
Parasite Survival and Disease Persistence in Cystic Fibrosis, Schistosomiasis and Pathogenic Bacterial Diseases: A Role for Universal Stress Proteins?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
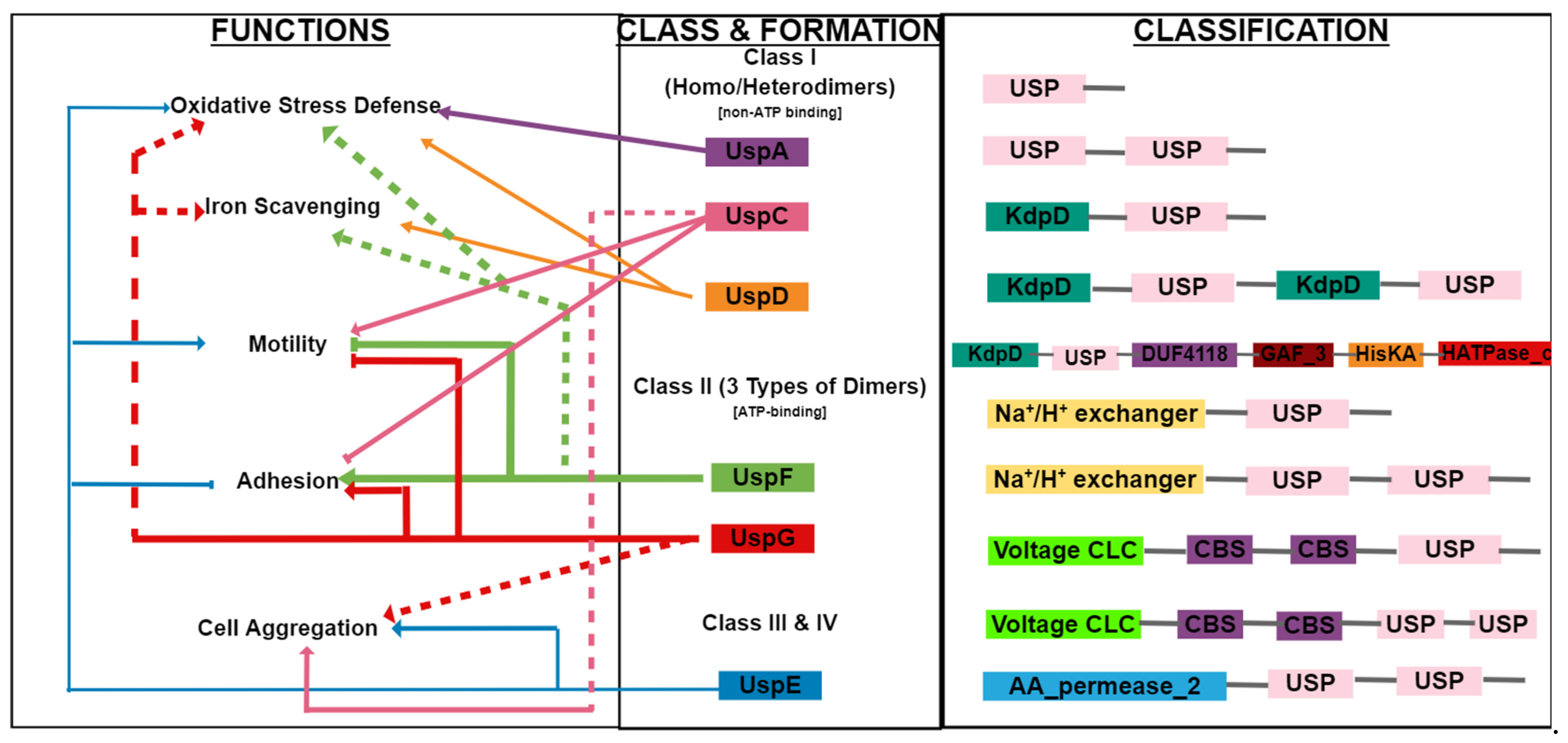
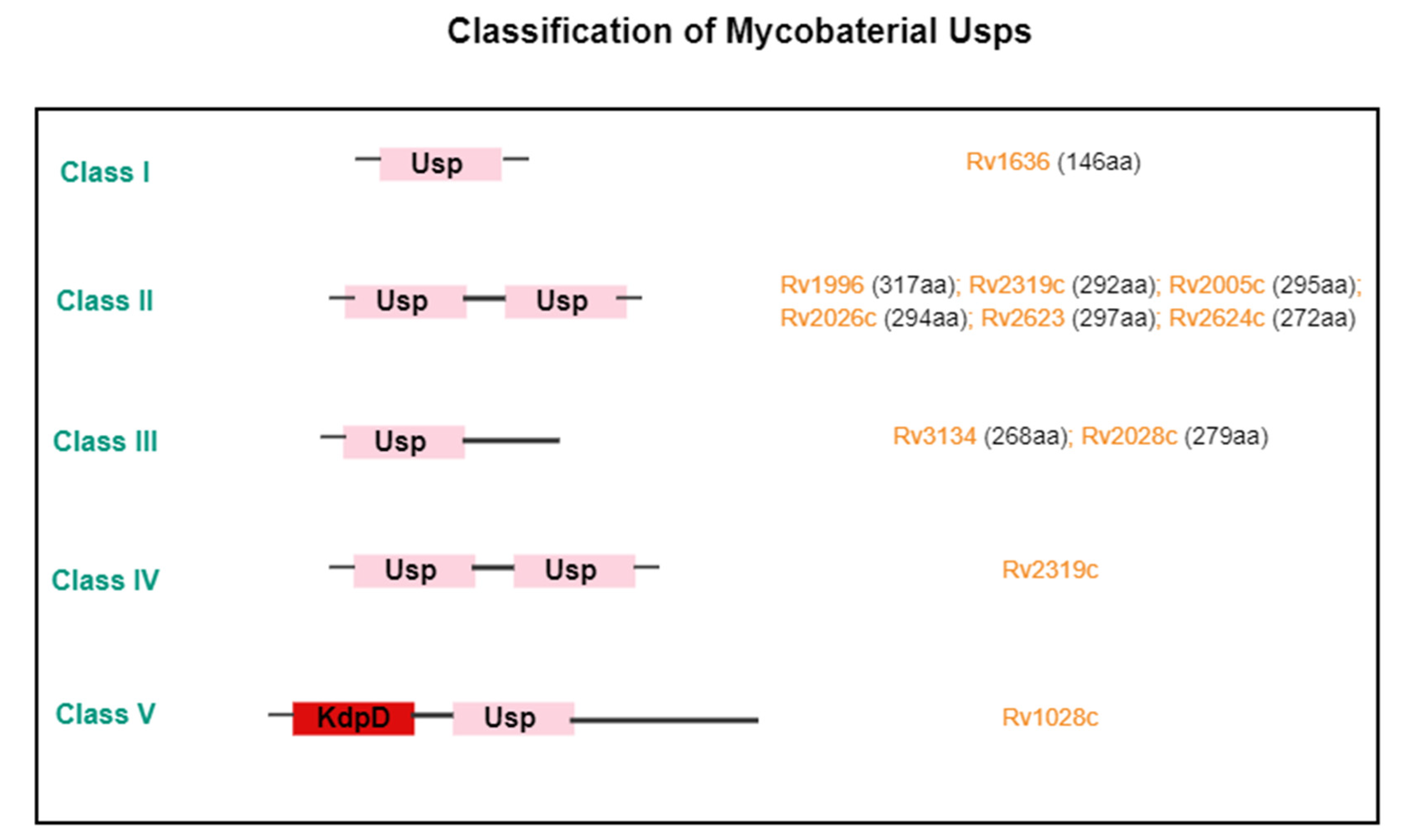
2. Structure of the USP Family
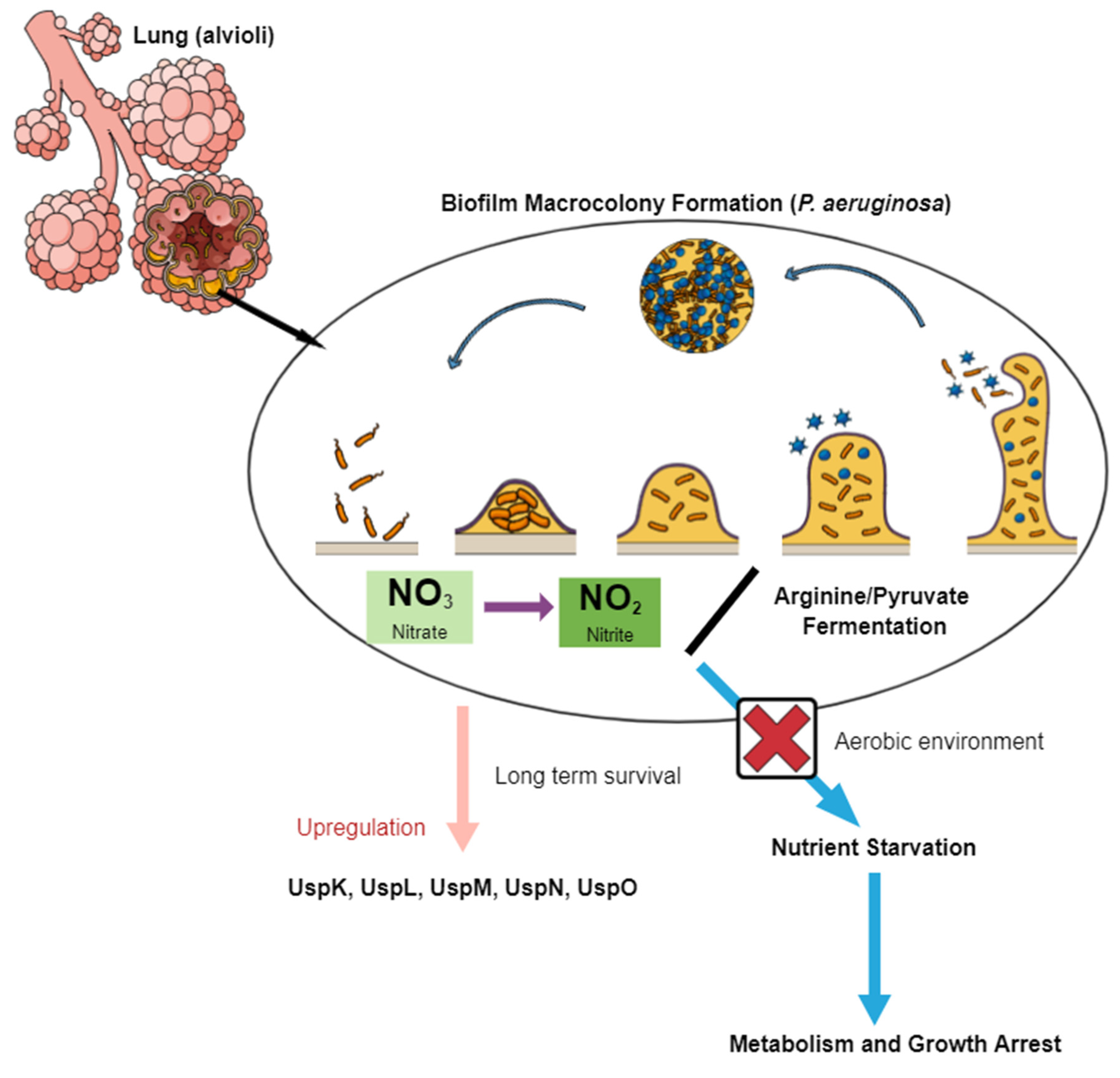
3. USPs in Cystic Fibrosis
4. USPs in Schistosomiasis
5. USPs of Pathogenic Bacteria
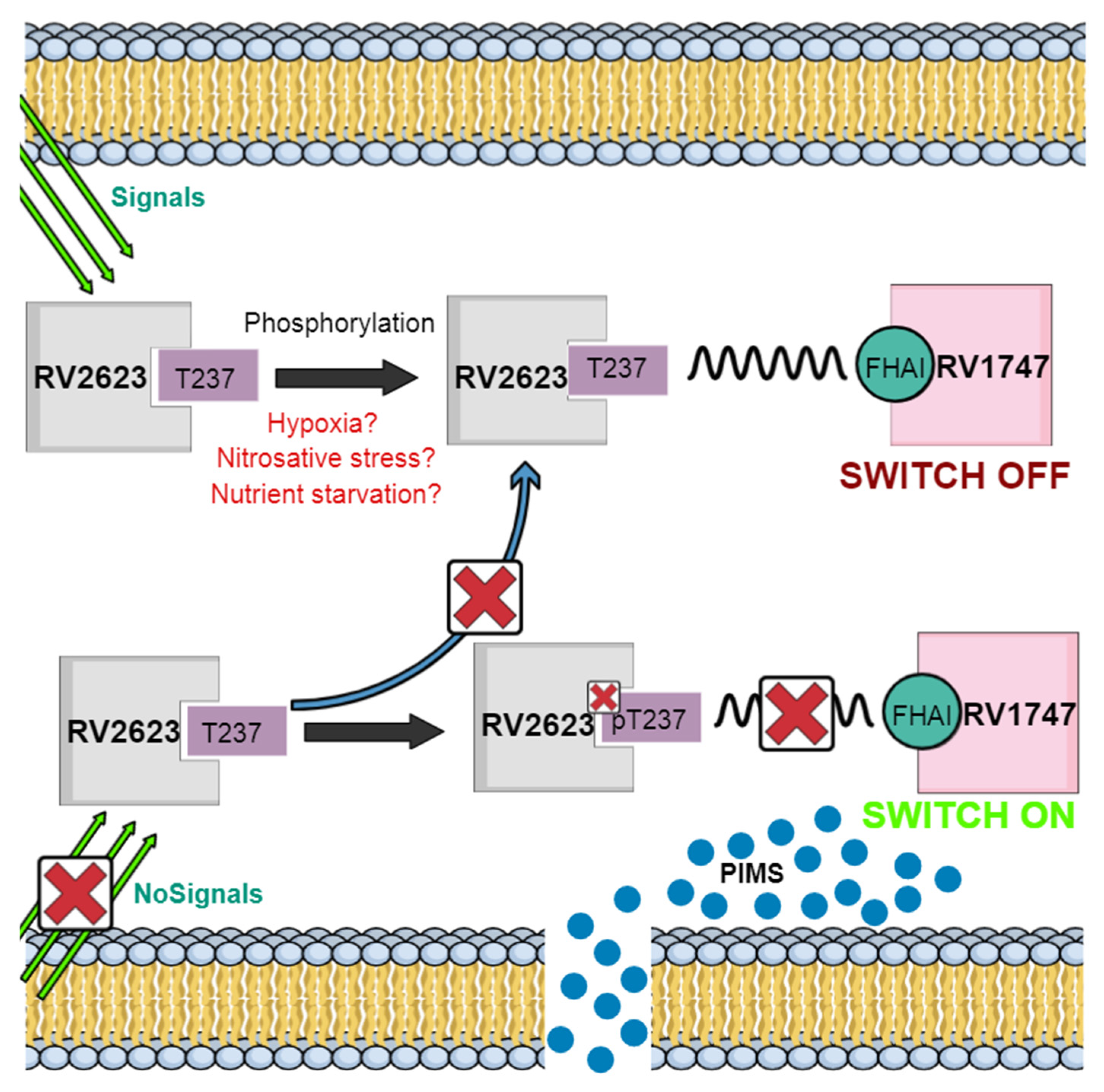
5.1. Tuberculosis
5.2. Salmonella
5.3. Porphyromonas gingivalis
5.4. Staphylococcus aureus
5.5. Usps of Other Pathogenic Bacteria
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matarredona, L.; Camacho, M.; Zafrilla, B.; Bonete, M.-J.; Esclapez, J. The role of stress proteins in Haloarchaea and their adaptive response to environmental shifts. Biomolecules 2020, 10, 1390. [Google Scholar] [CrossRef] [PubMed]
- Elhosseiny, N.M.; Amin, M.A.; Yassin, A.; Attia, A.S. Acinetobacter baumannii Universal Stress Protein A plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int. J. Med. Microbiol. 2015, 305, 114–123. [Google Scholar] [CrossRef]
- Chi, Y.H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, S.D.; Bin Bae, S.; Paeng, S.K.; Chae, H.B.; et al. The physiological functions of Universal Stress Proteins and their molecular mechanism to protect plants from environmental stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Q.-J.; Han, Y.-X.; Shi, Y.-J.; Huang, H.-Q.; Fang, Z.-G.; Hu, Y.-H. Universal Stress Proteins contribute Edwardsiella piscicida adversity resistance and pathogenicity and promote blocking host immune response. Fish Shellfish Immunol. 2019, 95, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Havis, S.; Bodunrin, A.; Rangel, J.; Zimmerer, R.; Murphy, J.; Storey, J.D.; Duong, T.D.; Mistretta, B.; Gunaratne, P.; Widger, W.R.; et al. A Universal Stress Protein that controls bacterial stress survival in Micrococcus luteus. J. Bacteriol. 2019, 201, e00497-19. [Google Scholar] [CrossRef]
- Tkaczuk, K.L.; Shumilin, I.A.; Chruszcz, M.; Evdokimova, E.; Savchenko, A.; Minor, W. Structural and functional insight into the Universal Stress Protein family. Evol. Appl. 2013, 6, 434–449. [Google Scholar] [CrossRef]
- Kvint, K.; Nachin, L.; Diez, A.; Nyström, T. The bacterial Universal Stress Protein: Function and regulation. Curr. Opin. Microbiol. 2003, 6, 140–145. [Google Scholar] [CrossRef]
- Vollmer, A.C.; Bark, S.J. Twenty-five years of investigating the Universal Stress Protein: Function, structure, and applications. Adv. Appl. Microbiol. 2018, 102, 1–36. [Google Scholar] [CrossRef]
- Nyström, T.; Neidhardt, F.C. Expression and role of the Universal Stress Protein, UspA, of Escherichia coli during growth arrest. Mol. Microbiol. 1994, 11, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Freestone, P.; Nyström, T.; Trinei, M.; Norris, V. The Universal Stress Protein, UspA, of Escherichia coli is phosphorylated in response to stasis. J. Mol. Biol. 1997, 274, 318–324. [Google Scholar] [CrossRef]
