H. Nitric oxide synthesis is increased in periodontal disease. J. Periodontal. Res. 1998, 33, 517–518. [Google Scholar] [CrossRef] [PubMed]
- Lappin, D.F.; Kjeldsen, M.; Sander, L.; Kinane, D.F. Inducible nitric oxide synthase expression in periodontitis. J. Periodontal. Res. 2000, 35, 369–373. [Google Scholar] [CrossRef]
- Hirose, M.; Ishihara, K.; Saito, A.; Nakagawa, T.; Yamada, S.; Okuda, K. Expression of cytokines and inducible nitric oxide synthase in inflamed gingival tissue. J. Periodontol. 2001, 72, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Shibata, K.; Warbington, M.L.; Gordon, B.J.; Kurihara, H.; Van Dyke, T.E. Nitric oxide synthase activity in neutrophils from patients with localized aggressive periodontitis. J. Periodontol. 2001, 72, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Gyurko, R.; Shoji, H.; Battaglino, R.A.; Boustany, G.; Gibson, F.C., III; Genco, C.A.; Stashenko, P.; Van Dyke, T.E. Inducible nitric oxide synthase mediates bone development and P. gingivalis-induced alveolar bone loss. Bone 2005, 36, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Fukada, S.Y.; Silva, T.A.; Saconato, I.F.; Garlet, G.P.; Avila-Campos, M.J.; Silva, J.S.; Cunha, F.Q. iNOS-derived nitric oxide modulates infection-stimulated bone loss. J. Dent. Res. 2008, 87, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- Machigashira, M.; Matsuyama, T.; Matsuguchi, T. Oxidative stress causes alveolar bone loss in metabolic syndrome model mice with type 2 diabetes. J. Periodontal. Res. 2009, 44, 43–51. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Xiao, E.; Graves, D.T. Diabetes mellitus related bone metabolism and periodontal disease. Int. J. Oral Sci. 2015, 7, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkmaz, Y.; Baumann, M.A.; Schroder, H.; Behrends, S.; Addicks, K.; Raab, W.H.; Bloch, W. Localization of the NO-cGMP signaling pathway molecules, NOS III-phosphorylation sites, ERK1/2, and Akt/PKB in osteoclasts. J. Periodontol. 2004, 75, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, Y.; Roggendorf, H.C.; Siefer, O.G.; Seehawer, J.; Imhof, T.; Plomann, M.; Bloch, W.; Friebe, A.; Huebbers, C.U. Downregulation of the α1- and β1-subunit of sGC in arterial smooth muscle cells of OPSCC is HPV-Independent. J. Dent. Res. 2018, 97, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Friebe, A.; Mergia, E.; Dangel, O.; Lange, A.; Koesling, D. Fatal gastrointestinal obstruction and hypertension in mice lacking nitric oxide-sensitive guanylyl cyclase. Proc. Natl. Acad. Sci. USA 2007, 104, 7699–7704. [Google Scholar] [CrossRef] [Green Version]
- Brezniak, N.; Wasserstein, A. Orthodontically induced inflammatory root resorption. Part I: The basic science aspects. Angle Orthod. 2002, 72, 175–179. [Google Scholar] [CrossRef]
- Alfuriji, S.; Alhazmi, N.; Alhamlan, N.; Al-Ehaideb, A.; Alruwaithi, M.; Alkatheeri, N.; Geevarghese, A. The effect of orthodontic therapy on periodontal health: A review of the literature. Int. J. Dent. 2014, 2014, 585048. [Google Scholar] [CrossRef] [PubMed]
- Yaroslavskiy, B.B.; Li, Y.; Ferguson, D.J.; Kalla, S.E.; Oakley, J.I.; Blair, H.C. Autocrine and paracrine nitric oxide regulate attachment of human osteoclasts. J. Cell. Biochem. 2004, 91, 962–972. [Google Scholar] [CrossRef]
- Yaroslavskiy, B.B.; Zhang, Y.; Kalla, S.E.; Palacios, V.G.; Sharrow, A.C.; Li, Y.; Zaidi, M.; Wu, C.; Blair, H.C. NO-dependent osteoclast motility: Reliance on cGMP-dependent protein kinase I and VASP. J. Cell Sci. 2005, 118, 5479–5487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.M.; Kraehling, J.R.; Eitner, F.; Bénardeau, A.; Sandner, P. The impact of the nitric oxide (NO)/Soluble Guanylyl Cyclase (sGC) signaling cascade on kidney health and disease: A preclinical perspective. Int. J. Mol. Sci. 2018, 19, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshua, J.; Schwaerzer, G.K.; Kalyanaraman, H.; Cory, E.; Sah, R.L.; Li, M.; Vaida, F.; Boss, G.R.; Pilz, R.B. Soluble guanylate cyclase as a novel treatment target for osteoporosis. Endocrinology 2014, 155, 4720–4730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahaman, M.M.; Nguyen, A.T.; Miller, M.P.; Hahn, S.A.; Sparacino-Watkins, C.; Jobbagy, S.; Carew, N.T.; Cantu-Medellin, N.; Wood, K.C.; Baty, C.J.; et al. Cytochrome b5 reductase 3 modulates soluble guanylate cyclase redox state and cGMP signaling. Circ. Res. 2017, 121, 137–148. [Google Scholar] [CrossRef]
- Meurer, S.; Pioch, S.; Pabst, T.; Opitz, N.; Schmidt, P.M.; Beckhaus, T.; Wagner, K.; Matt, S.; Gegenbauer, K.; Geschka, S.; et al. Nitric oxide-independent vasodilator rescues heme-oxidized soluble guanylate cyclase from proteasomal degradation. Circ. Res. 2009, 105, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, Y.; Lang, H.; Beikler, T.; Cho, B.; Behrends, S.; Bloch, W.; Addicks, K.; Raab, W.H. Irreversible inflammation is associated with decreased levels of the α1-, β1- and α2-subunit of sGC in human odontoblasts. J. Dent. Res. 2011, 90, 517–522. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korkmaz, Y.; Puladi, B.; Galler, K.; Kämmerer, P.W.; Schröder, A.; Gölz, L.; Sparwasser, T.; Bloch, W.; Friebe, A.; Deschner, J. Inflammation in the Human Periodontium Induces Downregulation of the α1- and β1-Subunits of the sGC in Cementoclasts. Int. J. Mol. Sci. 2021, 22, 539. https://doi.org/10.3390/ijms22020539
Korkmaz Y, Puladi B, Galler K, Kämmerer PW, Schröder A, Gölz L, Sparwasser T, Bloch W, Friebe A, Deschner J. Inflammation in the Human Periodontium Induces Downregulation of the α1- and β1-Subunits of the sGC in Cementoclasts. International Journal of Molecular Sciences. 2021; 22(2):539. https://doi.org/10.3390/ijms22020539
Chicago/Turabian StyleKorkmaz, Yüksel, Behrus Puladi, Kerstin Galler, Peer W. Kämmerer, Agnes Schröder, Lina Gölz, Tim Sparwasser, Wilhelm Bloch, Andreas Friebe, and James Deschner. 2021. "Inflammation in the Human Periodontium Induces Downregulation of the α1- and β1-Subunits of the sGC in Cementoclasts" International Journal of Molecular Sciences 22, no. 2: 539. https://doi.org/10.3390/ijms22020539