- Persson, O.; Valadi, A.; Nyström, T.; Farewell, A. Metabolic control of the Escherichia coli Universal Stress Protein response through fructose-6-phosphate. Mol. Microbiol. 2007, 65, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Diez, A.; Gustavsson, N.; Nyström, T. The Universal Stress Protein A of Escherichia coli is required for resistance to DNA damaging agents and is regulated by a RecA/FtsK-dependent regulatory pathway. Mol. Microbiol. 2002, 36, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Nyström, T.; Neidhardt, F.C. Isolation and properties of a mutant of Escherichia coli with an insertional inactivation of the uspA gene, which encodes a Universal Stress Protein. J. Bacteriol. 1993, 175, 3949–3956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, D. The Universal Stress Proteins of Bacteria. Ph.D. Thesis, Imperial College of London, London, UK, 2011. Available online: https://spiral.imperial.ac.uk/handle/10044/1/6946 (accessed on 15 April 2021).
- Nachin, L.; Nannmark, U.; Nyström, T. Differential roles of the Universal Stress Proteins of Escherichia coli in oxidative stress resistance, adhesion, and motility. J. Bacteriol. 2005, 187, 6265–6272. [Google Scholar] [CrossRef] [Green Version]
- Nyström, T.; Neidhardt, F.C. Effects of overproducing the Universal Stress Protein, UspA, in Escherichia coli K-12. J. Bacteriol. 1996, 178, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-T.; Karavolos, M.H.; Bulmer, D.M.; Allaoui, A.; Hormaeche, R.D.; Lee, J.J.; Khan, C.A. Role of the Universal Stress Protein UspA of Salmonella in growth arrest, stress and virulence. Microb. Pathog. 2007, 42, 2–10. [Google Scholar] [CrossRef]
- Wang, X.-F.; Su, J.; Yang, N.; Zhang, H.; Cao, X.-Y.; Kang, J.-F. Functional characterization of selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes 2017, 8, 224. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Goo, E.; Kang, Y.; Kim, J.; Hwang, I. Regulation of Universal Stress Protein genes by quorum sensing and RpoS in Burkholderia glumae. J. Bacteriol. 2011, 194, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Ziaf, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting Universal Stress Protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Van Der Does, C.; Albers, S.-V. SaUspA, the Universal Stress Protein of Sulfolobus acidocaldarius stimulates the activity of the PP2A phosphatase and is involved in growth at high salinity. Front. Microbiol. 2020, 11, 598821. [Google Scholar] [CrossRef]
- Masamba, P.; Adenowo, A.F.; Oyinloye, B.E.; Kappo, A.P. Universal Stress Proteins as new targets for environmental and therapeutic interventions of schistosomiasis. Int. J. Environ. Res. Public Health 2016, 13, 972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Guo, J.; Jin, X.; Kim, J.-S.; Ji, Y.; Fan, S.; Ha, N.-C.; Quan, C.-S. Crystal structure and functional implications of the tandem-type Universal Stress Protein UspE from Escherichia coli. BMC Struct. Biol. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, R.; Williams, H. Universal Stress Proteins and Mycobacterium tuberculosis. Res. Microbiol. 2003, 154, 387–392. [Google Scholar] [CrossRef]
- Hingley-Wilson, S.; Lougheed, K.; Ferguson, K.; Leiva, S.; Williams, H. Individual mycobacterium tuberculosis Universal Stress Protein homologues are dispensable in vitro. Tuberculosis 2010, 90, 236–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.; Jung, K. Biochemical properties of UspG, a Universal Stress Protein of Escherichia coli. Biochemistry 2006, 45, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Adolph, R.S.; Gopalakrishnapai, J.; Kleinboelting, S.; Emmerich, C.; Steegborn, C.; Visweswariah, S.S. A Universal Stress Protein (USP) in mycobacteria binds cAMP. J. Biol. Chem. 2015, 290, 12731–12743. [Google Scholar] [CrossRef] [Green Version]
- Udawat, P.; Jha, R.K.; Sinha, D.; Mishra, A.; Jha, B. Overexpression of a cytosolic abiotic stress responsive Universal Stress Protein (SbUSP) mitigates salt and osmotic stress in transgenic tobacco plants. Front. Plant Sci. 2016, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- Heermann, R.; Weber, A.; Mayer, B.; Ott, M.; Hauser, E.; Gabriel, G.; Pirch, T.; Jung, K. The Universal Stress Protein UspC scaffolds the KdpD/KdpE signaling cascade of Escherichia coli under salt stress. J. Mol. Biol. 2009, 386, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Anantharaman, V.; Koonin, E.V. Monophyly of class I aminoacyl tRNA synthetase, USPA, ETFP, photolyase, and PP-ATPase nucleotide-binding domains: Implications for protein evolution in the RNA world. Proteins Struct. Funct. Bioinform. 2002, 48, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bangera, M.; Panigrahi, R.; Sagurthi, S.; Savithri, H.; Murthy, M. Structural and functional analysis of two Universal Stress Proteins YdaA and YnaF from Salmonella typhimurium: Possible roles in microbial stress tolerance. J. Struct. Biol. 2015, 189, 238–250. [Google Scholar] [CrossRef]
- Jung, Y.J.; Melencion, S.M.B.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T.; Yun, D.-J.; Chi, Y.H.; Lee, S.Y. Universal Stress Protein exhibits a redox-dependent chaperone function in arabidopsis and enhances plant tolerance to heat shock and oxidative stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokry, A.M.; Al-Karim, S.; Ramadan, A.; Gadallah, N.; Al Attas, S.G.; Sabir, J.S.; Hassan, S.M.; Madkour, L.; Bressan, R.; Mahfouz, M.M.; et al. Detection of a Usp-like gene in Calotropis procera plant from the de novo assembled genome contigs of the high-throughput sequencing dataset. Comptes Rendus Biol. 2014, 337, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Melencion, S.M.B.; Chi, Y.H.; Pham, T.T.; Paeng, S.K.; Wi, S.D.; Lee, C.; Ryu, S.W.; Koo, S.S.; Lee, S.Y. RNA chaperone function of a Universal Stress Protein in arabidopsis confers enhanced cold stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 2546. [Google Scholar] [CrossRef] [Green Version]
- Isokpehi, R.D.; Mahmud, O.; Mbah, A.N.; Simmons, S.S.; Avelar, L.; Rajnarayanan, R.V.; Udensi, U.K.; Ayensu, W.K.; Cohly, H.H.; Brown, S.D.; et al. Developmental regulation of genes encoding Universal Stress Proteins in Schistosoma mansoni. Gene Regul. Syst. Biol. 2011, 5, GRSB-S7491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegele, D.A. Universal Stress Proteins in Escherichia coli. J. Bacteriol. 2005, 187, 6253–6254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, M.C.; McKay, D.B. Structure of the Universal Stress Protein of Haemophilus influenzae. Structure 2001, 9, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Zarembinski, T.I.; Hung, L.-W.; Mueller-Dieckmann, H.-J.; Kim, K.-K.; Yokota, H.; Kim, R.; Kim, S.-H. Structure-based assignment of the biochemical function of a hypothetical protein: A test case of structural genomics. Proc. Natl. Acad. Sci. USA 1998, 95, 15189–15193. [Google Scholar] [CrossRef] [Green Version]
- Isokpehi, R.D.; Mahmud, O.; Awofolu, O.R.; Mbah, A.N. Inferences on the biochemical and environmental regulation of Universal Stress Proteins from Schistosomiasis parasites. Adv. Appl. Bioinform. Chem. 2013, 6, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Davis, P.B. Cystic fibrosis since 1938. Am. J. Respir. Crit. Care Med. 2006, 173, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Knowles, M.R.; Durie, P.R. What is cystic fibrosis? N. Engl. J. Med. 2002, 347, 439–442. [Google Scholar] [CrossRef]
- Döring, G.; Gulbins, E. Cystic fibrosis and innate immunity: How chloride channel mutations provoke lung disease. Cell. Microbiol. 2009, 11, 208–216. [Google Scholar] [CrossRef]
- Yang, L.; Jelsbak, L.; Molin, S. Microbial ecology and adaptation in cystic fibrosis airways. Environ. Microbiol. 2011, 13, 1682–1689. [Google Scholar] [CrossRef]
- Millette, G.; Langlois, J.-P.; Brouillette, E.; Frost, E.H.; Cantin, A.M.; Malouin, F. despite antagonism in vitro, Pseudomonas aeruginosa enhances Staphylococcus aureus colonization in a murine lung infection model. Front. Microbiol. 2019, 10, 2880. [Google Scholar] [CrossRef] [Green Version]
- Sass, A.; Schmerk, C.; Agnoli, K.; Norville, P.J.; Eberl, L.; Valvano, M.; Mahenthiralingam, E. The unexpected discovery of a novel low-oxygen-activated locus for the anoxic persistence of Burkholderia cenocepacia. ISME J. 2013, 7, 1568–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, K.; Boes, N.; Eschbach, M.; Jaensch, L.; Wehland, J.; Bjarnsholt, T.; Givskov, M.; Hentzer, M.; Schobert, M. Anaerobic survival of Pseudomonas aeruginosa by pyruvate fermentation requires an usp-type stress protein. J. Bacteriol. 2006, 188, 659–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm. Pharmacol. Ther. 2008, 21, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J. Clin. Investig. 2002, 109, 317–325. [Google Scholar] [CrossRef]
- Borriello, G.; Werner, E.; Roe, F.; Kim, A.M.; Ehrlich, G.D.; Stewart, P.S. Oxygen limitation contributes to antibiotic tolerance of Pseudomonas aeruginosa in biofilms. Antimicrob. Agents Chemother. 2004, 48, 2659–2664. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell 2002, 3, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Boes, N.; Schreiber, K.; Härtig, E.; Jaensch, L.; Schobert, M. The Pseudomonas aeruginosa Universal Stress Protein PA4352 is essential for surviving anaerobic energy stress. J. Bacteriol. 2006, 188, 6529–6538. [Google Scholar] [CrossRef] [Green Version]
- Boes, N.; Schreiber, K.; Schobert, M. SpoT-Triggered stringent response controls usp gene expression in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7189–7199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schobert, M.; Jahn, D. Anaerobic physiology of Pseudomonas aeruginosa in the cystic fibrosis lung. Int. J. Med. Microbiol. 2010, 300, 549–556. [Google Scholar] [CrossRef]
- O’Connor, A.; Berisio, R.; Lucey, M.; Schaffer, K.; McClean, S. A Universal Stress Protein upregulated by hypoxia may contribute to chronic lung colonisation and intramacrophage survival in cystic fibrosis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Courtney, J.; Dunbar, K.; McDowell, A.; Moore, J.; Warke, T.; Stevenson, M.; Elborn, J. Clinical outcome of Burkholderia cepacia complex infection in cystic fibrosis adults. J. Cyst. Fibros. 2004, 3, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.; Dodd, M.; Webb, A. Burkholderia cepacia: Current clinical issues, environmental controversies and ethical dilemmas. Eur. Respir. J. 2001, 17, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Cullen, L.; O’Connor, A.; McCormack, S.; Owens, R.; Holt, G.S.; Collins, C.; Callaghan, M.; Doyle, S.; Smith, D.; Schaffer, K.; et al. The involvement of the low-oxygen-activated locus of Burkholderia cenocepacia in adaptation during cystic fibrosis infection. Sci. Rep. 2018, 8, 13386. [Google Scholar] [CrossRef]
- Goerke, C.; Wolz, C. Regulatory and genomic plasticity of Staphylococcus aureus during persistent colonization and infection. Int. J. Med. Microbiol. 2004, 294, 195–202. [Google Scholar] [CrossRef]
- Treffon, J.; Block, D.; Moche, M.; Reiss, S.; Fuchs, S.; Engelmann, S.; Becher, D.; Langhanki, L.; Mellmann, A.; Peters, G.; et al. Adaptation of Staphylococcus aureus to airway environments in patients with cystic fibrosis by upregulation of superoxide dismutase M and iron-scavenging proteins. J. Infect. Dis. 2018, 217, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, D.O.; Taylor, D.; Skerrett, S.J.; Rubens, C.E. Changes in the Staphylococcus aureus transcriptome during early adaptation to the lung. PLoS ONE 2012, 7, e41329. [Google Scholar] [CrossRef] [Green Version]
- Adenowo, A.F.; Oyinloye, B.E.; Ogunyinka, B.I.; Kappo, A.P. Impact of human schistosomiasis in sub-Saharan Africa. Braz. J. Infect. Dis. 2015, 19, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Berriman, M.; Haas, B.J.; LoVerde, P.T.; Wilson, R.A.; Dillon, G.; Cerqueira, G.C.; Mashiyama, S.T.; Al-Lazikani, B.; Andrade, L.F.; Ashton, P.D.; et al. The genome of the blood fluke Schistosoma mansoni. Nature 2009, 460, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, U.K.; Bender, R.C.; Bayne, C.J. Killing of Schistosoma mansoni sporocysts by hemocytes from resistant Biomphalaria glabrata: Role of reactive oxygen species. J. Parasitol. 2001, 87, 292. [Google Scholar] [CrossRef]
- Altaf, M.; Miller, C.H.; Bellows, D.S.; O’Toole, R. Evaluation of the mycobacterium smegmatis and BCG models for the discovery of mycobacterium tuberculosis inhibitors. Tuberculosis 2010, 90, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Bourzac, K. Infectious disease: Beating the big three. Nature 2014, 507, S4–S7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principi, N.; Esposito, S. The present and future of tuberculosis vaccinations. Tuberculosis 2015, 95, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Sherman, D.R.; Voskuil, M.; Schnappinger, D.; Liao, R.; Harrell, M.I.; Schoolnik, G.K. Regulation of the mycobacterium tuberculosis hypoxic response gene encoding crystallin. Proc. Natl. Acad. Sci. USA 2001, 98, 7534–7539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustad, T.R.; Sherrid, A.M.; Minch, K.J.; Sherman, D.R. Hypoxia: A window into mycobacterium tuberculosislatency. Cell. Microbiol. 2009, 11, 1151–1159. [Google Scholar] [CrossRef]
- Selvaraj, S.; Sambandam, V.; Sardar, D.; Anishetty, S. In silico analysis of DosR regulon proteins of mycobacterium tuberculosis. Gene 2012, 506, 233–241. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, S.; Liu, Y.; Zhang, X.; Wang, W.; Li, C. Potential role for Rv2026c and Rv2421c specific antibody responses in diagnosing active tuberculosis. Clin. Chim. Acta 2018, 487, 369–376. [Google Scholar] [CrossRef]
- Jacobs, A.J.; Mongkolsapaya, J.; Screaton, G.; McShane, H.; Wilkinson, R.J. Antibodies and tuberculosis. Tuberculosis 2016, 101, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Javid, B. Antibodies and tuberculosis: Finally coming of age? Nat. Rev. Immunol. 2018, 18, 591–596. [Google Scholar] [CrossRef]
- Provvedi, R.; Boldrin, F.; Falciani, F.; Palù, G.; Manganelli, R. Global transcriptional response to vancomycin in Mycobacterium tuberculosis. Microbiology 2009, 155, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Drumm, J.; Mi, K.; Bilder, P.; Sun, M.; Lim, J.; Bielefeldt-Ohmann, H.; Basaraba, R.; So, M.; Zhu, G.; Tufariello, J.M.; et al. Mycobacterium tuberculosis Universal Stress Protein Rv2623 regulates bacillary growth by ATP-binding: Requirement for establishing chronic persistent Infection. PLoS Pathog. 2009, 5, e1000460. [Google Scholar] [CrossRef]
- Oberschall, A.; Bourenkov, G.; Strizhov, N.; Bartunik, H. Universal Stress Protein Rv2623 from Mycobacterium Tuberculosis. Available online: http://hasyweb.desy.de/science/annual_reports/2005_report/part2/contrib/79/14846.pdf (accessed on 5 May 2021).
- Monahan, I.M.; Betts, J.; Banerjee, D.K.; Butcher, P.D. Differential expression of mycobacterial proteins following phagocytosis by macrophages. Microbiology 2001, 147, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Jung, Y.-J.; Tyagi, S.; Gennaro, M.L.; North, R.J. Expression of Th1-mediated immunity in mouse lungs induces a Mycobacterium tuberculosis transcription pattern characteristic of nonreplicating persistence. Proc. Natl. Acad. Sci. USA 2002, 100, 241–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.; Sharma, P.; Kumar, M.; Joshi, B.; Bisht, D. Analysis of intracellular expressed proteins of Mycobacterium tuberculosis clinical isolates. Proteome Sci. 2012, 10, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, L.N.; Swapna, G.; Chavadi, S.S.; Tufariello, J.M.; Mi, K.; Drumm, J.E.; Lam, T.T.; Zhu, G.; Zhan, C.; Vilchéze, C.; et al. Mycobacterium tuberculosis Universal Stress Protein Rv2623 interacts with the putative ATP binding cassette (ABC) transporter Rv1747 to regulate mycobacterial growth. PLoS Pathog. 2017, 13, e1006515. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, V.; Raghunandanan, S.; Gomez, R.L.; Jose, L.; Surendran, A.; Ramachandran, R.; Pushparajan, A.R.; Mundayoor, S.; Jaleel, A.; Kumar, R.A. Profiling the proteome of mycobacterium tuberculosis during dormancy and reactivation. Mol. Cell. Proteom. 2015, 14, 2160–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Lata, M.; Faheem, M.; Khan, A.U.; Joshi, B.; Venkatesan, K.; Shukla, S.; Bisht, D. Role of M. tuberculosis protein Rv2005c in the aminoglycosides resistance. Microb. Pathog. 2019, 132, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, J.M.; Soares, N.C. Associating H2O2-and NO-related changes in the proteome of Mycobacterium smegmatis with enhanced survival in macrophage. Emerg. Microbes Infect. 2018, 7, 1–17. [Google Scholar] [CrossRef]
- Anderson, C.; Kendall, M.M. Salmonella enterica Serovar Typhimurium strategies for host adaptation. Front. Microbiol. 2017, 8, 1983. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Li, Y.; Zhao, J.; Ma, H.; Wang, J.; Liang, B.; Du, X.; Wu, F.; Xia, S.; Yang, X.; et al. The phenotypic and molecular characteristics of antimicrobial resistance of Salmonella enterica subsp. enterica Serovar Typhimurium in Henan Province, China. BMC Infect. Dis. 2020, 20, 511. [Google Scholar] [CrossRef]
- Hermans, A.P.H.M. Stress Response and Virulence in Salmonella Typhimurium: A Genomics Approach. Ph.D. Thesis, Wageningen University and Research Staff, Wageningen, The Netherlands, 2007. Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/121871 (accessed on 25 March 2021).
- Kikuchi, Y.; Ohara, N.; Sato, K.; Yoshimura, M.; Yukitake, H.; Sakai, E.; Shoji, M.; Naito, M.; Nakayama, K. Novel stationary-phase-upregulated protein of Porphyromonas gingivalis influences production of superoxide dismutase, thiol peroxidase and thioredoxin. Microbiology 2005, 151, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Capestany, C.A.; Tribble, G.D.; Maeda, K.; Demuth, D.R.; Lamont, R.J. Role of the Clp system in stress tolerance, biofilm formation, and intracellular invasion in Porphyromonas gingivalis. J. Bacteriol. 2008, 190, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.; Bhat, S.G.; Sivaraman, K. Porphyromonas gingivalis adopts intricate and unique molecular mechanisms to survive and persist within the host: A critical update. J. Oral Microbiol. 2020, 12, 1090. [Google Scholar] [CrossRef]
- Chen, W.; Homma, K.; Sharma, A.; Kuramitsu, H.K. A Universal Stress Protein of Porphyromonas gingivalisis involved in stress responses and biofilm formation. FEMS Microbiol. Lett. 2006, 264, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Henry, L.G.; Boutrin, M.-C.; Aruni, A.W.; Robles, A.; Ximinies, A.; Fletcher, H.M. Life in a diverse oral community—Strategies for oxidative stress survival. J. Oral Biosci. 2014, 56, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Henry, L.G.; McKenzie, R.M.; Robles, A.; Fletcher, H.M. Oxidative stress resistance in Porphyromonas gingivalis. Futur. Microbiol. 2012, 7, 497–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutrin, M.-C.; Wang, C.; Aruni, W.; Li, X.; Fletcher, H.M. Nitric oxide stress resistance in Porphyromonas gingivalis is mediated by a putative hydroxylamine reductase. J. Bacteriol. 2012, 194, 1582–1592. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.P.; Iyer, D.; Anaya-Bergman, C. Adaptation of Porphyromonas gingivalis to microaerophilic conditions involves increased consumption of formate and reduced utilization of lactate. Microbiology 2009, 155, 3758–3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myles, I.; Datta, S.K. Staphylococcus aureus: An introduction. Semin. Immunopathol. 2012, 34, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Naber, C.K. Staphylococcus aureus bacteremia: Epidemiology, pathophysiology, and management strategies. Clin. Infect. Dis. 2009, 48, S231–S237. [Google Scholar] [CrossRef] [Green Version]
- Stark, L. Staphylococcus aureus: Aspects of Pathogenesis and Molecular Epidemiology. Ph.D. Thesis, Linköping University, Linköping, Sweden, 2013. Available online: http://www.diva-portal.org/smash/get/diva2:647005/FULLTEXT01.pdf (accessed on 12 April 2021).
- Cheng, A.G.; DeDent, A.C.; Schneewind, O.; Missiakas, D. A play in four acts: Staphylococcus aureus abscess formation. Trends Microbiol. 2011, 19, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Attia, A.S.; Cassat, J.E.; Aranmolate, S.O.; Zimmerman, L.J.; Boyd, K.L.; Skaar, E.P. Analysis of the Staphylococcus aureus abscess proteome identifies antimicrobial host proteins and bacterial stress responses at the host-pathogen interface. Pathog. Dis. 2013, 69, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Kylväjä, R.; Kankainen, M.; Holm, L.; Westerlund-Wikström, B. Adhesive polypeptides of Staphylococcus aureus identified using a novel secretion library technique in Escherichia coli. BMC Microbiol. 2011, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; McCann, M.; Kavanagh, K. Proteomic analysis of the proteins released from Staphylococcus aureus following exposure to Ag(I). Toxicol. Vitr. 2013, 27, 1644–1648. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Farber, J.; Peterkin, P. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.S.; Izar, B.; Pazan, F.; Mohamed, W.; Abu Mraheil, M.; Mukherjee, K.; Billion, A.; Aharonowitz, Y.; Chakraborty, T.; Hain, T. Universal Stress Proteins are important for oxidative and acid stress resistance and growth of Listeria monocytogenes EGD-e in vitro and in vivo. PLoS ONE 2011, 6, e24965. [Google Scholar] [CrossRef] [Green Version]
- Wemekamp-Kamphuis, H.H.; Wouters, J.A.; de Leeuw, P.P.L.A.; Hain, T.; Chakraborty, T.; Abee, T. Identification of sigma factor σ b-controlled genes and their impact on acid stress, high hydrostatic pressure, and freeze survival in listeria monocytogenes EGD-e. Appl. Environ. Microbiol. 2004, 70, 3457–3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hain, T.; Hossain, H.; Chatterjee, S.S.; Machata, S.; Volk, U.; Wagner, S.; Brors, B.; Haas, S.; Kuenne, C.T.; Billion, A.; et al. Temporal transcriptomic analysis of the Listeria monocytogenes EGD-e σB regulon. BMC Microbiol. 2008, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, S.S.; Hung, D.T. Persistent bacterial infections, antibiotic tolerance, and the oxidative stress response. Virulence 2013, 4, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, S.; Mitchell, M.; Havis, S.; Bodunrin, A.; Rangel, J.; Olson, G.; Widger, W.R.; Bark, S.J. A proteomic signature of dormancy in the actinobacterium Micrococcus luteus. J. Bacteriol. 2017, 199, e00206. [Google Scholar] [CrossRef] [Green Version]
- Havis, S.; Rangel, J.; Mali, S.; Bodunrin, A.; Housammy, Z.; Zimmerer, R.; Murphy, J.; Widger, W.R.; Bark, S.J. A color-based competition assay for studying bacterial stress responses in Micrococcus luteus. FEMS Microbiol. Lett. 2019, 366, fnz054. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masamba, P.; Kappo, A.P. Parasite Survival and Disease Persistence in Cystic Fibrosis, Schistosomiasis and Pathogenic Bacterial Diseases: A Role for Universal Stress Proteins? Int. J. Mol. Sci. 2021, 22, 10878. https://doi.org/10.3390/ijms221910878
Masamba P, Kappo AP. Parasite Survival and Disease Persistence in Cystic Fibrosis, Schistosomiasis and Pathogenic Bacterial Diseases: A Role for Universal Stress Proteins? International Journal of Molecular Sciences. 2021; 22(19):10878. https://doi.org/10.3390/ijms221910878
Chicago/Turabian StyleMasamba, Priscilla, and Abidemi Paul Kappo. 2021. "Parasite Survival and Disease Persistence in Cystic Fibrosis, Schistosomiasis and Pathogenic Bacterial Diseases: A Role for Universal Stress Proteins?" International Journal of Molecular Sciences 22, no. 19: 10878. https://doi.org/10.3390/ijms221910878